Abstract
The phytohormone cytokinin is a key player in many developmental processes and in the response of plants to biotic and abiotic stress. The cytokinin signal is perceived and transduced via a multistep variant of the bacterial two-component signaling system. Most of the research on cytokinin signaling has been done in the model plant Arabidopsis thaliana. Research on cytokinin signaling has expanded to a much broader range of plants species in recent years. This is due to the natural limitation of Arabidopsis as a model species for the investigation of processes like nodulation or wood formation. The rapidly increasing number of sequenced plant genomes also facilitates the use of other species in this line of research. This review summarizes what is known about the cytokinin signaling in the different organisms and highlights differences to Arabidopsis.
Phytohormones are well known regulators of plant development.Citation1 In addition, they also play an important role in mediating the response of plants to biotic and abiotic factors in the environment. Cytokinins, N6-substituted adenine derivates, are one class of phytohormones.Citation2 The cytokinin signal is perceived and transduced via a multi-step variant of the two-component system (TCS). While the TCS is the principal signaling system in bacteria, among higher eukaryotes it is found only in plants.Citation3
The current model of cytokinin signaling predicts that cytokinin is perceived by a membrane bound hybrid histidine kinase receptor (HK). The binding of the ligand leads to an autophosphorylation of the receptor and is followed by an intramolecular transfer of the phosphoryl residue to the receiver domain of the receptor. Subsequently, the phosphate is transferred to a histidine phosphotransfer protein (HPt), which translocates to the nucleus where it activates type-B response regulators (type-B RRs). This class of response regulators are Myb-type transcription factors and among their target genes are the type-A response regulators (type-A RRs). The type-A RRs act as negative feedback regulators of the cytokinin signaling pathway.Citation4
Recently, phylogenetic analysis aimed at unraveling the origin of this signaling pathway identified the moss Physcomitrella patens as the most early diverging organism examined to encode members of all four protein families in its genome. While the analysis of the also investigated algal species revealed the presence of genes for HPts and type-B RRs, no sequences encoding for putative cytokinin receptors or type-A RRs were detected.Citation5
Most of the research on cytokinin signaling to date has been carried out in the model plant Arabidopsis thaliana. However, studies in other plants have not only contributed to the elucidation of this pathway but also highlighted the cross-talk between cytokinin signaling and other non-hormone signaling pathways, i.e., signaling pathways regulating nitrogen and sulfur metabolism,Citation6–Citation9 nodulation,Citation10–Citation14 pathogen defense,Citation15 heavy metal responseCitation16 and drought stress.Citation17,Citation18
The function of cytokinin signaling genes are investigated in two types of experiments. Single genes of the cytokinin signaling pathway were identified in screens connecting cytokinin signaling to the respective stimuli. Systematic approaches characterizing all family members of the cytokinin signaling pathway in a given species were conducted in species with completely sequenced genomes. Examples for such methodical experiments can be found in rice, poplar, lotus and grape.Citation7,Citation19–Citation24 With the rapidly increasing number of completely sequenced plant genomes and the new possibilities offered by the arrival of the next generation sequencing techniques for transcriptome analysis, the opportunity for a larger number of targeted analysis in plant species other than Arabidopsis thaliana is growing dramatically. This is not only welcome, but also necessary as the influence or crosstalk of cytokinin in many biological processes, such as nodulation, wood formation and pathogen response to species not affecting Arabidopsis cannot at all or can only rudimentarily be investigated in Arabidopsis. Thus cytokinin research is bound to profit greatly from a broadening in focus to include diverse plant species.
