Abstract
Spermatogenesis and male fertility are dependent upon the presence of testosterone in the testis. In the absence of testosterone or the androgen receptor, spermatogenesis does not proceed beyond the meiosis stage. The major cellular target and translator of testosterone signals to developing germ cells is the Sertoli cell. In the Sertoli cell, testosterone signals can be translated directly to changes in gene expression (the classical pathway) or testosterone can activate kinases that may regulate processes required to maintain spermatogenesis (the non-classical pathway). Contributions of the classical and non-classical testosterone signaling pathways to the maintenance of spermatogenesis are discussed. Studies that may further elaborate the mechanisms by with the pathways support spermatogenesis are proposed.
Androgens are essential for male fertility and the maintenance of spermatogenesis.Citation1,Citation2 Testosterone is the androgen in the testis that is responsible for supporting spermatogenesis. In the absence of testosterone or functional androgen receptors (AR), males are infertile because spermatogenesis rarely progresses beyond meiosis.Citation3–Citation5
Testosterone is produced by Leydig cells in the interstitial space of the testis. As a result of the local production, testosterone levels in the testis in men are 25 to 125-fold greater in the testis (340 to 2,000 nM) as compared to serum (8.7–35 nM). Testosterone levels are similarly elevated in rodent testes.Citation6–Citation10 Thus far, the specific physiologic requirements for high levels of testosterone in the testis are not known. However, it has been established that spermatogenesis does not proceed in the absence of relatively high levels of testosterone (>70 nM in the rat).Citation11
Cellular Targets of Testosterone in the Testis
AR is present in the somatic Leydig, peritubular and Sertoli cells. The localization of AR to germ cells is controversial with some studies finding AR positive germ cells and other studies showing that there is no AR in germ cells (reviewed by Wang and colleagues).Citation12 Functional evidence suggests that if AR is expressed in germ cells it is not required. Specifically, chimeric male mice having both AR defective and wild type germ cells produced pups from the AR defective germ cells.Citation13 Also, AR defective germ cells transplanted into the testes of azoospermic male mice were able to form colonies of cells undergoing spermatogenesis.Citation14 Finally, cell-specific knock out of AR in germ cells such that AR is not expressed during or after meiosis did not alter spermatogenesis or fertility indicating that AR is not required in later stage germ cells.Citation15
Sertoli cells are thought to be the major cellular target for the testosterone signaling that is required to support male germ cell development and survival.Citation16,Citation17 AR expression levels rise and fall in adult Sertoli cells in a manner corresponding with the cyclical stages of the seminiferous epithelium. In the rat, the expression of AR protein is low and difficult to detect except during stages VI–VIII when AR levels increase dramatically.Citation18 AR expression is similarly cyclical in men.Citation19 It is during stages VI–IX that the lack of testosterone or AR most affects processes required for spermatogenesis.Citation5,Citation20,Citation21
Regulation of Spermatogenesis Control Points by Testosterone and AR
Testosterone deprivation studies performed in rodents have established that testosterone is required for germ cells to progress beyond meiosis and that testosterone is required for the release of mature spermatids during stage VIII in rats (reviewed by Sharpe).Citation1 Thus far, evidence of direct testosterone support of meiosis is lacking as there are few meiosis-specific processes that are known to be directly regulated by AR-dependent actions. Instead, testosterone may act indirectly to permit germ cells to complete meiosis.
Withdrawal of testosterone or knock out of AR in Sertoli cells results in three major impairments to fertility. First, the integrity of the blood testis barrier (BTB) is compromised, which exposes post meiotic germ cells, formerly in a secluded specialized environment, to autoimmune attack and cytotoxic factors.Citation22,Citation23 Second, there is a block in conversion of round spermatids to elongated spermatids due to a defect in cell adhesion that causes the premature detachment of round spermatids from Sertoli cells.Citation21,Citation24,Citation25 Third, fully mature spermatozoa cannot be released from Sertoli cells and the germ cells are phagocytized by the Sertoli cells.Citation21
The use of Cre-Lox conditional knockout techniques to create mice in which the loss of AR is restricted to Sertoli cells (SCARKO mice) has allowed for a more precise determination of the effects of testosterone action on Sertoli cells in an otherwise normal testis. These strategies determined that, in the absence of Sertoli cell AR, spermatogenesis in mice does not progress beyond the pachytene or diplotene stages of meiosisCitation5,Citation15 and the integrity of junctional complexes making up the BTB are not maintained.Citation20,Citation26 Specifically, studies of SCARKO mice indicate that androgens regulate the expression levels of BTB tight junction-associated proteins and their localization.Citation22 Studies of cultured Sertoli cells have determined that testosterone stimulation increases the rate at which integral membrane adhesion proteins are endocytosed and then recycled to the membrane suggesting that testosterone may assist in the cyclical reformation of the BTB after the passage of leptotene spermatocytes through the barrier.Citation27 More recent studies have identified AR expression in Sertoli cells as a factor that limits the expression of differentiation markers in spermatogonial germ cellsCitation28 and that Sertoli cell nuclei show signs of immaturity and are abnormally localized away from the basal lamina.Citation22,Citation26,Citation28,Citation29 The implications of these last two characteristics of AR deficient Sertoli cells for maintaining spermatogenesis have not yet been investigated.
