Abstract
Salicylic acid (SA) is a critical signal for activation of both local and systemic resistance responses. However, SA-deficient plants adapt to RNA virus infections better, which show a less-severe symptom and less reactive oxygen species (ROS) accumulation. The higher levels of reduced glutathione (GSH) and reduced ascorbic acid (AsA) in SA-deficient plants may contribute to their alleviated symptoms, which are consistent with their higher expression levels of dehydroascorbate reductase gene (DHAR) and glutathione reductase gene (GR). High-dose AsA or GSH treatment could alleviate the symptom and inhibit virus replication after 20 days, but ROS eliminators could not imitate the effect of AsA or GSH. The data show a new link between SA and AsA/GSH-mediated redox homeostasis.
The role of salicylic acid (SA) in defense mechanisms against pathogens has been known for several years. Many studies have demonstrated that SA plays a central role in signaling for the activation of both local resistance (LR) and systemic acquired resistance (SAR).Citation1 For example, tobacco and Arabidopsis plants that are SA-deficient or are unable to accumulate SA by expressing the Pseudomonas putida NahG gene (which encodes a SA degrading salicylate hydroxylase) fail to develop SAR and display enhanced susceptibility to pathogen infection.Citation2–Citation5 Similarly, SAR is compromised in the Arabidopsis isochorismate synthase (ICS)-deficient mutant, sid2, in which SA synthesis is blocked.Citation6 However, we found an interesting phenomenon that SA-deficient transgenic Arabidopsis plants NahG adapt to RNA virus infections better, which showed less-severe symptoms (less ROS accumulation and less cell death occurrence).Citation7
NahG plants and SA-deficient mutant eds5 (ENHANCED DISEASE SUSCEPTIBILITY 5) show a less-severe symptom compared with the wild-type plants Col-0 and Cvi-0 (an ecotype with a high endogenous SA level) when infected with RNA viruses, such as CMV, TCV or TuMV. Superoxide and H2O2 staining and cell-death determination indicated that they accumulated less ROS.Citation7 Virus replication levels were higher in the SA-deficient plants, than in Col-0 or Cvi-0 wild-type seedlings during the first 10 days after inoculation (dpi) suggesting the role of SA in innate immunity. However, for the long-term infections (more than 20 days), the less-severe symptoms in SA-deficient plants may eventually resulted in inhibition of viral multiplication.Citation7
The decreases in the generation of ROS and electrolyte leakage were observed in SA-deficient plants, which indicated less oxidative damages in SA-deficient plants.Citation7 Plants are capable of removing ROS using several antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX) glutathione reductase (GR) and dehydroascorbate reductase (DHAR) and so on. We found that SOD, CAT, APX activities increased significantly under virus infections, however not significantly higher in SA-deficient plants. In contrast, glutathione reductase (GR) and dehydroascorbate reductase (DHAR) activities in NahG and eds5-infected seedlings were almost twofold more than those of Col-0-infected seedlings which correspond to their higher ratios of GSH/GSSG (reduced glutathione/oxidized glutathione) and AsA/DHA (reduced ascorbic acid/dehydroascorbate). There are no distinct difference in the absolute AsA and glutathione contents between SA-deficient plants and wild-type plants.Citation7 Thus, symptom alleviation in SA-deficient plants could be attributed to their high ratios of GSH/GSSG and ASA/DHA, but not the absolute AsA and glutathione contents or heightened antioxidant enzyme activities (except GR and DHAR).
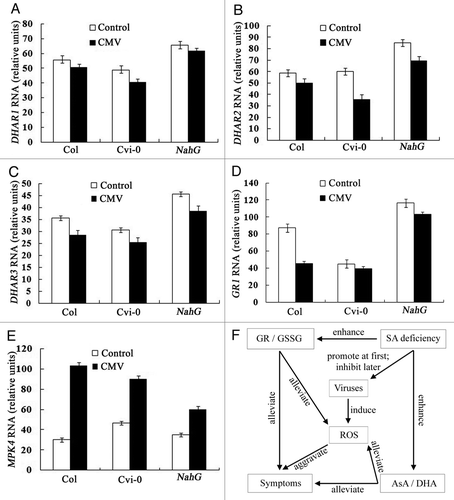
In order to further investigate the putative mechanism, DHAR1 (At1g19570), DHAR2 (At1g75270), DHAR3 (At5g16710), GR1 (At3g24170), MPK4 (mitogen-activated protein kinase 4, At4g01370) transcript levels were detected by quantitative realtime PCR analysis by comparative Ct (threshold cycle) method.Citation8 We found that DHAR1, DHAR2, DHAR3, GR1 gene expression were higher in NahG plant than in wild-type Col-0 and Cvi-0 plants when they were inoculated with or without CMV for 20 days (–D). The data were consistent with our previous report that glutathione reductase (GR) and dehydroascorbate reductase (DHAR) activities and the ratios of GSH/GSSG and AsA/DHA were higher in NahG than wildtype plants Col-0. But MPK4 gene expression was lower in NahG than in wild-type and Cvi-0 plants (). Ghanta et al. (2011), found that transgenic Nicotiana tabacum overexpressing Lycopersicon esculentum gammaglutamylcysteine synthetase gene had enhanced level of GSH in comparison with wild-type plants, exhibiting resistance to pathogenesis, which was consistent with our results.