The aim of this review is to summarize the current state of knowledge about cytokinin signaling in different plant species beyond Arabidopsis thaliana (for a comprehensive overview see Sup. Table 1). In the following sections we will review what is known for members of each of the four protein families involved in the cytokinin signaling pathway. For a detailed description of cytokinin signaling in Arabidopsis see recent reviews.Citation25–Citation27
Cytokinin Receptors
One of the processes where cytokinin plays a crucial role but cannot be investigated in Arabidopsis is root nodule formation. Three publications describe the role of a cytokinin receptor from lotus, Lotus Histidine Kinase 1 (LHK1), in this symbiotic process.Citation10,Citation12,Citation14 Receptor loss-of-function mutants failed to respond to the rhizobial signal and did not perform the first step of nodule formation—the division of cortical cells.Citation12 In contrast, a constitutively active mutant displayed spontaneous development of nodules even in the absence of the symbiotic rhizobial bacteria or their signaling molecules.Citation14 This spontaneous nodulation can be suppressed by addition of abscisic acid (ABA).Citation10 Expression analysis detected receptor transcripts in uninfected root tissues as well as in nodules. Heterologous complementation of bacterial and yeast mutants further confirmed LHK1 function as a cytokinin receptor.Citation12,Citation14 Medicago truncatula is also a root nodule forming species containing three cytokinin receptors. An RNAi approach indicated that at least one of the putative cytokinin receptors also acts in nodule formation.Citation11 Experimental data showing expression of the histidine kinase genes in newly formed nodules supported this assessment.Citation28,Citation29 Further expression analysis indicated that at least two of the cytokinin receptors in Medicago might play a role in abiotic stress response as well.Citation30
Another aspect of plant biology where Arabidopsis cannot serve as a model is wood formation and the influence that cytokinin has on this developmental process. With the completed genomic sequence at hand, poplar is often used as a model organism for tree biology. As cambium is the tissue which gives rise to wood and cytokinin plays a decisive role in cambium development,Citation31 putative cytokinin receptors of poplar have been investigated in this context.Citation32 For four of the five putative cytokinin receptors of poplar, expression was detected in the cambium. The same group investigated the expression pattern of a putative cytokinin receptor from birch and found expression in the cambial zone as well. Several experiments including analysis of grafts, callus formation and stem diameter using cytokinin deficient transgenic poplar plants clearly demonstrated the role of cytokinin as a major regulator of cambium development.Citation32
Other functional studies of cytokinin receptors have been focused on their core role in cytokinin signaling itself. In 2004, Yonekura-Sakakibara and colleagues investigated the expression patterns of three cytokinin receptors of corn in various tissues and while they found large overlaps in the temporal and spatial expression patterns of the three receptors, they also detected tissue specificity of the receptor transcripts.Citation33,Citation34 The cytokinin binding of the maize receptors revealed specificities similar to those detected in the respective homologs in Arabidopsis.Citation35,Citation36 Data describing putative cytokinin receptors in other plant species are limited to transcriptome analysis. Experiments in rice detected transcripts of putative cytokinin receptors in all tissues investigated.Citation19,Citation21,Citation37 At least in one case the transcription level of the receptors increased in response to salt stress and dehydration, indicating a role for cytokinin in response to abiotic stress.Citation17,Citation18 Northern blot analysis in Catharanthus roseus showed a strong expression of a putative cytokinin receptor into several organs, but mainly in the flower.Citation38 Expression of homologs of the Arabidopsis cytokinin receptors were also detected in lupine,Citation39 tomato,Citation40 potatoCitation15 and in grape.Citation7 The main features of the cytokinin receptors across the different species have been compiled in .
Histidine Phosphotransfer Proteins
For most HPt proteins the focus of research has been on their role within the cytokinin signaling pathway. Several lines of evidence suggest a conserved role of HPts in maize and Arabidopsis. Transiently expressed ZmHP1-GFP localized to both the cytosol and the nucleus, as one would predict from the current model for cytokinin signaling.Citation41 Phosphorelay and yeast two-hybrid experiments showed that at least ZmHP1, ZmHP2 and ZmHP3 are interacting with ZmRRs of both type-A and type-B families and that this interaction mediates a phosphotransfer.Citation41 In addition, the maize ZmHP2 is the only full-length protein of the cytokinin signaling pathway in any species for which the three dimensional structure has been solved. This structure revealed that all the amino acids, which are conserved in plant HPts, surround the canonical histidine residue—probably forming a docking interface for the receiver domain of the receptors and response regulators.Citation42 For ZmHP1, crystals and preliminary X-ray spectra were also reported, but thus far no crystal structure has been published.Citation43
Functional analysis of HPts was also carried out in Catharanthus roseus. In cell-lines transformed with a CrHP1 RNAi construct induction of the type-A RR CrRR1 transcript was reduced, while the expression of the type-B RR CrRR5 remained unchanged by cytokinin treatment.Citation13 The expression of CrHP1 itself in wild-type cell culture lines was not responsive to cytokinin. However, the addition of jasmonic acid or auxin led to an increase in CrHP1 expression, indicating a regulatory crosstalk between the signaling pathways of these phytohormones.Citation13
In Medicago one of the HPts, MtHP2, was found to be upregulated upon salt stress, further corroborating the link between cytokinin and abiotic stress signaling detected in Arabidopsis.Citation44,Citation45 Transcripts of the HPt genes were detected in additional species, namely in orange,Citation46 rice,Citation18,Citation19,Citation21,Citation37 poplar,Citation47 wheatCitation48 and in grape.Citation7 The main features of the histidine phosphotransfer proteins across the different species have been compiled in .
