Abstract
Jasmonate (JA) inhibits root growth of Arabidopsis thaliana seedlings. The mutation in COI1 that plays a central role in JA signaling displays insensitivity to JA inhibition of root growth. To dissect JA signaling pathway, we recently isolated one mutant named psc1, which partially suppresses coi1 insensitivity to JA inhibition of root growth. As we identified the PSC1 gene as an allele of DWF4 that encodes a key enzyme in brassinosteroid (BR) biosynthesis, we hypothesized and demonstrated that BR is involved in JA signaling and negatively regulates JA inhibition of root growth. In our Plant Physiology paper, we analyzed effects of psc1 or exogenous BR on the inhibition of root growth by JA. Here we show that treatment with brassinazole (Brz), a BR biosynthesis inhibitor, increased JA sensitivity in both coi1-2 and wild type, which further confirms that BR negatively regulates JA inhibition of root growth. Since effects of psc1, Brz, and exogenous BR on JA inhibition of root growth were mild, we suggests that BR negatively finely regulates JA inhibition of root growth in Arabidopsis.
Jasmonate (JA) regulates many plant developmental processes and stress responses.Citation1,Citation2 COI1 plays a central role in JA signaling and is required for all JA responses in Arabidopsis.Citation3,Citation4 coi1-1, a strong mutation in COI1, is male sterile and exhibits loss of all JA responses tested to date, such as JA inhibition of root growth, the expression of JA-induced genes, and susceptibility to insect attack and pathogen infection, and coi1-2, a weak mutant of COI1, shows similar JA responses to coi1-1 except for partially fertile that makes it able to produce a small quantity of seeds.Citation5
To investigate COI1-mediated JA responses and dissect JA signaling pathway, we conducted genetic screens for suppressors of coi1-2. Previously, we identified cos1 that completely suppresses coil-2 insensitive to JA.Citation6 Recently, we isolated the psc1 mutant that partially suppresses coi1-2 insensitivity to JA, and found that PSC1 is an allele of DWF4.Citation7
Since the DWF4 gene encodes a key enzyme in brassinosteroid (BR) biosynthesis,Citation8 we hypothesized that BR is involved in JA signaling. By physiological analysis, we showed that psc1 partially restored JA inhibition of root growth in coi1-2 background and displayed JA hypersensitivity in wild-type COI1 background, the effects of psc1 were eliminated by exogenous BR, and that exogenous BR could attenuated JA inhibition of root growth in wild type. These findings demonstrated that BR is involved in JA signaling and indicated that BR negatively regulates JA inhibition of root growth.
BR is a family of polyhydroxylated steroid hormones involved in many aspects of plant growth and development. The BR-deficient mutants exhibited severely retarded growth that was able to be rescued by exogenous BR.Citation9 Brassinazole (Brz) is a BR biosynthesis inhibitor. The Arabidopsis seedlings treated with Brz displayed a BR deficient-mutant-like phenotype, which could be elimilated by exogenous BR.Citation10
To determine wether treatment with Brz affects JA inhibition of root growth, the seedlings of wild type and coi1-2 were grown in MS medium supplemented with MeJA and/or Brz. As shown in , the relative root length was obviously reduced in both coi1-2 and wild type when treated with Brz relative to without Brz, indicating that the repression of BR biosynthesis by Brz could increase JA sensitivity. These results further confirm BR negatively regulates JA inhibition of root growth.
It has been demonstrated that JA connects with other plant hormones including auxin, ethylene, abscisic acid, salicylic acid and gibberellin to form complex regulatory networks modulating plant developmental and stress responses.Citation11–Citation15 We found that BR negatively regulates JA inhibition of root growth, suggesting that a cross talk between JA and BR exists in planta, which extends our understandings on the JA signal transduction.
COI1 is a JA receptorCitation16 and DWF4 catalyzes the rate-limiting step in BR-biosynthesis pathway.Citation8 We found that JA inhibits DWF4 expression, this inhibition was dependent on COI1,Citation7 indicating that DWF4 is downregulated by JA and is located downstream of COI1 in the JA signaling pathway.
