Abstract
The ability to distinguish self from non-self allows organisms to protect themselves against attackers. Sagebrush plants use volatile cues emitted by clipped neighbors to adjust their defenses against herbivores. Recently, we reported that cues from genetically identical ‘self’ clones were more effective at reducing damage than were cues from ‘non-self’ clones. This indicates that plants can distinguish self from non-self through volatiles and respond differentially. Identity recognition may be an essential step in enabling plants to behave cooperatively. Emission of cues which enable other plant tissues (on the same or other individual) to respond appropriately to herbivore risk may have evolved if cues are aimed primarily at self tissue.
The ability to recognize self from nonself is a fundamental property of individuals of all multicellular organisms. Distinguishing between molecules that are part of one's own tissues and those of an invader provides a first step towards the evolution of a functioning immune system. An immune system responds differently towards self and nonself tissues, destroying the later. In addition to immune responses, many other sophisticated behaviors have been described for animals that differentiate self from non-self and even kin from strangers.Citation1 For example, social behaviors including altruism can be favored by natural selection when animals are able to first distinguish kin from non-kin and respond differently to individuals in these two categories.Citation2 Although plant behavior is far less well studied, plants too display many sophisticated and context-dependent behaviors.Citation3
Plant biologists have described various situations in which plants exhibit different behaviors based on identity. It has been known for some years that many angiosperms choose mates based on genetic identity.Citation4 Numerous mechanisms have been described, primarily involving differential germination of pollen, growth of pollen tubes through stigmatic tissue, and production of competent zygotes. More recently, several workers have found that plants may differentiate self from non-self and alter their morphologies in response to cues from these two types of sources. Plants appeared to recognize their own roots and to grow fewer and shorter roots when they contacted self roots compared to non-self roots (reviewed in references Citation5–Citation7). A common feature of these experimental studies is that roots only showed self-recognition when they were physically attached. These experimental studies may be subject to alternate explanations.Citation8,Citation9
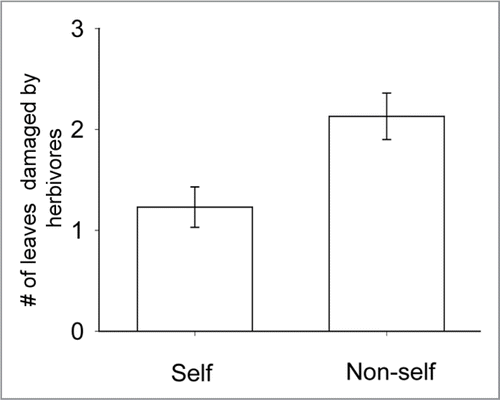
Recently we reported that sagebrush plants induced resistance more effectively against their herbivores in response to the volatile cues emitted by self clones compared to the cues of non-self clones.Citation10 We had previously found that experimental clipping to branches caused systemic induced resistance within an individual against herbivores only when volatile cues were transmitted.Citation11 To evaluate self/non-self discrimination we first produced clones of 60 parent plants in the field by root crown division. These potted clones were propagated and then placed back in the field near either their genetically identical parent (self treatment) or a genetically different parent (nonself treatment). The potted clones were experimentally clipped in spring for both treatments and the damage that accumulated over the growing season was recorded for parents near self and non-self clones. We found that plants near clipped self clones received approximately 42% less damage by their herbivores than plants near clipped non-self clones (, One-way ANOVA, F1,58 = 8.72, p = 0.005).
This result is novel in several ways. Past results showing self/nonself recognition between roots required that they be in physical contact for discrimination to occur; physical contact was not required in this case. In addition, this is the first identity study to measure responses in terms of damage by herbivores rather than plant morphology or reproduction. This result is more robust than the changes in root morphology because changes attributed to self or non-self volatiles cannot be explained by alternative hypotheses involving potentially confounding differences in resource availability or pot size.Citation8,Citation9 The ability of plants to differentiate self from non-self is important because it may enable differential treatment towards ramets that share genes.
