Abstract
Coronatine (COR), a jasmonate mimic produced by Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) is required for full virulence of Pst DC3000 in tomato and Arabidopsis. COR is shown to induce a range of physiological processes including chlorosis, root growth inhibition and anthocyanin accumulation in tomato. To elucidate the host/signaling genes involved in COR-responses, we utilized a forward genetics approach using Nicotiana benthamiana and virus-induced gene silencing (VIGS) and identified genes that play a role in COR-mediated chlorosis. We designated these genes as altered COR response (ALC). When silenced, one gene designated ALC1 produced a hypersensitive/necrosis-like phenotype after COR application in a coronatine insensitive 1 (COI1)-dependent manner. In pathogenicity assays performed on Arabidopsis thylakoid formation 1 (thf1) knockout lines and SlALC1-silenced tomato plants, Pst DC3000 induced coalescing necrotic lesions in an accelerated manner. Furthermore, we showed that COR affects ALC1 localization in chloroplast in a COI1-dependent manner. In conclusion, our results show the potential of VIGS-based, forward genetic screens to identify new players in COR-mediated signal transduction.
Coronatine (COR), a phytotoxin produced by P. syringae pv. tomato (Pst DC3000) contributes to the virulence of Pst DC3000 in Arabidopsis, tomato, collard and turnip.Citation1–Citation3 It has been shown that COR has structural and functional resemblance to 12-oxo-phytodienoic acid (12-OPDA), methyl jasmonate (MeJA), and derivatives related to jasmonic acid (JA).Citation4
During a compatible interaction, COR-producing Pst DC3000 activates JA signaling, which leads to a suppression of the salicylic acid (SA) pathway.Citation1,Citation5–Citation8 One hallmark of bacterial speck disease on tomato leaves is the formation of necrotic lesions surrounded by chlorosis. COR is shown to be required for chlorosis development.Citation1–Citation3,Citation9 In addition to chlorosis, COR induces a wide array of effects in plants including anthocyanin production, alkaloid accumulation, ethylene emission, tendril coiling, inhibition of root elongation, hypertrophy and stomatal opening.Citation4,Citation9–Citation13 Despite our present understanding of COR function, it is not clear how chlorosis impacts or benefits pathogen virulence. Moreover, the molecular targets for COR and the downstream signaling cascades are not well understood.
To identify plant genes that are involved in COR signaling, we used virus-induced gene silencing (VIGS)-based forward genetic screen in Nicotiana benthamina. A N. benthamiana cDNA library cloned in tobacco rattle virus (TRV) RNA2 based VIGS vector, was used.Citation14 N. benthamiana plants were individually inoculated with Agrobacterium tumefaciens TRV2 cDNA clones,Citation14 along with an Agrobacterium strain containing pTRV1, to silence the corresponding genes in N. benthamiana.Citation14 Two weeks after inoculation with TRV, COR (0.2 nmol) was spotted on the leaves of silenced plants, and the phenotypes were recorded 5–7 days later.
After screening ∼4,000 cDNA clones, we identified five non-redundant cDNA clones that when silenced resulted in an altered COR response (ALC) phenotype upon exogenous application of COR.Citation15 The silencing of a gene designated ALC1 resulted in a hypersensitive (HR)-like necrosis instead of chlorosis after COR inoculation. ALC1 is a homologue of the Arabidopsis Thylakoid formation 1 (THF1) gene.Citation15 We also demonstrated that the loss of the N. benthamiana gene NbALC1 and its orthologs, SlALC1 in tomato and AtTHF1 in Arabidopsis, resulted in a necrotic response to COR and Pst DC3000. When ALC1-silenced tomato plants were spray-inoculated with Pst DC3000, necrotic lesions without visible chlorosis developed in an accelerated manner, and the phenotype was distinctly different from typical bacterial speck symptoms with chlorotic halos.Citation5,Citation13 Furthermore, necrosis spread beyond the region where COR was applied beginning at 10 days post-inoculation (dpi), which is similar to the runaway cell death phenotype reported earlier in the Arabidopsis lsd1 mutant.Citation15,Citation16
To determine the role of ALC1 in response to COR and Pst DC3000, we used Arabidopsis since it is genetically tractable and a host of Pst DC3000. The ortholog of ALC1 in Arabidopsis, known as THF1, is a single-copy gene with no closely related sequences in the Arabidopsis genome.Citation17 Similar to ALC1-silenced tomato plants, inoculation of Arabidopsis thf1 mutants with Pst DC3000 did not result in a typical bacterial speck symptoms. Interestingly, no differences in the Pst DC3000 populations were seen between ALC1-silenced tomato plants and the thf1 mutant line when compared to wild-type plants. These results suggested that Pst DC3000 may tightly regulate chloroplast homeostasis during infection, and the levels of THF1 for controlled necrosis during infection may assist in pathogen dissemination and spread. We recently demonstrated that COR-induced effects on the photosynthetic machinery result in the generation of light-dependent, reactive oxygen species (ROS) in tomato seedlings.Citation18,Citation19 We speculate that in COR-treated or Pst DC3000-inoculated ALC1 silenced tomatoes and in the Pst DC3000-inoculated Arabidopsis thf1 mutant, the necrotic/HR-like cell death phenotype may appear because the effect of ROS supersedes the detoxifying capacity of antioxidants.
