Abstract
Biogenic volatile organic compounds (VOCs) serve as signals mediating information between plants and their higher trophic level beneficials, such as parasitoids and predators of herbivores. We recently demonstrated with oilseed rape (Brassica napus L.) plants, herbivorous diamond-back moth (Plutella xylostella L. (Lepidoptera: Yponomeutidae)) larvae and Cotesia vestalis (Haliday) (Hymenoptera: Braconidae) parasitoids that atmospheric pollution, i.e. elevated ozone (O3), can disturb attraction of natural enemies by plant-emitted host-induced volatile cues. Additionally, we found that the degree of herbivore feeding damage is an important contributor to this O3 interference. low feeding damage in herbivore-resistant plants was sufficient to attract C. vestalis females to host-damaged plants under ambient air, but this tritrophic signaling turned non-functional in the combination of low feeding damage and high O3 concentration. Here we present some additional data of how climate change factors may modify herbivore feeding and their growth. We further discuss how the degree of herbivore feeding damage and the tritrophic signaling interaction relaying on the herbivore-induced VOCs from attacked plants might change through direct and indirect effects of increased levels of carbon dioxide, temperature and O3.
The global atmospheric carbon dioxide (CO2) level is the highest met in the last 650,000 years and continues to contribute to the global average temperature rise.Citation1 Regional ozone (O3) concentrations have increased markedly due to environmental pollution,Citation1 and already act phytotoxically towards sensitive plant species.Citation2 Plants emit various types of volatile organic compounds (VOCs) with diverging ecological roles into their surrounding atmosphere.Citation3 Higher trophic level parasitoids and predators utilize herbivore-inducible VOC cues, i.e., compounds emitted after feeding leading to quantitative or even qualitative differences in the volatile profile, to separate host-carrying from intact plants.Citation4 Elevated CO2 has most often caused seasonal reductions in the biogenic emissions of terpenoids,Citation5,Citation6 which are among main VOCs affecting plant-herbivore-parasitoid interactions.Citation3,Citation4 O3 directly reacts with and degrades O3-sensitive terpenoids, yet certain O3-resistant VOCs, such as 1,8-cineole, will remain unaffected.Citation7 However, disturbance of multitrophic VOC signaling by changes in CO2 or O3 levels have yet seldom been reported.Citation7,Citation8 One example exists: elevated CO2 disturbing orientation of Cotesia vestalis to diamond-back moth (Plutella xylostella L.)-damaged cabbage (Brassica oleracea ssp. capitata) plants.Citation9 Nevertheless, more experimentation is definitely needed. Previously, O3 has not affected the orientation of this parasitoid to host-damaged cabbage plants,Citation7 which demonstrated its sophisticated olfactory ability to use specific O3-resistant cues for host finding in Brassicas. Yet another important aspect is the active role of the herbivore in plant-parasitoid interactions affected by elevated CO2 or O3. This was studied recently by us with the same parasitoid species and experimental system, but including herbivore-resistant and -sensitive oilseed rape (Brassica napus L.) plants.Citation10
Plant resistance to herbivores is an important target of breeding allowing combating against rising pest problems in agriculture. The first commercial transgenic insect-resistant plants produce specific Bacillus thuringiensis (Bt) toxins contributing resistance against their target insects.Citation11 In our recent work we used Lepidoptera-resistant Bt oilseed rape plants to compare the herbivore-induction of VOCs from these and their herbivore-sensitive parent line plants under elevated CO2 or O3 level.Citation10 Interestingly, we showed that O3 interference of volatile induction became relevant with the resistant Bt plants. Even though the resistant plants allowed feeding initiation that enabled the parasitoids to find host-damaged resistant plants under ambient air, elevated O3 rendered these emissions undistinguishable for the parasitoid compared to intact plant emissions. As important for the sustainable use of herbivore-resistant cultivars, either of genetically modified or traditionally bred origin, their wider use in O3-polluted areas might therefore result in lowered natural enemy population abundance (with the example of C. vestalis) by lowered VOC attraction and also by direct negative effects by reduced host density or poorer host quality.Citation12 However, as this type of bred resistance is a means of specific herbivore control, comparative direct control by pesticides might have even more negative effects.Citation13 Importantly, mechanical damage has induced similar VOC emissions from Bt and non-Bt line plants.Citation14 The unchanged ability for induction by non-target herbivores might assist in attracting beneficials, which could be killed when using non-specific pesticides.