Type-B Response Regulators
In Medicago truncatula not only the cytokinin receptors were indicated in the nodule formation as described above, but also one of the three type-B RRs, MtRR1. This type-B RR was strongly induced in the earlier stages of nodule formation. However, MtRR1 RNAi plants did not display an altered nodulation phenotype and further research is needed to clarify the role of MtRR1 in this process.Citation11
One of the poplar type-B RRs, PtRR13, was identified in a microarray experiment to be involved in adventitious root development. This finding was later confirmed by the phenotypes displayed by PtRR13 RNAi and transgenic dominant negative (ΔDDKPtRR13) plants, which showed defects in this process.Citation24 Analysis of the expression patterns of all type-B RRs from poplar showed for at least three of the eleven type-B RRs a clear induction of gene expression after cytokinin treatment.Citation23 This is surprising as according to the current model of cytokinin signaling, the type-A RRs are the only component of the pathway being transcriptionally regulated by cytokinin treatment and numerous microarray experiments from Arabidopsis confirmed this hypothesis.Citation26,Citation49,Citation50
In maize, Asakura and colleagues showed that the three analyzed type-B RRs, ZmRR8, ZmRR9 and ZmRR10 can interact with the three ZmHPs in a yeast two-hybrid assay. These interactions were also verified in phosphorelay experiments for ZmRR8 and ZmRR9.Citation41 Subcellular localization showed ZmRR8 to be located in the nucleus. The expression of all three type-B ZmRRs was not affected by cytokinin treatment.Citation41
Taking advantage of the complete genome sequence, two groups analyzed the expression patterns of various TCS components in rice and could show differential expression patterns for the type-B OsRRs in various tissues, developmental stages and in response to abiotic stresses.Citation17,Citation19 The main features of the type-B RRs across the different species have been compiled in .
Type-A Response Regulators
Research on type-A RRs in maize points to roles in phyllotaxis and seed development. Aberrant phyllotaxy1 (abphyl1; also Zmrr3) was shown to be a regulator of embryo morphogenesis and shoot phyllotaxis.Citation51 The ABPHYL1 expression was localized to the embryo and shoot apex.Citation52 Detailed analysis of the mutant concluded that the phenotype was caused by reduced auxin levels leading to a larger shoot apical meristem, a delayed leaf initiation and altered leaf phyllotaxy.Citation53 The type-A RRs ZmTcRR1 and ZmTcRR2 were reported to be specifically expressed in the transfer cell layer of developing maize kernels; however, so far no specific function was assigned to these genes.Citation33,Citation54 Other members of type-A RRs of maize were characterized for their role in cytokinin signaling. On the protein level, type-A RRs were shown to interact with ZmHPs in yeast two-hybrid and phosphorelay experiments. These in vitro experiments showed also that type-A ZmRRs auto-dephoshorylated much faster than type-B ZmRRs,Citation41 which might be an important trait for their function as negative regulators of the cytokinin signaling pathway. Three type-A RRs, ZmRR1, ZmRR2 and ZmRR3, were shown to localize to the cytosol and the nucleus, while ZmRR4, ZmRR5 and ZmRR6 were found exclusively in the nucleus of transiently transformed onion epidermis cells.Citation41 The transcription of all type-A ZmRRs expressed in the leaf blade was shown to be strongly induced by cytokinin.Citation6,Citation41,Citation55 ZmRR1 and ZmRR2 were found to be involved in the nitrogen signal transduction,Citation8,Citation9,Citation56,Citation57 a node of crosstalk which has also been discovered in Arabidopsis.Citation58
