Abstract
Using a microarray-based approach the root meristem of maize was profiled.6 Three populations were characterized; the root cap, the quiescent center and the proximal meristem. From these data we here consider the central role of auxin in defining and establishing the root stem cell niche.
The plant growth regulator, auxin plays an important role in integrating the development of the root apical meristem. Auxin does so, in part by specifying the location of the stem cell niche (SCN), which develops both temporally and spatially co-incident with the establishment of the auxin maximum in the root.Citation1–Citation3 As a consequence of being located within the niche, cells take on stereotypical stem cell characteristics, including an apparent ability for unlimited proliferation, self-maintenance and self-renewal.Citation4,Citation5 In Angiosperm roots the SCN is populated by two sub-populations of cells designated the quiescent center (QC), and the proximal meristem (PM). A continuous communication occurs between these two sister stem cell populations, with this cross talk likely influenced by the positioning of the auxin maximum.Citation6 The activities of the niche and signaling between the QC and PM can be perturbed by subjecting roots to environmental extremes, by surgical removal of the root cap (decapping), or by treating roots with ascorbate.7 Here we consider auxin-related/-regulated transcription in the SCN, specifically noting differences and similarities between the two adjacent sister cell populations comprising the QC and the PM.
Expression of Auxin-Transport Genes Differs in the QC and PM
It has already been well-established that the PIN/AUX family of proteins plays an important role in positioning the auxin maximum in the root.Citation8,Citation9 In maize we found that the gradient of auxin along the root axis10 paralleled the expression of three putative auxin-efflux carrier (ZmPIN) genes, which increased in expression from the elongation zone (EZ) to the root cap (RC) (). These observations lead us to suggest that the distribution of these proteins, and the resulting auxin maximum, may be related and understood in terms of the levels and spatial expression patterns of auxin efflux carrier transcripts along the root axis. In contrast, for ZmAUX1, a maize auxin influx carrier, expression was greatest in the QC (), increasing markedly when the niche is perturbed (decapping) (), thereby suggesting that this increased expression specifically in the QC may be associated with a re-establishment of the auxin maximum and the niche.
Auxin Mediates Niche Formation Through AUXIN RESPONSE FACTORS (ARFs)
In ARF (AUXIN RESPONSE FACTOR) mutants (arf5/mp or arf5arf7) the expression of PLETHORA (PLT1) is reduced, thereby preventing niche establishment during embryogenesis,Citation11 and also suggesting that ARFs likely play an important role in early steps of auxin-regulated SCN establishment. Previously we reported that homologues of the Arabidopsis ARF family are expressed, some at high levels (e.g., ZmARF7a and ZmARF5/MP), in the QC portion of the maize SCN (), and that these genes can show dynamic expression ().Citation6 Perturbing the niche (decapping) results in ZmARF7a’s decreased expression in the QC, which parallels increases in cell division activity in the QC (), suggesting ZmARF7a might act as a QC marker. Also, the expression of such a large number of ARF genes in the QC points to the possibility of multiple pathways through which members of this family could influence niche specification and maintenance. Whereas all of the ARF family members that we measured are expressed in the QC, most (except ZmARF7a and ZmARF5/MP) are expressed at higher levels in the root cap, suggesting that these genes may also mediate auxin-related events in the root cap.
PLT genes are essential for QC specification and stem cell activity.Citation11,Citation12 For the intact root we report a greater (4-fold) increase in ZmPLT expression specifically in the QC compared to the adjacent PM, and a 160-fold increase compared to the elongation zone (EZ) (Table 3).Citation6 Moreover, decapping results in a greater fold and more sustained increase in ZmPLT expression in the QC than in the PM (). These two results thus support a role for ZmPLT in QC specification in maize, as earlier suggested for Arabidopsis by Aida et al.Citation11 Interestingly, in the intact maize root the spatial expression of ZmPLT correlates with that of ZmAFR7a and ZmARF5/MP, supporting the view that ZmPLT might act downstream of ZmARF5/MP and/or ZmARF7a, as also suggested for Arabidopsis.Citation11 However, unlike Aida et al.Citation11 whose work with knock-outs suggests an ARF7 induction of the expression of PLT1, we conclude differently for ZmARF7a. Our decapping experiments show that a reduction in ZmARF7a expression is not accompanied by a reduction in ZmPLT expression (), which suggests that ZmARF7a might suppress ZmPLT expression, or that ZmPLT might be not regulated by ZmARF7a in the native situation.
