Abstract
The phytohormone abscisic acid (ABA) is known to be a negative regulator of legume root nodule formation. By screening Lotus japonicus seedlings for survival on an agar medium containing 70 μM ABA, we obtained mutants that not only showed increased root nodule number, but also enhanced nitrogen fixation. The mutant was designated enf1 (enhanced nitrogen fixation 1) and was confirmed to be monogenic and incompletely dominant. In long-term growth experiments with M. loti, although some yield parameters were the same for both enf1 and wild-type plants, both the dry weight and N content of 100 seeds and entire enf1 plants were significantly larger compared than those traits in wild-type seeds and plants. The augmentation of the weight and N content of the enf1 plants most likely reflects the increased N supplied by the additional enf1 nodules and the concomitant increase in N fixation activity. We determined that the endogenous ABA concentration and the sensitivity to ABA of enf1 were lower than that of wild-type seedlings. When wild-type plants were treated with abamine, a specific inhibitor of 9-cis-epoxycarotenoid dioxygenase (NCED), which results in reduced ABA content, the N fixation activity of abamine-treated plants was elevated to the same levels as enf1. We also determined that production of nitric oxide (NO) in enf1 nodules was decreased. We conclude that endogenous ABA concentration not only regulates nodulation, but also nitrogen fixation activity by decreasing NO production in nodules.
Phytohormones are known to be important for regulating the number of nodules established on the root of legumes.Citation1 For example, ethylene is a well-known negative regulator of nodulation, influencing the earliest stages from the perception of Nod factor to the growth of infection threads.Citation2–Citation4 In contrast, cytokinin is a positive regulator of nodulation. The cytokinin insensitive mutant hit1 (loss-of-function) of Lotus japonicus and the snf2 (gain-of-function) mutants of Medicago truncatula provide genetic evidence demonstrating that cytokinin plays a critical role in the activation of nodule primordia.Citation5–Citation7
Abscisic acid (ABA), added at concentrations that do not affect plant growth, also negatively regulates nodulation in some legumes.Citation8–Citation11 Recently, Medicago truncatula overexpressing abi1-1, a gene that encodes a mutated protein phosphatase of the type IIC class derived from Arabidopsis and that suppresses the ABA signaling pathway,Citation12,Citation13 was shown to exhibit ABA insensitivity as well as a hypernodulating phenotype.Citation14
In this study, we isolated a Lotus japonicus (Miyakojima MG20) mutant that showed an increased root nodule phenotype and a lowered sensitivity to ABA, and proceeded to carry out its characterization. This mutant, named enf1 (enhanced nitrogen fixation 1) exhibit enhanced symbiotic N fixation activity. Most legume N fixation activity mutants, such as ign1, sen1 and sst1, are Fix-.Citation15–Citation17
At first, to obtain ABA-insensitive or low-sensitive mutants of Lotus japonicus, we treated Miyakojima MG20 with EMS to induce base substitutions randomly in the genome. M3 seeds were sown on an agar-solidified medium containing 70 µM ABA, a concentration that inhibits the germination of wild-type MG20 seeds. M4 plants obtained by the screening were inoculated with rhizobia (Mesorhizobium loti MAFF303099), and the number of nodules per plant was counted 35 days after inoculation (DAI). Plant No. 12 not only formed more root nodules than did the wild-type MG20 plants, but surprisingly it also exhibited increased nitrogen fixation activity per plant. Both mutant phenotypes were stably inherited in the M4 and M5 generation. Back-crossing mutant No. 12 to wild-type MG20 yielded 153 F2 progeny from which a line that showed the highest N fixation activity and more nodules per plant was derived. This line was designated enf1 (enhanced nitrogen fixation 1).
At 28 DAI, the number of nodules formed on enf1 roots was approximately 1.7 times greater than that of MG20, and the N fixation activity per enf1 plant was elevated 1.8 times over that of the wildtype plants. Because the N fixation activity per unit of enf1 nodule weight was also increased 1.7 times, we concluded that the increased N fixation activity was not solely due to the enhanced number of root nodules.
The endogenous ABA concentration and the sensitivity to ABA of enf1 were lower than those of wild-type seedlings. ABA is believed to regulate early nodulation stages negatively by inhibiting Nod factor signaling, bacterial infection, and nodule initiation.Citation14,Citation18 Elongated ITs were more common in enf1 root hairs at later stages of development (8–12 DAI). Furthermore, ITs were detected in nodule primordia more frequently in enf1 compared to MG20. These results suggest that the earliest stages of nodule development are not as strongly inhibited in enf1 as they are in wild-type MG20.