Experimental characterization of the type-A RRs of rice further emphasized the role of this protein family in plant development. Calli overexpressing OsRR6 were severely retarded in shoot regeneration and transgenic plants displayed a general dwarf phenotype with a poorly developed root system—consistent with a role of OsRR6 as a negative regulator of cytokinin signaling.Citation59 Rice plants overexpressing either OsRR3 or OsRR5 exhibited a lower cytokinin sensitivity in root elongation and callus growth assays. Furthermore, in these transgenic plants other type-A RRs such as OsRR1, OsRR7, OsRR14 and OsRR15 were downregulated, while other members of the same gene family showed a higher level of transcription than in the wild type control.Citation60 Also type-A RRs from other species were implicated in plant development. A recent study hints at a role for type-A RR, PipiRR1, during caulogenic induction in Pinus pinea. The respective transcript was detected in the cotyledons and increased after treatment with cytokinin in a dose-dependent manner.Citation61
Type-A RRs of other species were shown to react to abiotic stimuli. The transcript level of PvRR1 of Phaseolus vulgaris increased during starvation experiments related to macronutrients such as phosphorus, potassium and nitrogen as well as upon addition of cytokinin, while it decreased after resupply of the nutrients.Citation62 Two type-A RRs of Medicago, MtRR4 and MtRR5, were upregulated in response to salt stress, but also during nodulation.Citation30 In contrast, the three type-A RRs of Catharanthus roseus seemed to be specifically induced by cytokinin.Citation13,Citation63,Citation64
Systematic expression studies for all type-A RRs of a given species have been done in rice, lotus and poplar. The extensive analysis of the complete set of type-A RRs in rice revealed different temporal and spatial expression patterns of this class of OsRRs. The transcript level of most type-A OsRRs increased upon cytokinin treatment.Citation19,Citation21,Citation22 Under different abiotic stress treatments, the expression of the type-A OsRRs was specifically and differentially up or downregulated.Citation17 In Lotus japonicus, the expression of six of the seven tested type-A RRs was induced by cytokinin. For the seventh gene no transcript could be detected and the authors suspected it to be a non-functional gene as parts of the C-terminus are missing.Citation20 In a similar experiment, investigating the members of this gene family in poplar, seven of the eleven type-A PtRRs were found to be induced by cytokinin.Citation23 For PtRR7, a strong expression was found in the cambium. In plants with a lower cytokinin status due to the ectopic expression of AtCKX2, the expression level of PtRR7 was clearly reduced.Citation32 The main features of the type-A RRs across the different species have been compiled in .
Conclusions
In this review we summarized the state of the art in the field of cytokinin signaling beyond Arabidopsis. Some of the signaling components were identified in genetic screens while others were analyzed in systematic approaches following the sequencing of the respective genomes. Many studies link cytokinin to development, crosstalk with other hormones and also to other processes such as abiotic stress or nutrients deficiency response—just as has been shown for Arabidopsis. However, new aspects of cytokinin action that cannot be investigated in Arabidopsis were also discovered, clearly highlighting the necessity to look beyond a single model plant to understand the full spectra of cytokinin-regulated processes. Thus the benefits of cytokinin research in different plant species include: (i) examination of proteins or even whole protein families of the cytokinin signaling pathway that might behave differently than those in Arabidopsis—e.g., cytokinin inducible HPts and type-B RRs; (ii) indentifying the role of cytokinin in other pathways, morphological structures and developmental processes not present in Arabidopsis and (iii) using the wealth of information on the different signaling components to understand the evolution and the evolutionary trajectories of the cytokinin signaling pathway.
Currently experimental data concerning cytokinin signaling derive almost exclusively from angiosperm species (). A deeper insight into the evolution of this pathway requires the inclusion of more early-diverging plant species into this analysis. The rapidly increasing number of sequenced genomes combined with the already established tools of cytokinin research will enhance our understanding of this fascinating plant hormone. The future of cytokinin research is very bright, indeed.