Recent work has also suggested that plants may be able to discriminate between kin and strangers. Cakile edentula and Impatiens pallida changed their morphologies depending upon whether their roots contacted kin or strangers.Citation12,Citation13 These altered morphologies were consistent with the notion that kin cooperated and non-kin competed. Examination of self/non-self recognition and kin/stranger recognition patterns in Arabidopsis thaliana indicated that these two forms of identity discrimination were affected differently by inhibitors and therefore suggested that they may involve different signaling mechanisms.Citation14
Plants that emit volatile cues that other individuals can use to adjust their defenses (eavesdropping) may be at a selective disadvantage.Citation15 Why should a plant dispense information that allows its neighbors to fine tune their defenses against herbivory? One possible answer to this conundrum may be that plants emit volatile cues to coordinate their own defenses since volatile cues are active over relatively short distances. A second possible answer is that greater sensitivity to self volatiles reduces the cost of eaves-dropping. In designing our sagebrush experiment we cloned plants as a means of producing physically separate pairs of plants that were either genetically identical or different. Early genetic work indicated that populations of sagebrush were highly structured genetically.Citation16 In other words, relatedness decreased as a function of the distance between individuals, also known as population viscosity. Recent genetic analyses of microsatellites indicate that vegetative reproduction by rhizomes also occurs in this species and some neighbors in nature are genetically identical (Ishizaki, et al. in review). Population viscosity has been considered to increase the likelihood of cooperation, in part because neighbors already share genes.Citation2,Citation17 Applying similar logic, communication is facilitated by kin recognition if relatives are better able to communicate than non-kin. Communication may be favored if the tissue emitting cue is surrounded by primarily self tissue or if the exchange of cues is more effective and likely to occur between self tissues. In conclusion, plant communication using volatile cues may have evolved because individual plants were communicating primarily with themselves.
Figures and Tables
Figure 1 The mean number of leaves that were damaged by herbivores (grasshoppers, caterpillars and deer) on assay branches of sagebrush (±1 se). Cuttings were either genetically identical (self) or different (non-self) from the assay branch; assay branches were within 5 cm of potted cuttings but not in physical contact. Cuttings were experimentally clipped to simulate herbivory in May and herbivore damage accumulated on the assay branches until season's end in September when damage was assayed.
Acknowledgements
We thank Susan Dudley for comments that improved this paper and the JSPS for financial support.
Addendum to:
References
- Wilson EO. Sociobiology 1975; Cambridge Massachusetts Belknap Press
- Hamilton WD. The genetical evolution of social behavior. J Theor Biol 1964; 7:1 - 52
- Karban R. Plant behaviour and communication. Ecol Lett 2008; 11:727 - 739
- Charlesworth D, Vekemans X, Castric V, Glemin S. Plant self-incompatibility systems: a molecular evolutionary perspective. New Phytol 2005; 168:61 - 69
- Mahall BE, Callaway RM. Root communication among desert shrubs. Proc Natl Acad Sci USA 1991; 88:874 - 876
- Holzapfel C, Alpert P. Root cooperation in a clonal plant; connected strawberries segregate roots. Oecologia 2003; 134:72 - 77
- Gruntman M, Novoplansky A. Physiologically mediated self/non-self discrimination in roots. Proc Natl Acad Sci USA 2004; 101:3863 - 3867
- Schenk HJ. Root competition: beyond resource depletion. J Ecol 2006; 94:725 - 739
- Hess L, De Kroon H. Effects of rooting volume and nutrient availability as an alternative explanation for root self/non-self discrimination. J Ecol 2007; 95:241 - 251
- Karban R, Shiojiri K. Self-recognition affects plant communication and defense. Ecol Lett 2009; 12:502 - 506
- Karban R, Shiojiri K, Huntzinger M, McCall A. Damage-induced resistance in sagebrush: volatiles are key to intra- and interplant communication. Ecology 2006; 87:922 - 930
- Dudley SA, File AL. Kin recognition in an annual plant. Biol Lett 2007; 3:435 - 438
- Murphy GP, Dudley SA. Kin recognition: competition and cooperation in Impatiens (Balsaminaceae). Am J Bot 2009; 96:1990 - 1996
- Biedrzycki ML, Jilany TA, Dudley SA, Bais HP. Root exudates mediate kin recognition in plants. Commun Integr Biol 2010; 3:1 - 8
- Heil M, Karban R. Explaining evolution of plant communication by airborne signals. Trends Ecol Evol 2010; 25:137 - 144
- Cook CW, Stoddart LA. Physiological responses of big sagebrush to different types of herbage removal. J Range Manag 1960; 13:14 - 16
- Platt TG, Bever JD. Kin competition and the evolution of cooperation. Trends Ecol Evol 2009; 24:370 - 377