Because enf1 had a low endogenous ABA concentration, we hypothesized that the decrease in ABA concentration caused the elevation of N fixation activity. To test this hypothesis, we treated wild-type plants at 28 DAI with 20 µM abamine, a specific inhibitor of ABA synthesis.Citation19 After a three day-treatment period, acetylene reduction activity was measured. Such short treatment periods of abamine are not expected to induce new nodule development. Wild-type plants treated with abamine had a reduced endogenous ABA concentration in roots, to about one-fourth of the level of control plants. However, N fixation activity was elevated to about 170% over the non-treated controls ( and B). This result phenocopies enf1, which shows decreased endogenous ABA concentration as well as elevated N fixation activity. These results strongly suggest that the decrease in endogenous ABA concentration in enf1 was responsible for the increased levels of N fixation activity. Applying 0.5 µM ABA did not result in a further increase in N fixation activity even though the endogenous ABA concentrations are presumed to increase ( and B).
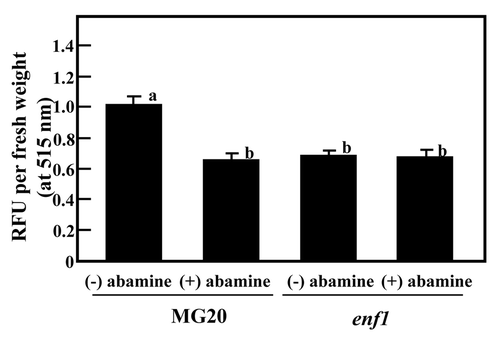
Nitric oxide (NO) is known as a strong inhibitor of N fixation activity,Citation20 as well as a signal component in ABA signaling pathway.Citation21,Citation22 NO production in root nodules formed by enf1 21 DAI and 28 DAI was examined by using the fluorescent dye diaminofluorescein-FM (DAF-FM), a NO specific detector, and relative fluorescence unit (RFU) values were estimated. The RFU values of enf1 nodules 21 DAI were clearly decreased compared with that of MG20; this trend was more obvious at 28 DAI. Moreover, the effect of reduced ABA concentration caused by treatment with abamine on NO production was analyzed (). When nodules formed on the roots of 28-d-old plants were treated, the RFU value of the enf1 mutant was almost the same for (−) abamine and (+) abamine-treated, whereas, the RFU value of abamine-treated MG20 plants was significantly reduced compared to untreated MG20 (). These results strongly suggest that decreased production of NO caused by the low concentration of ABA in enf1 nodules was responsible for the increase in N fixation activity.
Until now, the majority of symbiotic mutants that have been described represents loss of or defects in root nodule formation.Citation6,Citation23,Citation24 Many of these mutants induce nodules that are Fix-.Citation15–Citation17 Although reports of mutants that show increased root nodule numberCitation25–Citation28 or spontaneous root nodule formation exist,Citation7,Citation29 reports concerning mutations where N fixation activity is elevated without deleterious effects on plant growth and development are limited. One exception is the L. japonicus rdh1 mutant, which also exhibits a hypernodulation and enhanced nitrogen fixation phenotype.Citation30
In this report, we have shown that mutating the ENF1 gene leads to an elevation of N fixation activity without accompanying adverse growth effects. In long-term growth experiments, some yield parameters were the same for both enf1 and wild-type plants, but both the dry weight and N content of 100 seeds and entire enf1 plants were significantly larger compared to those parameters in wild-type seeds and plants. These results strongly suggest that more nitrogen is fixed in the enf1 mutant than in wild-type plants. Therefore, this gene should be an important target for molecular breeding. We have determined that ENF1 gene is inherited in a monogenic and incompletely dominant manner. Our future work will identify the gene responsible for these positive growth effects.