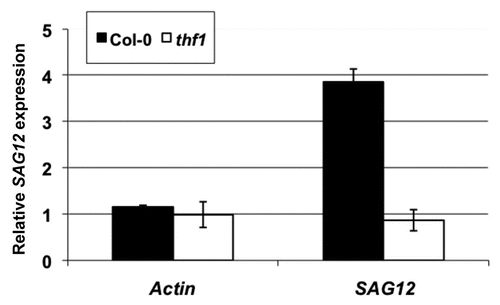
The COR-induced necrotic phenotype in ALC1-silenced plants is COI1-dependent, and the JA-mediated defense pathway is functional but subdued in Arabidopsis thf1 mutants.Citation15 Since COR is a functional mimic of JA and is shown to induce senescence in Arabidopsis,Citation20 we investigated whether the senescence-associated gene SAG12 was differentially expressed in the Arabidopsis thf1 mutant in response to Pst DC3000. Using qRT-PCR, we analyzed the transcript level of SAG12 four days after infiltration with Pst DC3000. Infected leaves of Col-0 expressed approximately four-fold higher levels of SAG12 () as compared to the thf1 mutant line. These results suggested that the typical Pst DC3000-induced chlorosis is associated with senescence.
Furthermore, results from GFP-tagged ALC1 suggest that COR has direct effects on ALC1 and might target ALC1 to degradation in a COI1-dependent manner.Citation15 Based on these results it is tempting to speculate that ALC1/THF1 may directly interact with COR if localized in the chloroplast membrane. Interestingly, a chloroplast protein in wheat, ToxABP1 (an ortholog of THF1), directly interacts with the Pyrenophora tritici-repentis protein ToxA.Citation21
In conclusion, we have developed a VIGS-based forward genetic screen for identification of new targets involved in COR signaling. Although the precise role of THF1 in COR signaling pathway could be argued and needs further confirmation, our results present a new role for chloroplast-localized THF1 in bacterial speck disease development.
Figures and Tables
Figure 1 Senescence-associated SAG12 expression in thf1 mutants in response to Pst DC3000. Transcripts of SAG12 were quantified by real time quantitative PCR in wild-type (Col-0, black bars) and thf1 mutant (open bars) lines of Arabidopsis after Pst DC3000 inoculation. Four-week old plants of Col-0 and the thf1 mutant were syringe-infiltrated with either Pst DC3000 (106 CFU/ml) or buffer (mock control). The transcript levels were quantified relative to the transcript levels on mock control, which was assigned a value of 1.
Acknowledgements
This work was supported by the Samuel Roberts Noble Foundation and in part by a grant to S.R. Uppalapati from Oklahoma Center for Advancement of Science and Technology (PSB09-021). C.L. Bender acknowledges financial support from National Science Foundation grant (IOB-0620469). A spinning disk confocal microscope was purchased by an equipment grant from NSF (DBI-0722635).
