Abstract
Using two lowland rice (Oryza sativa L.) cultivars we found that in both cases submerged-induced elongation early after germination depends on gibberellins (GAs). Submergence increases the content of the active GA1 by enhancing the expression of GA biosynthesis genes, thus facilitating the seedlings to escape from the water and preventing asphyxiation. However, the two cultivars differ in their response to ethylene. The cultivar Senia (short), by contrast to cultivar Bomba (tall), does not elongate after ethylene application, and submerged-induced elongation is not negated by an inhibitor of ethylene perception. Also, while ethylene emanation in Senia is not altered by submergence, Bomba seedlings emanate more ethylene upon de-submergence, associated with enhanced expression of the ethylene biosynthesis gene OsACS5. The cultivar Senia thus allows the possibility of clarifying the role of ethylene and other factors as triggers of GA biosynthesis enhancement in rice seedlings under submergence.
To Grow or Not to Grow, That is the Question
Many plants (e.g., from Rumex and Echinochloa genera) are able to survive in flooded soils as a result of faster growth induced by gibberellins (GA), thus keeping their leaves in the air and preventing asphyxiation.Citation1,Citation2 This is also the strategy used by deepwater rice, grown in heavy rain areas from southeastern Asia.Citation3,Citation4 By contrast, lowland rice cultured in regions subjected to occasional and temporal flooding, that do not grow fast enough to escape from flooding, may benefit from shoot elongation restriction under these conditions because this allows saving energy reserves until resuming development following eventual de-submergence.Citation5–Citation7 This kind of rice tolerance to submergence is conferred by an ethylene responsive factor (encoded by the gene Sub1A), that reduces GA-inducible expression under submergence by increasing the levels of the GA signaling repressors SLR1 and SLRL1.Citation8,Citation9
Lowland rice is germinated in the field under a layer of water in many countries (e.g., in most Mediterranean regions), but at later developmental stages only the basal part of the plant is maintained under water. In this case, increased elongation capacity of the seedlings is a clear advantage to help reaching contact with air and start active photosynthesis as soon as possible. This is opposite to the situation described above, where reduced growth of adult plants under water facilitates eventual growth resumption after temporary flooding receding.
Submergence Induces Elongation of Lowland Rice Seedlings by Increasing their Content in Active GA1 through Alteration of GA Metabolism
A comparison study of two lowland rice cultivars grown in the Comunidad Valenciana, Spain (Senia, low and Bomba, tall), showed that early growth after germination, and also that the enhanced-induced elongation under water depend on GA, in agreement with results obtained using other cultivars.Citation4 This conclusion was based on the observation that rice elongation was reduced by paclobutrazol (an inhibitor of GA biosynthesis) and that this effect was counteracted by application of GA3. More important, enhanced elongation was associated with an increase on GA1 content, the purported active GA in rice.Citation10 The contents of GA53 and GA19, but not of GA20, immediate GA1 precursors,Citation10,Citation11 also increased. The submergence increases GA1 content in the seedlings as a result of its effect on GA metabolism, mainly by enhancing the expression of OsGA20ox1 and −2, and of OsGA3ox2 (GA biosynthesis genes). It is no clear whether the decrease of GA inactivating genes (OsGA2ox) has also a role in the process.
Submergence-induced Elongation is not Mediated by Ethylene in the Lowland Rice Cultivar Senia
It is known that in the case of deepwater rice stem elongation under submergence is due to the increase of ethylene content in the tissues (due to less diffusion under water but also to enhanced new synthesis),Citation2–Citation4,Citation12 that stimulates GA biosynthesis. A similar conclusion was obtained in the case of the tall lowland rice cultivar IR36.Citation4 We therefore investigated whether ethylene was also active in enhancing lowland rice seedling elongation of the Bomba and Senia cultivars. Interestingly, while Bomba seedling elongated in response to ethylene or 1-aminocyclopropane-1-carboxylic-acid (ACC; an ethylene precursor) application, as expected, Senia seedlings did not. Moreover, submerged-induced elongation in Bomba was negated in the presence of 1-methylcyclopropene (1-MCP; an ethylene perception inhibitor), but not in Senia. These results suggested that ethylene is not a factor triggering GA-induced elongation upon submergence in Senia. This hypothesis was also supported by the observation that while ethylene emanation increased about twice in Bomba seedlings following de-submergence it was not affected in Senia (where the rate of ethylene emanation was similar to that found in Bomba submerged seedlings). The increase of ethylene emanation in Bomba was associated with rapid enhanced expression of the ethylene biosynthetic gene OsACS5 upon submergence, an effect not found in cultivar Senia. The expression of other genes involved in ethylene biosynthesis (OsACS1, −2 and −3 and OsACO1) did not change. In agreement with previous observations in deep-water rice,Citation13,Citation14 our results suggest that in Bomba OsACC5 plays an important role in early submergence-induced ethylene biosynthesis.