Abbreviations
| TCS | = | two-component system |
| HK | = | histidine kinase |
| HPt | = | histidine phosphotransfer protein |
| RR | = | response regulator |
Figures and Tables
Figure 1 Phylogenetic relation of plant species used in cytokinin signaling research. Species written in grey were used in bioinformatic analysis.Citation5 Black font marks species in which experimental data for cytokinin signaling components have been obtained on the RNA level. In species shaded in grey functional assays have been performed for members of the cytokinin signaling pathway (tree based on Sitte et al. 2002).Citation65
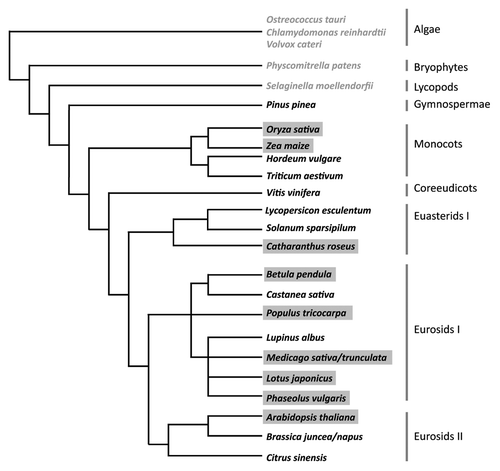
Table 1 Compilation of the function associated with cytokinin receptors from different species
Table 2 Compilation of the function associated with phosphotransfer proteins from different species
Table 3 Compilation of the function associated with type-B response regtulators from different species
Table 4 Compilation of the function associated with type-A response regulators from different species
Additional material
Download Zip (113.8 KB)Acknowledgements
This work was supported by a Elsa Neumann stipend to E.H., by a stipend from the Volkswagen Foundation to N.G. and support of the Dahlem Centre of Plant Science to A.H.. We are also very grateful to Anahid E. Powell for critically reading the manuscript.
References
- Santner A, Calderon-Villalobos LI, Estelle M. Plant hormones are versatile chemical regulators of plant growth. Nat Chem Biol 2009; 5:301 - 307
- Mok DW, Mok MC. Cytokinin metabolism and action. Annu Rev Plant Physiol Plant Mol Biol 2001; 52:89 - 118
- Heyl A, Schmülling T. Cytokinin signal perception and transduction. Curr Opin Plant Biol 2003; 6:480 - 488
- Hwang I, Sheen J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001; 413:383 - 389
- Pils B, Heyl A. Unraveling the evolution of cytokinin signaling. Plant Physiol 2009; 151:782 - 791
- Deji A, Sakakibara H, Okumura S, Matsuda T, Ishida Y, Yamada S, et al. Accumulation of maize response regulator proteins in mesophyll cells after cytokinin treatment. Biosci Biotechnol Biochem 2002; 66:1853 - 1858
- Fernandes J, Tavares S, Amâncio S. Identification and expression of cytokinin signaling and meristem identity genes in sulfur deficient grapevine (Vitis vinifera L.). Plant Signal Behav 2009; 4:1128 - 1135
- Takei K, Sakakibara H, Taniguchi M, Sugiyama T. Nitrogen-dependent accumulation of cytokinins in root and the translocation to leaf: implication of cytokinin species that induces gene expression of maize response regulator. Plant Cell Physiol 2001; 42:85 - 93
- Takei K, Takahashi T, Sugiyama T, Yamaya T, Sakakibara H. Multiple routes communicating nitrogen availability from roots to shoots: a signal transduction pathway mediated by cytokinin. J Exp Bot 2002; 53:971 - 977
- Ding Y, Kalo P, Yendrek C, Sun J, Liang Y, Marsh JF, et al. Abscisic acid coordinates nod factor and cytokinin signaling during the regulation of nodulation in Medicago truncatula. Plant Cell 2008; 20:2681 - 2695
- Gonzalez-Rizzo S, Crespi M, Frugier F. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 2006; 18:2680 - 2693
- Murray JD, Karas BJ, Sato S, Tabata S, Amyot L, Szczyglowski K. A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science 2007; 315:101 - 104
- Papon N, Vansiri A, Gantet P, Chénieux JC, Rideau M, Crèche J. Histidine-containing phosphotransfer domain extinction by RNA interference turns off a cytokinin signalling circuitry in Catharanthus roseus suspension cells. FEBS Lett 2004; 558:85 - 88
- Tirichine L, Sandal N, Madsen LH, Radutoiu S, Albrektsen AS, Sato S, et al. A gain-of-function mutation in a cytokinin receptor triggers spontaneous root nodule organogenesis. Science 2007; 315:104 - 107
