Abstract
The accumulation of silicon (Si) differs greatly with plant species and cultivars due to different ability of the roots to take up Si. In Si accumulating plants such as rice, barley and maize, Si uptake is mediated by the influx (Lsi1) and efflux (Lsi2) transporters. Here we report isolation and functional analysis of two Si efflux transporters (CmLsi2-1 and CmLsi2-2) from two pumpkin (Cucurbita moschata Duch.) cultivars contrasting in Si uptake. These cultivars are used for rootstocks of bloom and bloomless cucumber, respectively. Different from mutations in the Si influx transporter CmLsi1, there was no difference in the sequence of either CmLsi2 between two cultivars. Both CmLsi2-1 and CmLsi2-2 showed an efflux transport activity for Si and they were expressed in both the roots and shoots. These results confirm our previous finding that mutation in CmLsi1, but not in CmLsi2-1 and CmLsi2-2 are responsible for bloomless phenotype resulting from low Si uptake.
Silicon (Si) is the second most abundant elements in earth's crust.Citation1 Therefore, all plants rooting in soils contain Si in their tissues. However Si accumulation in the shoot differs greatly among plant species, ranging for 0.1 to 10% of dry weight.Citation1–Citation3 In higher plants, only Poaceae, Equisetaceae and Cyperaceae show a high Si accumulation.Citation2,Citation3 Si accumulation also differs with cultivars within a species.Citation4,Citation5 These differences in Si accumulation have been attributed to the ability of the roots to take up Si.Citation6,Citation7
Genotypic difference in Si accumulation has been used to produce bloomless cucumber (Cucumis sativus L.).Citation8 Bloom (white and fine powders) on the surface of cucumber fruits is primarily composed of silica (SiO2).Citation9 However, nowadays, cucumber without bloom (bloomless cucumber) is more popular in Japan due to its more attractive and distinctly shiny appearance. Bloomless cucumber is produced by grafting cucumber on some specific pumpkin (Cucurbita moschata Duch.) cultivars. These pumpkin cultivars used for bloomless cucumber rootstocks have lower silicon accumulation compared with the rootstocks used for producing bloom cucumber.Citation9
Our study showed that the difference in Si accumulation between bloom and bloomless root stocks of pumpkin cultivars results from different Si uptake by the roots.Citation10 Si uptake has been demonstrated to be mediated by two different types of transporters (Lsi1 and Lsi2) in rice, barley and maize.Citation11–Citation15 Lsi1 is an influx transporter of Si, belonging to a NIP subfamily of aquaporin family.Citation10,Citation11,Citation13,Citation14 This transporter is responsible for transport of Si from external solution to the root cells.Citation11 On the other hand, Lsi2 is an efflux transporter of Si, belonging to putative anion transporter.Citation12 Lsi2 releases Si from the root cells towards the xylem. Both Lsi1 and Lsi2 are required for Si uptake by the roots.Citation11,Citation12 To understand the mechanism underlying genotypic difference in Si uptake, we have isolated and functionally characterized an influx Si transporter CmLsi1 from two pumpkin cultivars used for rootstocks of bloomless and bloom cucumber.Citation10 Sequence analysis showed only two amino acids difference of CmLsi1 between two pumpkin cultivars. However, CmLsi1 from bloom rootstock [CmLsi1(B+)] showed transport activity for Si, whereas that from bloomless rootstock [CmLsi1(B−)] did not.Citation10 Furthermore, we found that loss of Si transport activity was caused by one amino acid mutation at the position of 242 (from proline to leucine).Citation10 This mutation resulted in failure to be localized at the plasma membrane, which is necessary for functioning as an influx transporter. The mutated protein was localized at the ER.Citation10 Here, we report isolation and expression analysis of Si efflux transporters from two pumpkin cultivars contrasting in Si uptake and accumulation to examine whether Si efflux transporter is also involved in the bloom and bloomless phenotypes.
Isolation of Si Efflux Transporters from Pumpkin
Based on the conserved sequence between OsLsi2 and melon EST information, we designed the degenerated primers and performed PCR to obtain partial sequences of pumpkin Lsi2 from two cultivars [Shintosa (B+) and Super-unryu (B−)] used for rootstocks of bloom and bloomless cucumber, respectively.Citation16 With the help of 5′- and 3′-RACE (Rapid Amplification of cDNA End), we were able to obtain two copies of the Si efflux transporter candidate genes, CmLsi2-1 and CmLsi2-2, from either cultivar. There was no difference in the nucleotide sequences of either gene between two pumpkin cultivars (GenBank/EMBL accession numbers: AB551951 and AB551952; and in amino acid sequence).