Another interesting perception raised by our previous work was that in general, the changes in the degree of herbivore feeding damage by climate change factors potentially affecting multitrophic signaling has been largely understudied. Predicting effects of changed CO2 and temperature or O3 on volatile-mediated multitrophic interactions is highly complex (), since their effects can act on more than only one level of the food chain and might further interact. For example, alterations in host plant quality can change the degree or pattern of herbivore feeding, which could lead to an altered defence response in the plant itself, such as increased release of VOCs. Then, altered host quality may affect the population development of the third trophic level parasitoid and VOC cues guiding to host may be different.
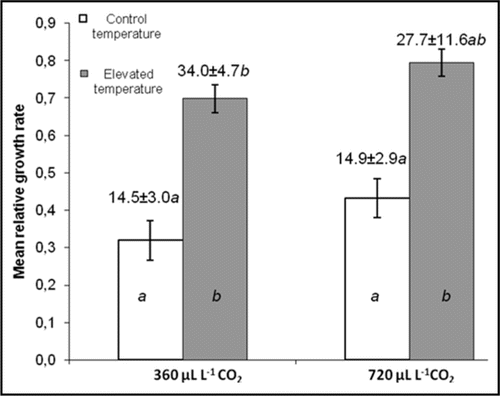
It is known that elevated CO2 often causes compensatory feeding through reduced plant nutritional quality, e.g., by lowered nitrogen and higher carbon content,Citation15,Citation16 which might lead to increased VOC emissions, assuming that feeding damage is directly proportional to the amount of emissions. We detected higher induction of emissions from oilseed rape under elevated CO2, yet the degree of feeding damage caused in 48 hours was not different between the control CO2 and doubled CO2-grown plants.Citation10 However, if feeding patterns were recorded for a longer time, differences between the CO2 treatments might have arisen. In another experiment on oilseed rape, we measured the leaf area consumed by diamond-back moth larvae and their mean relative growth rate under elevated CO2, elevated temperature and combined elevated CO2 and temperature. Again, there was no difference between the CO2 treatments of this 48 h trial (). However, the clear increase by elevated temperature in both parameters measured suggests that direct effects by climate change-related abiotic factors may cause rapid changes in herbivory, whereas plant-mediated effects predominantly appear over longer time scales.Citation16 These might have longer-lasting seasonal effects, yet rapid changes by direct effects can be more unpredictable and could have even dramatic regional consequences.
Effects of O3 on VOC signaling are complicated, since O3 has been reported to induce volatile emissionsCitation8,Citation17,Citation18 and thus potential interference with herbivore-induced VOCs is possible, although this was not observed in a tritrophic interaction between lima bean (Phaseolus lunatus) plants, two-spotted spider mites (Tetranychus urticae) and their specialist predators.Citation8 O3 usually makes leaves more palatable to herbivores by increased solubility and availability of nitrogenous compounds,Citation19,Citation20 though the possibility of direct negative effects by high O3 on feeding of P. xylostella larvae was recently suggested.Citation21 The challenge of working with highly reactive O3 and VOCs requires sophisticated experimental set-ups,Citation7 which to date has restricted more general findings of VOCs and multitrophic communication in field set-ups with elevated O3.Citation22 Recent reports that root herbivory affects shoot VOC signaling and behavior of aboveground natural enemiesCitation23 or vice versa,Citation24 allows speculating that climate change factors might affect defence signaling between belowground and aboveground plants parts through their known effects on within-plant assimilate partitioning.Citation25 Simultaneous analysis of belowground and aboveground VOC interactions in CO2 and O3 studies might help to unravel whether the growth-defence trade-off or altered within-plant resource allocation affects VOC signaling.