Figures and Tables
Figure 1 Effects of ABAconcentration on nitrogen fixation activity. M. loti-inoculated plants were grown for 28 days on vermiculite-filled pots supplied with B & D medium. Plant roots 28 DAI were treated with 0.5 µM ABA, 20 µM abamine, with both ABA and abamine, or were untreated (B & D medium control), respectively, for 3 days. (A) ARA per nodule weight. (B) ABA concentration in root. At least 15 plants were used in acetylene reduction assay. Four different plants were used for the measurement of ABA concentration and 3 repeats were performed. Error bars indicate the standard error, and the significance of differences between untreated control and treated values was determined by the two-tailed multiple t-test with Bonferroni correction following ANOVA (three comparisons in four groups), *p < 0.05, **p < 0.01.
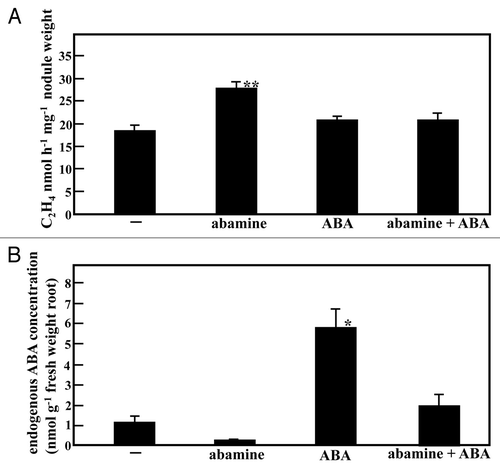
Figure 2 NO production in nodules. Quantification of nitric oxide in nodules that were treated with abamine. Nodules on the root of 28-day-old plants were treated with 20 µM abamine for 3 days. Relative fluorescent units (RFU) per nodule fresh weight at 515 nm, normalized against MG20 plants, are shown. The data represent the average ± standard error of 3 independent experiments derived from nodules of 6 to 8 plants. The significance of differences among the four groups was determined by the two-tailed multiple t-test with Bonferroni correction following ANOVA (six comparisons in four groups) and the different letters refer to significant differences at p < 0.01.
Addendum to:
References
- Hirsch AM, Fang Y. Plant hormones and nodulation: What's the connection?. Plant Mol Biol 1994; 26:5 - 9
- Nukui N, Ezura H, Yuhashi K-I, Yasuta T, Minamisawa K. Effects of ethylene precursor and inhibitors for ethylene biosynthesis and perception on nodulation in Lotus japonicus and Macroptilium atropurpureum. Plant Cell Physiol 2000; 41:893 - 897
- Ding Y, Kalo P, Yendrek C, Sun J, Liang Y, Marsh JF, et al. Abscisic acid coordinates Nod Factor and cytokinin signaling during the regulation of nodulation in Medicago truncatula. Plant Cell 2008; 20:2681 - 2695
- Ma W, Guinel FC, Glick BR. Rhizobium leguminosarum biovar viciae 1-aminocyclopropane-1-carboxylate deaminase promotes nodulation of pea plants. Appl Environ Microbiol 2003; 69:4396 - 4402
- Gonzalez-Rizzo S, Crespi M, Frugier F. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 2006; 18:2680 - 2693
- Murray JD, Karas BJ, Sato S, Tabata S, Amyot L, Szczyglowski K. A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science 2007; 315:101 - 104
- Tirichine L, Sandal N, Madsen LH, Radutoiu S, Albrektsen AS, Sato S, et al. A gain-of-function mutation in a cytokinin receptor triggers spontaneous root nodule organogenesis. Science 2007; 315:104 - 107
- Phillips DA. Abscisic acid inhibition of root nodule initiation in Pisum sativum. Planta 1971; 100:181 - 190
- Bano A, Harper JE. Plant growth regulators and phloem exudates modulate root nodulation of soybean. Funct Plant Biol 2002; 29:1299 - 1307
- Suzuki A, Akune M, Kogiso M, Imagama Y, Osuki K, Uchiumi T, et al. Control of nodule number by the phytohormone abscisic acid in the roots of two leguminous species. Plant Cell Physiol 2004; 45:914 - 922
- Liang YJ, Mitchell DM, Harris JM. Abscisic acid rescues the root meristem defects of the Medicago truncatula latd mutant. Dev Biol 2007; 304:297 - 307
- Gampala SS, Hagenbeek D, Rock CD. Functional interactions of lanthanum and phospholipase D with the abscisic acid signaling effectors VP1 and ABI1-1 in rice protoplasts. J Biol Chem 2001; 276:9855 - 9860