CmLsi2-1 and CmLsi2-2 genes encode a peptide with 549 and 529 amino acids, respectively ( and ). They shared 87% identity each other at the amino acid level. CmLsi2-1 and CmLsi2-2 share 80% identity with Os10g0547500, a homolog of OsLsi2 in rice (). The identity of CmLsi2-1 and CmLsi2-2 with OsLsi2 was 67% and 63%, respectively; at amino acid level ( and ).
Efflux Activity of Si of CmLsi2-1 and CmLsi2-2
To investigate whether CmLsi2-1 and CmLsi2-2 have an efflux transport activity for silicic acid (Si) like OsLsi2, they were heterologously expressed in the Xenopus laevis oocytes. When 1 mM Si labeled with 68Ge was preloaded to the oocytes by micro-injector, a significant higher release from the oocytes expressing either CmLsi2-1 or CmLsi2-2 was found than no expression control (water injected instead of cRNA) (). This result indicates that similar to OsLsi2, both CmLsi2-1 and CmLsi2-2 have an efflux transport activity for Si.
Expression Pattern of CmLsi2-1and CmLsi2-2
Tissue-specific expression of both CmLsi2-1 and CmLsi2-2 was investigated by quantitative real-time RT-PCR in two pumpkin cultivars. Both CmLsi2-1 and CmLsi2-2 were expressed in the roots and shoots of either cultivar ( and B). This expression pattern is different from OsLsi2, ZmLsi2 and HvLsi2, which are mainly expressed in the roots at young stage.Citation12,Citation15 The expression level of either CmLsi2-1 or CmLsi2-2 in the shoots was similar between two cultivars, but that in the roots was higher in the bloom rootstock cultivar than bloomless rootstock cultivar ( and B). Although the efflux Si transporter Lsi2 is also important for Si uptake as demonstrated in rice,Citation12 the lower expression level of both CmLsi2-1 and CmLsi2-2 in the bloomless rootstock cultivar is not likely responsible for bloomless phenotype because the influx Si transporter CmLsi1 in this cultivar is not able to transport Si from external solution to the root cells due to its subcellular mislocalization.Citation10
CmLsi2-1 and CmLsi2-2 are the first Si efflux transporters identified in dicotyledonous plant. In monocots such as rice, barley and maize, one Si efflux transporter from each species has been functionally characterized. OsLsi2 from rice is localized at both exodermis and endodermis with polarity,Citation12 while ZmLsi2 and HvLsi2 are localized only at the endodermis without polarity.Citation15 In rice genome, there are four homologs of OsLsi2 (),Citation12 which has not been functionally characterized. CmLsi2-1 and CmLsi2-2 may be involved in Si uptake and/or cellular transport of Si. More detailed functional analysis on both CmLsi2-1 and CmLsi2-2 such as cellular localization and tissue specific expression will be required to understand the exact role of these two transporters in pumpkin.
Accession Numbers
Sequence data from this article can be found in the Arabidopsis Genome Initiative or GenBank/EMBL databases under the following accession numbers: AB551951 and AB551952 for CmLsi2-1 and CmLsi2-2, respectively.
Figures and Tables
Figure 1 Alignment of amino acid sequence of Lsi2-like Si efflux transporters in rice (OsLsi2) and pumpkin. Two copies of Lsi2-like gene encoding CmLsi2-1 and CmLsi2-2 were obtained from pumpkin cultivars; Shintosa (B+) and Super-unryu (B−).
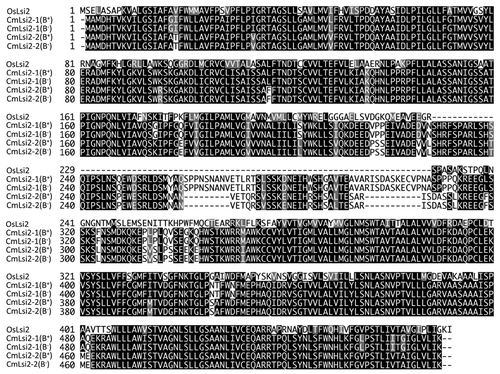
Figure 2 Phylogenetic tree of Si efflux transporter gene homologs in pumpkin (Cm−), rice (Os−), maize (Zm−), barley (Hv−) and Arabidopsis (At−). The 0.1 scale shows substitution distance.