To conclude, we underline the importance of the degree of herbivore feeding damage responding directly to elevated CO2 or O3 or indirectly to plant-mediated changes, when considering functioning of multitrophic communication in future atmospheres. An increasing number of multitrophic systems, representing different genera of plants and feeding guilds of insects, as well as both direct and indirect effects of CO2 and O3 need to be included in order to evaluate overall effects by these abiotic factors on VOC signaling. The various ecological roles suggested for volatile signals emitted, and being detected by many types of perceivers,Citation3 make predictions of potential changes in volatile infochemical networks by climate change an important task to be figured out for allowing understanding of multitrophic interactions in future crop and natural ecosystems.
Figures and Tables
Figure 1 Proposed effects of elevated temperature, carbon dioxide or ozone concentrations on three different trophic levels, which in turn might affect multitrophic volatile infochemical signaling. The example illustrates a Brassica plant, the specialist herbivore Plutella xylostella L. and its endoparasitoid Cotesia vestalis.
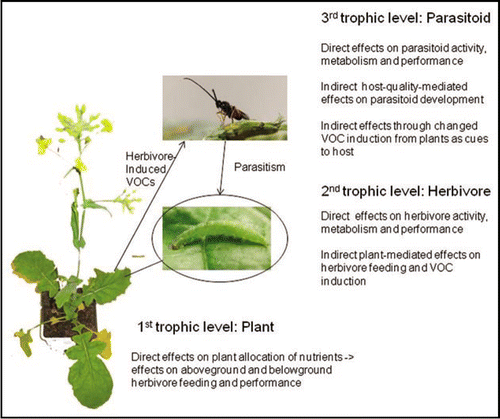
Figure 2 Mean relative growth rate ± SEM (bars) of diamondback moth (Plutella xylostella L.) larvae and the leaf area consumed ± SEM (numbers above bars, mm2) by these larvae feeding on 19-d old vegetative stage oilseed rape (Brassica napus L.) plants grown and tested under control CO2 (360 µl l−1) or elevated (720 µl l−1) CO2 and control (16/20°C) or elevated (20/24°C) temperature. Different letters indicate statistically significant differences between treatments (ANOVA followed by LSD post hoc test), n = 6. Treatments were performed as in reference 26 and mean relative growth rate and feeding area determined as in reference Citation21.
Acknowledgements
This work was supported by the Academy of Finland (ESGEMO Programme, decision no. 105209).
Addendum to:
References
- IPCC, Intergovernmental Panel on Climate Change. Working group I report: The physical science basis, Technical summary 2007; http://www.ipcc.ch/
- Ashmore MR. Assessing the future global impacts of ozone on vegetation. Plant Cell Environm 2005; 28:949 - 964
- Holopainen JK. Multiple functions of inducible plant volatiles. Trends Plant Sci 2004; 11:529 - 533
- De Moraes CM, Lewis WJ, Pare PW, Alborn HT, Tumlinson JH. Herbivore-infested plants selectively attract parasitoids. Nature 1998; 393:570 - 573
- Loreto F, Fischbach RJ, Schnitzler JP, Ciccioli P, Brancaleoni E, Calfapietra C, et al. Monoterpene emission and monoterpene synthase activities in the Mediterranean evergreen oak Quercus ilex L. grown at elevated CO2 concentrations. Glob Change Biol 2001; 7:709 - 717
- Rapparini F, Baraldi R, Miglietta F, Loreto F. Isoprenoid emission in trees of Quercus pubescens and Quercus ilex with lifetime exposure to naturally high CO2 environment. Plant Cell Environm 2004; 27:381 - 391
- Pinto DM, Blande JD, Nykänen R, Dong W-X, Nerg A-M, Holopainen JK. Ozone degrades common herbivore-induced plant volatiles: does this affect herbivore prey location by predators and parasitoids?. Journal of Chemical Ecology 2007; 33:683 - 694
- Vuorinen T, Nerg A-M, Holopainen JK. Ozone exposure triggers the emission of herbivore-induced plant volatiles, but does not disturb tritrophic signalling. Environm Poll 2004; 131:305 - 311
- Vuorinen T, Nerg A-M, Ibrahim MA, Reddy GVP, Holopainen JK. Emission of Plutella xylostella-induced compounds from cabbages grown at elevated CO2 and orientation behavior of the natural enemies. Plant Physiol 2004; 135:1984 - 1992