Symptom alleviation and ROS restriction could be imitated in Arabidopsis or tobacco seedlings (Nicotiana benthamiana) by exogenously high-dose AsA treatment ().Citation7 We found that ROS eliminator treatment alone, such as dimethylthiourea (DMTU, a trap for H2O2) or Tiron (a scavenger for superoxide), could neither alleviate the symptom (necrosis and chlorosis), nor inhibit virus replication after 20 dpi.Citation7 Indeed, both necrosis and chlorosis symptom could be attributed to a kind of programmed cell death (PCD), which is mediated by multiple protein kinase cascades.Citation10,Citation11 Although AsA functions to detoxicate ROS directly, it has also been implicated in other metabolism regulation, such as violaxanthin de-epoxidase in photosynthesis, cell wall expansion, cell division and mitogen-activated protein kinases (MAPK) pathways. Therefore, AsA may prevent PCD more directly and effectively than ROS eliminators.Citation7,Citation10,Citation11
Figures and Tables
Figure 1 DHAR1 (A), DHAR2 (B), DHAR3 (C), GR1 (D) and MPK4 (E) expression levels of wild-type Col-0, Cvi-0 and NahG transgenic plants and the summarized relationship among AsA, GSH, SA deficiency, ROS, symptom and virus replication (F). Plants were inoculated with CMV or without CMV for 20 days. Transcripts were detected by quantitative real-time PCR analysis (at 20 dpi). Amplifications of Actin2/8 genes were used as an internal control. Three technical replicates were performed for each experiment. Error bars show standard deviations (n = 3).
Addendum to:
References
- Vlot AC, Dempsey DA, Klessig DF. Salicylic acid, a multifaceted hormone to combat disease. Annu Rev Phytopathol 2009; 47:177 - 206
- Durner J, Shah J, Klessig DF. Salicylic acid and disease resistance in plants. Trends Plant Sci 1997; 2:266 - 274
- Dong X. Genetic dissection of systemic acquired resistance. Curr Opin Plant Biol 2001; 4:309 - 314
- Glazebrook J. Genes controlling expression of defense responses in Arabidopsis. Curr Opin Plant Biol 2001; 4:301 - 308
- Kunkel BN, Brooks DM. Cross talk between signaling pathways in pathogen defense. Curr Opin Plant Biol 2002; 5:325 - 331
- Nawrath C, Métraux JP. Salicylic acid induction—deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 1999; 1:1393 - 1404
- Wang SD, Zhu F, Yuan S, Yang H, Xu F, Shang J, et al. The roles of ascorbic acid and glutathione in symptom alleviation to SA-deficient plants infected with RNA viruses. Planta 2011; 234:171 - 181
- Shang J, Xi DH, Xu F, Wang SD, Cao S, Xu MY, et al. A broad-spectrum, efficient and nontransgenic approach to control plant viruses by application of salicylic acid and jasmonic acid. Planta 2011; 233:299 - 308
- Ghanta S, Bhattacharyya D, Sinha R, Banerjee A, Chattopadhyay S. Nicotiana tabacum overexpressing c-ECS exhibits biotic stress tolerance likely through NPR1-dependent salicylic acid-mediated pathway. Planta 2011; 233:895 - 910
- He K, Gou XP, Yuan T, Lin HH, Asami T, Yoshida S, et al. BAK1 and BKK1 regulate brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr Biol 2007; 17:1109 - 1115
- Yuan S, Lin HH. Role of salicylic acid in plant abiotic stress. Z Naturforsch 2008; 63:313 - 320