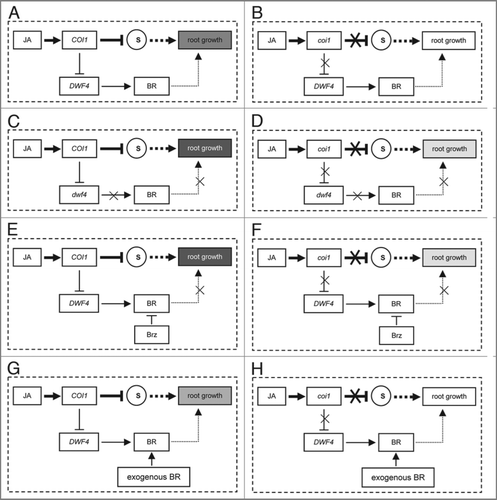
Since the effects of psc1, Brz, and exogenous BR on JA inhibition of root growth were mild, and the DWF4 expression was partially repressed by JA (Ren et al. 2009, ), we suggest that BR negatively finely regulates JA inhibition of root growth, and propose a model for these regulations. As shown in , JA signal passes COI1 repressing substrates, such as JAZs,Citation17,Citation18 i.e., JA activates degradation of substrates via SCFCOI1-26S proteasome,Citation16–Citation18 whereas substrates positively regulate root growth through other regulators. JA also partially inhibits DWF4 expression through COI1, reducing BR that is required for root growth.Citation7,Citation9 Mutation in COI1 interrupts JA signaling for failing in degradation of substrates and repression of DWF4 as well, resulting in JA-insensitivity (). However, mutation in DWF4 or treatment with Brz causes a reduction in BR, which affects root growth, leading to JA-hypersensitivity in wild-type COI1 background () and partial restoration of JA sensitivity in coi1-2 background (). Whereas, an application of exogenous BR could eliminate the effect of BR reduction resulted from repression of DWF4 by JA on root growth, attenuating JA sensitivity in wild type (). Because the inhibition of DWF4 expression by JA is dependent on COI1, the coi1 mutant treated with exogenous BR do not show alteration in JA sensitivity ().
Figures and Tables
Figure 1 Effect of Brz on JA inhibition of root growth. Brz increased JA inhibition of root growth in both coi1-2 and wild type (WT). Root length of 7-day-old seedlings grown in MS medium containing 0, 5 and 10 μM MeJA without (−) or with (+) 0.5 μM Brz was expressed as a percentage of root length in MS without (−) or with (+) 0.5 µM Brz. Error bars represent SE (n > 30).
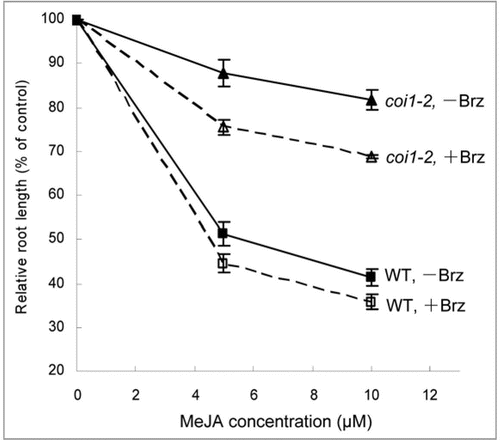
Figure 2 A model for that BR negatively finely regulates JA inhibition of root growth in Arabidopsis. (A–D) Treatment with JA in wild type (A), coi1-2 (B), psc1 (C) and psc1coi1 (D). (E and F) Treatments with JA and Brz in wild type (E) and coi1-2 (F). (G and H) Treatments with JA and exogenous BR in wild type (G) and coi1-2 (H). Arrows indicate positive regulation or enhancement, whereas blunted lines indicate repression or negative regulation. Crosses indicate interruption or impairment. The letter “S” indicates substrates of SCFCOI1. Thicker arrows and blunted lines represent the central JA signaling pathway regulating JA inhibition of root growth. Broken arrows represent JA signaling pathway in which other regulators are involved. The intensity of gray boxes represents the degree of JA inhibition on root growth.