- Himanen SJ, Nerg AM, Nissinen A, Pinto DM, Stewart CN Jr, Poppy GM, et al. Effects of elevated carbon dioxide and ozone on volatile terpenoid emissions and multitrophic communication of transgenic insecticidal oilseed rape (Brassica napus). New Phytol 2009; 181:174 - 186
- James C. Executive summary of global status of commercialized biotech/GM crops: 2007 2007; Ithaca NY ISAAA ISAAA Brief No. 37 http://www.isaaa.org
- Schuler TH, Denholm I, Clark SJ, Stewart CN Jr, Poppy GM. Effects of Bt plants on the development and survival of the parasitoid Cotesia plutellae (Hymenoptera: Braconidae) in susceptible and Bt-resistant larvae of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). J Ins Physiol 2004; 50:435 - 440
- Romeis J, Meissle M, Bigler F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat Biotechnol 2006; 24:63 - 71
- Dean JM, De Moraes CM. Effects of genetic modification on herbivore-induced volatiles from maize. J Chem Ecol 2006; 32:713 - 724
- Stiling P, Cornelissen T. How does elevated carbon dioxide (CO2) affect plant-herbivore interactions? A field experiment and meta-analysis of CO2-mediated changes on plant chemistry and herbivore performance. Glob Change Biol 2007; 13:1823 - 1842
- Zvereva EL, Kozlov MV. Consequences of simultaneous elevation of carbon dioxide and temperature for plant-herbivore interactions: a meta-analysis. Glob Change Biol 2006; 12:27 - 41
- Heiden AC, Hoffmann T, Kahl J, Kley D, Klockow D, Langebartels C, et al. Emission of volatile organic compounds from ozone-exposed plants. Ecol Appl 1999; 9:1160 - 1167
- Llusiá J, Peñuelas J, Gimeno BS. Seasonal and species-specific response of VOC emissions by Mediterranean woody plant to elevated ozone concentrations. Atmosph Environm 2002; 36:3931 - 3938
- Valkama E, Koricheva J, Oksanen E. Effects of elevated O3, alone and in combination with elevated CO2, on tree leaf chemistry and insect herbivore performance: a meta-analysis. Glob Change Biol 2007; 13:184 - 201
- Jondrup PM, Barnes JD, Port GR. The effect of ozone fumigation and different Brassica rapa lines on the feeding behavior of Pieris brassicae larvae. Entomol Exp Appl 2002; 104:143 - 151
- Himanen SJ, Nerg A-M, Nissinen A, Stewart CN Jr, Poppy GM, Holopainen JK. Elevated atmospheric ozone increases concentration of insecticidal Bacillus thuringiensis (Bt) Cry1Ac protein in Bt Brassica napus and reduces feeding of a Bt target herbivore on the non-trangenic parent. Environm Poll 2009; 157:181 - 185
- Pinto DM, Himanen SJ, Nissinen A, Nerg A-M, Holopainen JK. Host location behavior of Cotesia plutellae Kurdjumov (Hymenoptera: Braconidae) in ambient and moderately elevated ozone in field conditions. Environm Poll 2008; 156:227 - 231
- Soler R, Harvey JA, Kamp AFD, Vet LEM, van der Putten WH, van Dam NM, et al. Root herbivores influence the behaviour of an aboveground parasitoid through changes in plant-volatile signals. Oikos 2007; 116:367 - 376
- Kaplan I, Halitschke R, Kessler A, Rehill BJ, Sardanelli S, Denno RF. Physiological integration of roots and shoots in plant defense strategies links above- and belowground herbivory. Ecol Letts 2008; 11:841 - 851
- Grantz DA, Gunn S, Vu HB. O3 impacts on plant development: a meta-analysis of root/shoot allocation and growth. Plant Cell Environm 2006; 29:1193 - 1209
- Himanen SJ, Nissinen A, Dong W-X, Nerg A-M, Stewart CN Jr, Poppy GM, et al. Interactions of elevated carbon dioxide and temperature with aphid feeding on transgenic oilseed rape: Are Bacillus thuringiensis (Bt) plants more susceptible to nontarget herbivores in future climate?. Glob Change Biol 2008; 14:1437 - 1454