- Jolivet K, Grenier E, Bouchet JP, Esquibet M, Kerlan MC, Caromel B, et al. Identification of plant genes regulated in resistant potato Solanum sparsipilum during the early stages of infection by Globodera pallida. Genome 2007; 50:422 - 427
- Srivastava S, Srivastava AK, Suprasanna P, D'Souza SF. Comparative biochemical and transcriptional profiling of two contrasting varieties of Brassica juncea L. in response to arsenic exposure reveals mechanisms of stress perception and tolerance. J Exp Bot 2009; 60:3419 - 3431
- Jain M, Tyagi AK, Khurana JP. Differential gene expression of rice two-component signaling elements during reproductive development and regulation by abiotic stress. Funct Integr Genomics 2008; 8:175 - 180
- Karan R, Singla-Pareek SL, Pareek A. Histidine kinase and response regulator genes as they relate to salinity tolerance in rice. Funct Integr Genomics 2009; 9:411 - 417
- Du L, Jiao F, Chu J, Jin G, Chen M, Wu P. The two-component signal system in rice (Oryza sativa L.): a genome-wide study of cytokinin signal perception and transduction. Genomics 2007; 89:697 - 707
- Ishida K, Niwa Y, Yamashino T, Mizuno T. A genome-wide compilation of the two-component systems in Lotus japonicus. DNA Res 2009; 16:237 - 247
- Ito Y, Kurata N. Identification and characterization of cytokinin-signalling gene families in rice. Gene 2006; 382:57 - 65
- Jain M, Tyagi AK, Khurana JP. Molecular characterization and differential expression of cytokinin-responsive type-A response regulators in rice (Oryza sativa). BMC Plant Biol 2006; 6:1
- Ramírez-Carvajal GA, Morse AM, Davis JM. Transcript profiles of the cytokinin response regulator gene family in Populus imply diverse roles in plant development. New Phytol 2008; 177:77 - 89
- Ramírez-Carvajal GA, Morse AM, Dervinis C, Davis JM. The cytokinin type-B response regulator PtRR13 is a negative regulator of adventitious root development in Populus. Plant Physiol 2009; 150:759 - 771
- Bishopp A, Help H, Helariutta Y. Cytokinin signaling during root development. Int Rev Cell Mol Biol 2009; 276:1 - 48
- Heyl A, Werner T, Schmülling T. Hedden P, Thomas S. Cytokinin metabolism and signal transduction. Plant hormone signaling, Annual Plant Reviews 2006; Blackwell Publishing Ltd 93 - 123
- Perilli S, Moubayidin L, Sabatini S. The molecular basis of cytokinin function. Curr Opin Plant Biol 2010; 13:21 - 26
- Coba de la Peña T, Cárcamo CB, Almonacid L, Zaballos A, Lucas MM, Balomenos D, et al. A salt stress-responsive cytokinin receptor homologue isolated from Medicago sativa nodules. Planta 2008; 227:769 - 779
- Lohar DP, Sharopova N, Endre G, Penuela S, Samac D, Town C, et al. Transcript analysis of early nodulation events in Medicago truncatula. Plant Physiol 2006; 140:221 - 234
- Merchan F, de Lorenzo L, Rizzo SG, Niebel A, Manyani H, Frugier F, et al. Identification of regulatory pathways involved in the reacquisition of root growth after salt stress in Medicago truncatula. Plant J 2007; 51:1 - 17
- Mähönen AP, Bishopp A, Higuchi M, Nieminen KM, Kinoshita K, Tormakangas K, et al. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 2006; 311:94 - 98
- Nieminen K, Immanen J, Laxell M, Kauppinen L, Tarkowski P, Dolezal K, et al. Cytokinin signaling regulates cambial development in poplar. Proc Natl Acad Sci USA 2008; 105:20032 - 20037
- Muñiz LM, Royo J, Gómez E, Baudot G, Paul W, Hueros G. Atypical response regulators expressed in the maize endosperm transfer cells link canonical two component systems and seed biology. BMC Plant Biol 2010; 10:84