Acknowledgements
This research was funded by the National Science Foundation of China, the National Basic Research 973 Program of China and the Scientific Research Fund of Hunan Provincial Education Department.
Addendum to:
Related Research Data
References
- Shan XY, Wang ZL, Xie D. Jasmonate signal pathway in Arabidopsis. J Integr Plant Biol 2007; 49:81 - 86
- Browse J. Jasmonate passes muster: a receptor and targets for the defense hormone. Annu Rev Plant Biol 2009; 60:183 - 205
- Feys BJF, Benedetti CE, Penfold CN, Turner JG. Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistant to a bacterial pathogen. Plant Cell 1994; 6:751 - 759
- Xie DX, Feys BF, James S, Nieto-Rostro M, Turner JG. COI1: An Arabidopsis gene required for jasmonate-regulated defense and fertility. Science 1998; 280:1091 - 1094
- Xu L, Liu F, Lechner E, Genschik P, Crosby WL, Ma H, et al. The SCFCOI1 ubiquitin-ligase complexes are required for jasmonate response in Arabidopsis. Plant Cell 2002; 14:1919 - 1935
- Xiao S, Dai L, Liu L, Wang Z, Peng W, Xie D. COS1: An Arabidopsis coronatine insensitive1 suppressor essential for regulation of jasmonate-mediated plant defense and senescence. Plant Cell 2004; 16:1132 - 1142
- Ren C, Han C, Peng W, Huang Y, Peng Z, Xiong X, et al. A leaky mutation in DWARF4 reveals an antagonistic role of brassinosteroid in the inhibition of root growth by jasmonate in Arabidopsis. Plant Physiol 2009; 151:1412 - 1420
- Choe S, Dilkes BP, Fujioka S, Takatsuto S, Sakurai A, Feldmann KA. The DWF4 gene of Arabidopsis encodes a cytochrome P450 that mediates multiple 22α-hydroxylation steps in brassinosteroid biosynthesis. Plant Cell 1998; 10:231 - 243
- Belkhadir Y, Chory J. Brassinosteroid Signaling: A paradigm for steroid hormone signaling from the cell surface. Science 2006; 314:1410 - 1411
- Asami T, Min YK, Nagata N, Yamagishi K, Takatsuto S, Fujioka S, et al. Characterization of brassinazole, a triazole-type brassinosteroid biosynthesis inhibitor. Plant Physiol 2000; 123:93 - 99
- Ellis C, Turner JG. A conditionally fertile coi1 allele indicates cross-talk between plant hormone signalling pathways in Arabidopsis thaliana seeds and young seedlings. Planta 2002; 215:549 - 556
- Tiryaki I, Staswick PE. An Arabidopsis mutant defective in jasmonate response is allelic to the auxin-signaling mutant axr1. Plant Physiol 2002; 130:887 - 894
- Lorenzo O, Piqueras R, Sanchez-Serrano JJ, Solano R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 2003; 15:165 - 178
- Ren CM, Zhu Q, Gao BD, Ke SY, Yu WC, Xie DX, et al. Transcription factor WRKY70 displays important but no indispensable roles in jasmonate and salicylic acid signaling. J Integr Plant Biol 2008; 50:630 - 637
- Cheng H, Song S, Xiao L, Soo HM, Cheng Z, Xie D, et al. Gibberellin acts through jasmonate to control the expression of MYB21, MYB24 and MYB57 to promote stamen filament growth in Arabidopsis. PLoS Genet 2009; 5:1000440
- Yan J, Zhang C, Gu M, Bai Z, Zhang W, Qi T, et al. The Arabidopsis CORONATINE INSENSITIVE1 protein is a jasmonate receptor. Plant Cell 2009; 21:2220 - 2236
- Chini A, Fonseca S, Fernandez G, Adie B, Chico JM, Lorenzo O, et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007; 448:666 - 671
- Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, et al. JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling. Nature 2007; 448:661 - 665