Conclusions
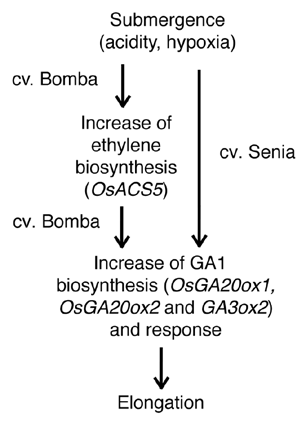
As summarized in , submerged-induced seedling elongation of lowland rice is a consequence of increased GA1 content due to enhanced expression of several GA biosynthesis genes (OsGA20ox1, OsGA20ox2 and OsGA3ox2). This effect, as expected, seems to be mediated by ethylene in the tall cultivar Bomba, where submergence also enhances expression of the ethylene biosynthesis gene OsACS5 and higher ethylene emanation. By contrast, in the short cultivar Senia submergence-induced elongation does not seem to be mediated by ethylene. In this case, the signal triggering GA biosynthesis may be the acidity produced by CO2 accumulation, as occurs in Potamogeton pectinatus.Citation15,Citation16 Another factor that might also act as a triggering signal for submergence-induced elongation is hypoxia, because sheath elongation of both Senia and Bomba seedlings was enhanced when grown in a 3% O2 atmosphere.
Transgenic rice plants expressing antisense OsEin2, a positive regulator of the signal ethylene pathway, have reduced stature and display higher rate of ethylene biosynthesis.Citation17 In Arabidopsis thaliana, blocking the ethylene-signaling pathway also enhances ethylene biosynthesis rate.Citation18 Therefore, the insensitivity of Senia to ethylene and 1-MCP, and its higher ethylene biosynthesis compared to Bomba, suggests that ethylene reception and/or signal transduction pathway may be impaired in that cultivar, thus affecting ethylene metabolism.
Figures and Tables
Figure 1 Scheme summarizing the different mechanisms explaining the submergence-induced elongation in two lowland rice cultivars, Bomba (tall) and Senia (short). In both cases, elongation is the result of an increase of active GA (GA1) biosynthesis due to enhanced expression of GA biosynthesis genes (OsGA20ox1, OsGA20ox2 and OsGA3ox2). In the case of Bomba this is induced, at least partially, by an increase of ethylene biosynthesis due to upregulation of OsACS5 expression. In the case of Senia, submergence-induced elongation does not depend on ethylene, and the GA-mediated response is triggered by an still unknown mechanism, probably involving increase of acidity and/or hypoxia.
Addendum to:
References
- Bailey-Serres J, Voesenek LACJ. Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol 2008; 59:313 - 339
- Jackson MB. Ethylene-promoted elongation: an adaptation to submergence stress. Ann Bot 2008; 101:229 - 248
- Kende H, van der Knaap E, Cho HT. Deepwater rice: a model plant to study stem elongation. Plant Physiol 1998; 118:1105 - 1110
- Van Der Straeten D, Zhou Z, Prinsen E, Van Onckelen HA, Van Montagu MC. A comparative molecular-physiological study of submergence response in lowland and deepwater rice. Plant Physiol 2001; 125:955 - 968
- Ismail AM, Ella ES, Vergara GV, Mackill DH. Mechanisms associated with tolerance in flooding during germination and early seedling growth in rice (Oryza sativa). Ann Bot 2009; 103:197 - 209
- Kawano N, Ito O, Sakagami JI. Morphological and physiological responses of rice seedlings to complete submergence (flash flooding). Ann Bot 2009; 103:161 - 169
- Bailey-Serres J, Voesenek LACJ. Life in the balance: a signalling network controlling survival of flooding. Curr Opin Plant Biol 2010; 13:1 - 6
- Perata P, Voesenek LACJ. Submergence tolerance in rice requires Sub1A, an ethylene-response-factor-like gene. Trends Plant Sci 2007; 12:43 - 46
- Fukao T, Bailey-Serres J. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc Natl Acad Sci USA 2008; 105:16814 - 16819
- Kobayashi M, MacMillan J, Phinney B, Gaskin P, Spray CR, et al. Gibberellin biosynthesis: metabolic evidence for three steps in the early-13-hydroxylation pathway of rice. Phytochemistry 2000; 55:317 - 321
- Yamaguchi S. Gibberellin metabolism and its regulation. Annu Rev Plant Biol 2008; 59:225 - 251
- Vriezen WH, Zhou Z, Van Der Straeten D. Regulation of submergence-induced enhanced shoot elongation in Oryza sativa L. Ann Bot 2003; 91:263 - 270
- Van Der Straeten D, Anuntalabhochai S, Van Caeneghem W, Zhou Z, Gielen J, Van Montagu M. Expression of three members of the ACC synthase gene family in deepwater rice by submergence, wounding and hormonal treatments. Plant Sci 1997; 124:79 - 87
- Zhou Z, Engler JA, Rouan D, Michiels F, Van Montagu M, et al. Tissue localization of a submergence-induced 1-aminocyclopropane-1-carboxylic acid synthase in rice. Plant Physiol 2002; 129:72 - 84
- Summers JE, Jackson MB. Anaerobic promotion of stem extension in Potamogeton pectinatus. Roles for carbon dioxide, acidification and hormones. Physiol Plant 1996; 96:615 - 622
- Voesenek LACJ, Blom CWPM. Lerner HP. Stimulated shoot elongation: a mechanism of semiaquatic plants to avoid submergence stress. Plant Responses to Environmental Stresses: From Phytohormones to Genome Reorganization 1999; New York Marcel Dekker Inc 431 - 448
- Jun SH, Han MJ, Lee S, Seo YS, Kim WT, An G. OsEIN2 is a positive component in ethylene signaling in rice. Plant Cell Physiol 2004; 45:281 - 289
- Guzmán P, Ecker JR. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell 1990; 2:513 - 523