- Yonekura-Sakakibara K, Kojima M, Yamaya T, Sakakibara H. Molecular characterization of cytokinin-responsive histidine kinases in maize. Differential ligand preferences and response to cis-zeatin. Plant Physiol 2004; 134:1654 - 1661
- Romanov GA, Spíchal L, Lomin SN, Strnad M, Schmülling T. A live cell hormone-binding assay on transgenic bacteria expressing a eukaryotic receptor protein. Anal Biochem 2005; 347:129 - 134
- Spíchal L, Rakova NY, Riefler M, Mizuno T, Romanov GA, Strnad M, et al. Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay. Plant Cell Physiol 2004; 45:1299 - 1305
- Hirano K, Aya K, Hobo T, Sakakibara H, Kojima M, Shim RA, et al. Comprehensive transcriptome analysis of phytohormone biosynthesis and signaling genes in microspore/pollen and tapetum of rice. Plant Cell Physiol 2008; 49:1429 - 1450
- Papon N, Clastre M, Andreu F, Gantet P, Rideau M, Crèche J. Expression analysis in plant and cell suspensions of CrCKR1, a cDNA encoding a histidine kinase receptor homologue in Catharanthus roseus (L.). G Don J Exp Bot 2002; 53:1989 - 1990
- Coba de la Peña T, Cárcamo CB, Almonacid L, Zaballos A, Lucas MM, Balomenos D, et al. A cytokinin receptor homologue is induced during root nodule organogenesis and senescence in Lupinus albus L. Plant Physiol Biochem 2008; 46:219 - 225
- Kumar V, Mills DJ, Anderson JD, Mattoo AK. An alternative agriculture system is defined by a distinct expression profile of select gene transcripts and proteins. Proc Natl Acad Sci USA 2004; 101:10535 - 10540
- Asakura Y, Hagino T, Ohta Y, Aoki K, Yonekura-Sakakibara K, Deji A, et al. Molecular characterization of His-Asp phosphorelay signaling factors in maize leaves: implications of the signal divergence by cytokinin-inducible response regulators in the cytosol and the nuclei. Plant Mol Biol 2003; 52:331 - 341
- Sugawara H, Kawano Y, Hatakeyama T, Yamaya T, Kamiya N, Sakakibara H. Crystal structure of the histidine-containing phosphotransfer protein ZmHP2 from maize. Protein Sci 2005; 14:202 - 208
- Sugawara H, Yamaya T, Sakakibara H. Crystallization and preliminary X-ray diffraction study of the histidine-containing phosphotransfer protein ZmHP1 from maize. Acta Crystallogr Sect F Struct Biol Cryst Commun 2005; 61:366 - 368
- de Lorenzo L, Merchan F, Blanchet S, Megias M, Frugier F, Crespi M, et al. Differential expression of the TFIIIA regulatory pathway in response to salt stress between Medicago truncatula genotypes. Plant Physiol 2007; 145:1521 - 1532
- Tran LS, Urao T, Qin F, Maruyama K, Kakimoto T, Shinozaki K, et al. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought and salt stress in Arabidopsis. Proc Natl Acad Sci USA 2007; 104:20623 - 20628
- Maul P, Bausher M, McCollum G, Mozoruk J, Niedz R. CsHPt1, a putative histidine containing phosphotransmitter protein induced during early somatic embryogenensis in Valencia sweet orange. Plant Science 2006; 170:44 - 53
- Chefdor F, Bénédetti H, Depierreux C, Delmotte F, Morabito D, Carpin S. Osmotic stress sensing in Populus: components identification of a phosphorelay system. FEBS Lett 2005; 580:77 - 81
- Ma QH, Tian B. Characterization of a wheat histidine-containing phosphotransfer protein (HP) that is regulated by cytokinin-mediated inhibition of leaf senescence. Plant Sci 2005; 168:1507 - 1514
- Brenner WG, Romanov GA, Köllmer I, Bürkle L, Schmülling T. Immediate-early and delayed cytokinin response genes of Arabidopsis thaliana identified by genome-wide expression profiling reveal novel cytokinin-sensitive processes and suggest cytokinin action through transcriptional cascades. Plant J 2005; 44:314 - 333