Addendum to:
References
- Brooks DM, Hernandez-Guzman G, Kloek AP, Alarcon-Chaidez F, Sreedharan A, Rangaswamy V, et al. Identification and characterization of a well-defined series of coronatine biosynthetic mutants of Pseudomonas syringae pv. tomato DC3000. Mol Plant-Microbe Interact 2004; 17:162 - 174
- Elizabeth SV, Bender CL. The phytotoxin coronatine from Pseudomonas syringae pv. tomato DC3000 functions as a virulence factor and influences defence pathways in edible brassicas. Mol Plant Pathol 2007; 8:83 - 92
- Uppalapati SR, Ishiga Y, Wangdi T, Kunkel BN, Anand A, Mysore KS, Bender CL. The phytotoxin coronatine contributes to pathogen fitness and is required for suppression of salicylic acid accumulation in tomato inoculated with Pseudomonas syringae pv. tomato DC3000. Mol Plant-Microbe Interact 2007; 20:955 - 965
- Weiler EW, Kutchan TM, Gorba T, Brodschelm W, Niesel U, Bublitz F. The Pseudomonas phytotoxin coronatine mimics octadecanoid signalling molecules of higher plants. FEBS Lett 1994; 345:9 - 13
- Zhao Y, Thilmony R, Bender CL, Schaller A, He SY, Howe GA. Virulence systems of Pseudomonas syringae pv. tomato promote bacterial speck disease in tomato by targeting the jasmonate signaling pathway. Plant J 2003; 36:485 - 499
- Laurie-Berry N, Joardar V, Street IH, Kunkel BN. The Arabidopsis thaliana JASMONATE INSENSITIVE 1 gene is required for suppression of salicylic acid-dependent defenses during infection by Pseudomonas syringae. Mol Plant-Microbe Interact 2006; 19:789 - 800
- Kloek AP, Verbsky ML, Sharma SB, Schoelz JE, Vogel J, Klessig DF, Kunkel BN. Resistance to Pseudomonas syringae conferred by an Arabidopsis thaliana coronatine-insensitive (coi1) mutation occurs through two distinct mechanisms. Plant J 2001; 26:509 - 522
- Kunkel BN, Brooks DM. Cross talk between signaling pathways in pathogen defense. Curr Opin Plant Biol 2002; 5:325 - 331
- Bender CL, Stone HE, Sims J, Cooksey DA. Reduced pathogen fitness of Pseudomonas syringae pv. tomato Tn5 mutants defective in coronatine production. Physiol Mol Plant Pathol 1987; 30:272 - 283
- Feys B, Benedetti CE, Penfold CN, Turner JG. Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistant to a bacterial pathogen. Plant Cell 1994; 6:751 - 759
- Lauchli R, Boland W. Indanoyl amino acid conjugates: tunable elicitors of plant secondary metabolism. Chem Rec 2003; 3:12 - 21
- Melotto M, Underwood W, Koczan J, Nomura K, He SY. Plant stomata function in innate immunity against bacterial invasion. Cell 2006; 126:969 - 980
- Uppalapati SR, Ayoubi P, Weng H, Palmer DA, Mitchell RE, Jones W, Bender CL. The phytotoxin coronatine and methyl jasmonate impact multiple phytohormone pathways in tomato. Plant J 2005; 42:201 - 217
- Anand A, Vaghchhipawala Z, Ryu CM, Kang L, Wang K, del-Pozo O, et al. Identification and characterization of plant genes involved in Agrobacterium-mediated plant transformation by virus-induced gene silencing. Mol Plant-Microbe Interact 2007; 20:41 - 52
- Wangdi T, Uppalapati SR, Nagaraj S, Ryu CM, Bender CL, Mysore KS. A virus-induced gene silencing screen identifies a role for Thylakoid Formation1 in Pseudomonas syringae pv tomato symptom development in tomato and Arabidopsis. Plant Physiol 2010; 152:281 - 292
- Jabs T, Dietrich RA, Dangl JL. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science 1996; 273:1853 - 1856
- Wang Q, Sullivan RW, Kight A, Henry RL, Huang J, Jones AM, Korth KL. Deletion of the chloroplast-localized Thylakoid formation1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves. Plant Physiol 2004; 136:3594 - 3604
- Ishiga Y, Uppalapati SR, Ishiga T, Elavarthi S, Martin B, Bender CL. The phytotoxin coronatine induces light-dependent reactive oxygen species in tomato seedlings. New Phytol 2009; 181:147 - 160
- Ishiga Y, Uppalapati SR, Ishiga T, Elavarthi S, Martin B, Bender CL. Involvement of coronatine-inducible reactive oxygen species in bacterial speck disease of tomato. Plant Signal Behav 2009; 4:237 - 239
- He Y, Fukushige H, Hildebrand DF, Gan S. Evidence supporting a role of jasmonic acid in Arabidopsis leaf senescence. Plant Physiol 2002; 128:876 - 884
- Manning VA, Hardison LK, Ciuffetti LM. Ptr ToxA interacts with a chloroplast-localized protein. Mol Plant-Microbe Interact 2007; 20:168 - 177