Ascorbate Affects the Location of the Auxin Maximum in the Maize Root
Based on the fact that applications of ascorbate perturb the niche, or may even abolish it, and on earlier biochemical results suggesting a linkage between auxin and the oxidized redox status of the niche,Citation10,Citation13 we explored further a possible linkage between redox status and the positioning of the auxin maximum. Treating intact maize roots with 0.5 mM ascorbate (AA) altered the location of the auxin maximum and this was paralleled by a changed status of the niche (). Twenty-four h after ending the AA treatment the auxin maximum in the niche has diminished and it has also been displaced from its pre-treatment location in the columella initials/distal QC (). After an additional 24 h (48 h after ending the AA treatment), a new auxin maximum reappears within the (reforming) niche, over the region of the original QC (). We conclude that AA-induced perturbation of the niche is associated with changes in the accumulation of auxin, and with changes in the location of the auxin maximum, thereby supporting the suggestion of a linkage of the activities of auxin and redox-associated genes in niche specification and maintenance.Citation6 A similar conclusion was recently advanced by De Tullio et al.Citation7 for the SCN in Arabidopsis roots.
In conclusion, we suggest that integration of activities in the root SCN is largely mediated by the positioning of the auxin maximum, and that this underlies the differences and similarities in auxin-related/-regulated events in the QC and PM.
Abbreviations
| AA | = | ascorbic acid |
| ARF | = | auxin response factor |
| AUX1 | = | auxin influx transporter |
| EZ | = | elongation zone |
| PIN | = | pin-formed |
| PLT | = | plethora |
| PM | = | proximal meristem |
| QC | = | quiescent center |
| RC | = | root cap |
| SCN | = | stem cell niche |
| Zm | = | Zea mays |
Figures and Tables
Figure 1 (A-D) immunolocalization of auxin in Zea mays roots treated for 24 h with ascorbic acid (AA ) and then examined 24 and 48 h after ending the AA treatment. Arrows point to the position of the auxin maximum. Immunolocalization of auxin was done as described by Jiang et al.Citation10
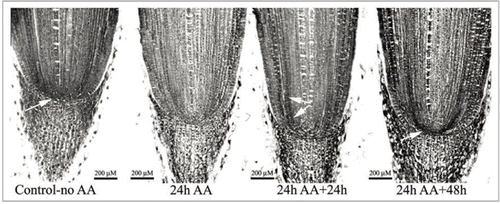
Table 1 Auxin-transport gene expression in the maize root tip
Table 2 Relative levels of expression of auxin-related genes in the proximal meristem (PM) and quiescent center (QC), at various times (0, +6, +20, +32 h) after removal of the root cap and activation of the QCTable Footnotea
Table 3 AFRs and PLT expression in the maize root tipTable Footnotea
Addendum to:
Related Research Data
References
- Friml J, Vieten A, Sauer M, Weijers D, Schwarz H, Hamann T, et al. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003; 426:147 - 153
- Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoye T, Malamy J, et al. An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell 1999; 99:463 - 472
- Petersson SV, Johansson AI, Kowalczyk M, Makoveychuk A, Wang JY, Moritz T, et al. An auxin gradient and maximum in the Arabidopsis root apex shown by high-resolution cell-specific analysis of IAA distribution and synthesis. Plant Cell 2009; 21:1659 - 1668
- Barlow P. Potten CS. Stem cells and founder zones in plants, particularly their roots. Stem Cells 1997; London Academic 29 - 57
- Ivanov VB. The problem of stem cells in plants. Russ J Dev Biol 2003; 34:205 - 212
- Jiang K, Zhu T, Diao Z, Huang H, Feldman LJ. The maize root stem cell niche: a partnership between two sister cell populations. Planta 2010; 231:411 - 424
- De Tullio MC, Jiang K, Feldman LJ. Redox regulation of root apical meristem organization: connecting root development to its environment. Plant Physiol Biochem (in press)
- Fischer U, Ikeda Y, Ljung K, Serralbo O, Singh M, Heidstra H, et al. Vectorial information for Arabidopsis planar polarity is mediated by combined AUX1, EIN2 and GNOM activity. Curr Biol 2006; 16:2143 - 2149
- Grieneisen VA, Xu J, Marée AFM, Hogeweg P, Scheres B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007; 449:1008 - 1013
- Jiang K, Meng YL, Feldman LJ. Quiescent center formation in maize roots is associated with an auxin-regulated oxidizing environment. Development 2003; 130:1429 - 1438
- Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, et al. The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 2004; 119:109 - 120
- Galinha C, Hofhuis H, Juijten M, Willemsen V, Blilou I, Heidstra R, et al. PLETHORA proteins as dose-dependent master regulators of Arabidopsis root development. Nature 2007; 449:1053 - 1057
- Kerk NM, Feldman LJ. A biochemical model for the initiation and maintenance of the quiescent center: implications for organization of root meristems. Development 1995; 121:2825 - 2833