- Wu Y, Sanchez JP, Lopez-Molina L, Himmelbach A, Grill E, Chua NH. The abi1-1 mutation blocks ABA signaling downstream of cADPR action. Plant J 2003; 34:307 - 315
- Ding Y, Kalo P, Yendrek C, Sun J, Liang Y, Marsh JF, et al. Abscisic acid coordinates Nod Factor and cytokinin signaling during the regulation of nodulation in Medicago truncatula. Plant Cell 2008; 20:2681 - 2695
- Suganuma N, Nakamura Y, Yamamoto M, Ohta T, Koiwa H, Akao S, Kawaguchi M. The Lotus japonicus Sen1 gene controls rhizobial differentiation into nitrogen-fixing bacteroids in nodules. Mol Genet Genomics 2003; 269:312 - 320
- Krusell L, Krause K, Ott T, Desbrosses G, Kramer U, Sato S, et al. The sulfate transporter SST1 is crucial for symbiotic nitrogen fixation in Lotus japonicus root nodules. Plant Cell 2005; 17:1625 - 1636
- Kumagai H, Hakoyama T, Umehara Y, Sato S, Kaneko T, Tabata S, Kouchi H. A novel ankyrinrepeat membrane protein, IGN1, is required for persistence of nitrogen-fixing symbiosis in root nodules of Lotus japonicus. Plant Physiol 2007; 143:1293 - 1305
- Miwa H, Sun J, Oldroyd GE, Downie JA. Analysis of Nod-factor-induced calcium signaling in root hairs of symbiotically defective mutants of Lotus japonicus. Mol Plant Microbe Interact 2006; 19:914 - 923
- Han S-Y, Kitahata N, Sekimata K, Saito T, Kobayashi M, Nakashima K, et al. A novel inhibitor of 9-cis-epoxycarotenoid dioxygenase in abscisic acid biosynthesis in higher plants. Plant Physiol 2004; 135:1574 - 1582
- Trinchant J-C, Rigaud J. Nitrite and nitric oxide as inhibitors of nitrogenase from soybean bacteroids. Appl Environ Microbiol 1982; 44:1385 - 1388
- García-Mata C, Lamattina L. Nitric oxide and abscisic acid cross talk in guard cells. Plant Physiol 2002; 128:790 - 792
- Neill SJ, Desikan R, Clarke A, Hancock JT. Nitric oxide is a novel component of abscisic acid signaling in stomatal guard cells. Plant Physiol 2002; 128:13 - 16
- Schauser L, Roussis A, Stiller J, Stougaard J. A plant regulator controlling development of symbiotic root nodules. Nature 1999; 402:191 - 195
- Imaizumi-Anraku H, Takeda N, Charpentier M, Perry J, Miwa H, Umehara Y, et al. Plastid proteins crucial for symbiotic fungal and bacterial entry into plant roots. Nature 2005; 433:527 - 531
- Penmetsa RV, Cook DR. A legume ethylene-insensitive mutant hyperinfected by its rhizobial symbiont. Science 1997; 275:527 - 530
- Nishimura R, Ohmori M, Kawaguchi M. The novel symbiotic phenotype of enhanced-nodulating mutant of Lotus japonicus: astray mutant is an early nodulating mutant with wider nodulation zone. Plant Cell Physiol 2002; 43:853 - 859
- Oka-Kira E, Tateno K, Miura K, Haga T, Hayashi M, Harada K, et al. klavier (klv), A novel hypernodulation mutant of Lotus japonicus affected in vascular tissue organization and floral induction. Plant J 2005; 44:505 - 515
- Magori S, Oka-Kira E, Shibata S, Umehara Y, Kouchi H, Hase Y, et al. TOO MUCH LOVE, a root regulator associated with the long-distance control of nodulation in Lotus japonicus. Mol Plant Microbe Interact 2009; 22:259 - 268
- Tirichine L, Imaizumi-Anraku H, Yoshida S, Murakami Y, Madsen LH, Miwa H, et al. Deregulation of a Ca2+/calmodulin-dependent kinase leads to spontaneous nodule development. Nature 2006; 441:1153 - 1156
- Ishikawa K, Yokota K, Li YY, Wang Y, Liu C-T, Suzuki S, et al. Isolation of a novel root-determined hypernodulation mutant rdh1 of Lotus japonicus. Soil Sci Plant Nutr 2008; 54:259 - 263