- Kiba T, Naitou T, Koizumi N, Yamashino T, Sakakibara H, Mizuno T. Combinatorial microarray analysis revealing Arabidopsis genes implicated in cytokinin responses through the His→Asp Phosphorelay circuitry. Plant Cell Physiol 2005; 46:339 - 355
- Jackson D, Hake S. Control of phyllotaxy in maize by the abphyl1 gene. Development 1999; 126:315 - 323
- Giulini A, Wang J, Jackson D. Control of phyllotaxy by the cytokinin-inducible response regulator homologue ABPHYL1. Nature 2004; 430:1031 - 1034
- Lee BH, Johnston R, Yang Y, Gallavotti A, Kojima M, Travencolo BA, et al. Studies of aberrant phyllotaxy1 mutants of maize indicate complex interactions between auxin and cytokinin signaling in the shoot apical meristem. Plant Physiol 2009; 150:205 - 216
- Muñiz LM, Royo J, Gómez E, Barrero C, Bergareche D, Hueros G. The maize transfer cell-specific type-A response regulator ZmTCRR-1 appears to be involved in intercellular signalling. Plant J 2006; 48:17 - 27
- Deji A, Sakakibara H, Ishida Y, Yamada S, Komari T, Kubo T, et al. Genomic organization and transcriptional regulation of maize ZmRR1 and ZmRR2 encoding cytokinin-inducible response regulators. Biochim Biophys Acta 2000; 1492:216 - 220
- Sakakibara H, Hayakawa A, Deji A, Gawronski SW, Sugiyama T. His-Asp phosphotransfer possibly involved in the nitrogen signal transduction mediated by cytokinin in maize: molecular cloning of cDNAs for two-component regulatory factors and demonstration of phosphotransfer activity in vitro. Plant Mol Biol 1999; 41:563 - 573
- Sakakibara H, Suzuki M, Takei K, Deji A, Taniguchi M, Sugiyama T. A response-regulator homologue possibly involved in nitrogen signal transduction mediated by cytokinin in maize. Plant J 1998; 14:337 - 344
- Taniguchi M, Kiba T, Sakakibara H, Ueguchi C, Mizuno T, Sugiyama T. Expression of Arabidopsis response regulator homologs is induced by cytokinins and nitrate. FEBS Lett 1998; 429:259 - 262
- Hirose N, Makita N, Kojima M, Kamada-Nobusada T, Sakakibara H. Overexpression of a type-A response regulator alters rice morphology and cytokinin metabolism. Plant Cell Physiol 2007; 48:523 - 539
- Cheng X, Jiang H, Zhang J, Qian Y, Zhu S, Cheng B. Overexpression of type-A rice response regulators, OsRR3 and OsRR5, results in lower sensitivity to cytokinins. Genet Mol Res 2010; 9:348 - 359
- Cortizo M, Alvarez JM, Rodríguez A, Fernández B, Ordás RJ. Cloning and characterization of a type-A response regulator differentially expressed during adventitious shoot formation in Pinus pinea L.. J Plant Physiol 2010; 167:1023 - 1026
- Camacho Y, Martinez-Castilla L, Fragoso S, Vázquez S, Martínez-Barajas E, Coello P. Characterization of a type A response regulator in the common bean (Phaseolus vulgaris) in response to phosphate starvation. Physiol Plant 2008; 132:272 - 282
- Papon N, Oudin A, Vansiri A, Rideau M, Chénieux JC, Créche J. Differential expression of two type-A response regulators in plants and cell cultures of Catharanthus roseus (L.). G Don J Exp Bot 2003; 54:1793 - 1795
- Papon N, Clastre M, Gantet P, Rideau M, Chénieux °C, Créche J. Inhibition of the plant cytokinin transduction pathway by bacterial histidine kinase inhibitors in Catharanthus roseus cell cultures. FEBS Lett 2003; 537:101 - 105
- Sitte P, Weiler EW, Kadereit JW, Bresinsky A, Körner C. Strasburger, Lehrbuch der Botanik 2002; Berlin Spektrum Akademischer Verlag
- Hren M, Nikolic P, Rotter A, Blejec A, Terrier N, Ravnikar M, Dermastia M, Gruden K. ‘Bois noir’ phytoplasma induces significant reprogramming of the leaf transcriptome in the field grown grapevine. BMC Genomics 2009; 10:460
- Imin N, Goffard N, Mizamidin M, Rolfe BG. Genome-wide transcriptional analysis of super-embryogenic Medicago truncatula explant cultures. BMC Plant Biol 2008; 8:110