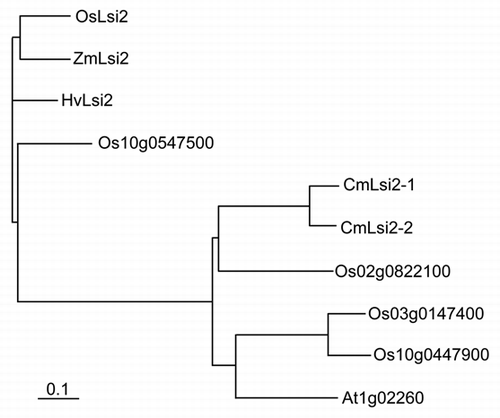
Figure 3 Efflux transport activity for Si in Xenopus oocyte. Oocytes expressing CmLsi2-1, CmLsi2-2 or no expression control (water injected) were injected with Si labeled with 68Ge. The radioactivity released from the oocytes was determined. Data are means ± SD of three biological replicates. * means significant difference at p < 0.05 (by student's t test).
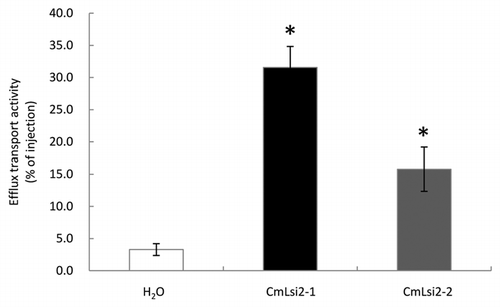
Figure 4 Tissue-specific expression of CmLsi2-1 (A) and CmLsi2-2 (B) in the roots and leaves of two pumpkin cultivars: Shintosa (B+, bloom rootstock) and Super-unryu (B−, bloomless rootstock). The expression level was determined by absolute quantitative RT-PCR. Data are means ± SD of three biological replicates. Different letters above the columns indicate statistically significant differences at p < 0.05 by Tukey's test.
Acknowledgments
This work was supported by a Grant-in-Aid for Scientific Research on Innovative Areas from the Ministry of Education, Culture, Sports, Science and Technology of Japan (22119002 to J.F.M.), a grant from the Ministry of Agriculture, Forestry and Fisheries of Japan, Genomics for Agricultural Innovation IPG-0006 (to J.F.M.) and Ohara Foundation for Agricultural Research.
Addendum to:
References
- Epstein E. Silicon. Annu Rev Plant Physiol Plant Mol Biol 1999; 50:641 - 664
- Hodson MJ, White PJ, Mead A, Broadley MR. Phylogenetic variation in the silicon composition of plants. Ann Bot 2005; 96:1027 - 1046
- Ma JF, Takahashi E. Soil, Fertilizer and Plant Silicon Research in Japan 2002; Amsterdam Elsevier Science
- Ma JF, Higashitani A, Sato K, Tateda K. Genotypic variation in Si content of barley grain. Plant Soil 2003; 249:383 - 387
- Deren CW, Datnoff LE, Snyder GN. Variable silicon content of rice cultivars grown on Everglades histosols. J Plant Nutr 1992; 15:2363 - 2368
- Takahashi E, Ma JF, Miyake Y. The possibility of silicon as an essential element for higher plants. Comments Agr Food Chem 1990; 2:99 - 122
- Mitani N, Ma JF. Uptake system of silicon in different plant species. J Exp Bot 2005; 56:1255 - 1261
- Seki M, Hotta Y. Effect of bloomless stock cultivar on the growth and mineral uptake of cucumber plants. Res Bull Aichi Agric Res Ctr 1997; 29:127 - 133
- Yamamoto Y, Hayashi M, Kanamaru T, Watanabe T, Mametsuka S, Tanaka Y. Studies on bloom on the surface of cucumber fruits, 2: Relation between the degree of bloom occurrence and contents of mineral elements. Bull Fukuoka Agr Res Ctr 1989; 9:1 - 6
- Mitani N, Yamaji N, Ago Y, Iwasaki K, Ma JF. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in Si accumulation. Plant J 2011; 66:231 - 240
- Ma JF, Tamai K, Yamaji N, Mitani N, Konishi S, Katsuhara M, et al. A silicon transporter in rice. Nature 2006; 440:688 - 691
- Ma JF, Yamaji N, Mitani N, Tamai K, Konishi S, Fujiwara T, et al. An efflux transporter of silicon in rice. Nature 2007; 448:209 - 212
- Chiba Y, Mitani N, Yamaji N, Ma JF. HvLsi1 is a silicon influx transporter in barley. Plant J 2009; 57:810 - 818
- Mitani N, Yamaji N, Ma JF. Identification of maize silicon influx transporters. Plant Cell Physiol 2009; 50:5 - 12
- Mitani N, Chiba Y, Yamaji N, Ma JF. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009; 21:2133 - 2142
- Sakata Y, Sugiyama M, Ohara T, Morishita M. Influence of rootstocks on the resistance of grafted cucumber (Cucumis sativus L.) scions to powdery mildew (Podosphaera xanthii Braun U, Shishkoff N). J Japan Soc Hort Sci 2006; 75:135 - 140