Classical and Non-Classical Testosterone Actions in Sertoli Cells
The classical testosterone signaling pathway.
Testosterone has been shown to act via two pathways: the classical and the non-classical.Citation30,Citation31 In the classical pathway (, left), testosterone diffuses through the plasma membrane and binds AR that is sequestered by heat shock proteins in the cytoplasm. A conformational change in AR causes the receptor to be released from heat shock proteins. AR then translocates to the nucleus where it binds to androgen response elements (AREs) in gene promoter regions, recruits co-regulator proteins and regulates gene transcription. Activation of the classical pathway requires at least 30 to 45 min to initiate changes in gene expression.Citation32
Several microarray studies using various models have been performed to survey testicular gene expression in the presence and absence of testosterone signaling (reviewed by Verhoeven and colleagues).Citation33 A broad spectrum of genes in the testis were found to be regulated by testosterone, but the number of Sertoli cell-specific genes that are regulated by testosterone make up a small subset. Furthermore, the genes identified in the microarray studies performed thus far show relatively little overlap and the number of genes displaying a two-fold or greater change in expression are limited.Citation33 Interestingly, a relatively high percentage of the regulated genes are inhibited by testosterone. Although one study determined that 65% of AR-regulated genes were linked to a conserved ARE within 6 kb of their transcription start sites, only the Rhox5 (Pem) homeobox transcription factor encoding gene, has been shown to be induced in Sertoli cells by androgens through AR binding to ARE promoter elements.Citation34 Presently, there is no evidence that any one AR-regulated gene is critical for the completion of spermatogenesis; however, it is likely that spermatogenesis would be disrupted as a result of the mutation or elimination of multiple AR-regulated genes.Citation28 Further work is required to characterize the AR-regulated genes regulated by testosterone via the classical pathway as being essential or nonessential for spermatogenesis.
Because testosterone and AR are essential for spermatogenesis and male fertility, it is surprising that the gene survey studies have not identified more testosterone-regulated genes expressed in Sertoli cells that are required for spermatogenesis. One explanation for the lack of identified genes responsible for spermatogenesis may lie in the animal models used to obtain the microarray data. Thus far, gene expression data has been obtained from either prepubertal rats and mice or from AR knock out mice in which AR expression is eliminated before birth. In both models, the testes lack full complements of germ cells, which decreases the complexity of the signals received by Sertoli cells. One solution to the problem may be to selectively knock out AR in Sertoli cells in adult mice and obtain gene expression profiles prior to the loss of germ cells. Fortunately, at least one group is developing an adult mouse model in which the AR gene can be inducibly extinguished.Citation35 Further confirmation of the importance of AR-regulated genes for maintaining fertility in mice may be obtained in the future through comparisons to genetic surveys of mutated genes found in infertile men.
The non-classical testosterone signaling pathway.
There are at least two non-classical mechanisms of testosterone action in Sertoli cells. In the testosterone-mediated [Ca2+] influx pathway (, right), testosterone rapidly induces the influx of [Ca2+] into Sertoli cells within 20–40 sec through L-Type [Ca2+] channels.Citation36,Citation37 Testosterone also is thought to cause the activation of an unidentified Gq type G protein coupled receptor and the activation of phospholipase C (PLC) that then hydrolyzes PIP2 in the plasma membrane to produce IP3 and diacylglycerol (DAG). The decrease in the levels of PIP2, an inhibitor of ATP-mediated activation of K+ATP channels, promotes the closing of these channels causing an increase in membrane resistance and depolarization of the cell. As a result, voltage dependent L-type Ca2+ channels open and allow the influx of Ca2+, which may alter many cellular processes.Citation38 Thus far, potential cellular targets and spermatogenesis processes regulated by the testosterone-mediated [Ca2+] influx pathway have not been investigated.Citation39
Testosterone also has been shown to rapidly activate a series of kinases in Sertoli cells that are known to regulate spermatogenesis. Stimulation of Sertoli cells with levels of testosterone (10–250 nM) that are similar to or lower than those found in the testis causes AR to transiently localize to the plasma membrane and results in AR interacting with and activating Src tyrosine kinase ( and middle).Citation40 Androgen stimulation triggers the direct association of the proline rich region of AR (amino acids 352–359) and the SH3 domain of Src.Citation41 Testosterone-mediated activation of Src causes the phosphorylation and stimulation of the EGF receptor (EGFR) via an intracellular pathway. Stimulation of EGFR is required to activate the MAP kinase cascade (Raf, MEK, ERK) that causes p90Rsk kinase to phosphorylate the CREB transcription factor.Citation40 Activation of the non-classical pathway has been shown to induce the expression of CREB-mediated gene expression.Citation42 In contrast to the classical pathway, induction of ERK and CREB phosphorylation by testosterone is rapid (within 1 min) and can be sustained for at least 12 hr.Citation42 The regulation of additional gene expression by other transcription factors downstream of ERK and Src remains to be investigated.
The activation of Src and Erk kinases by non-classical testosterone signaling was found to alter processes that are critical to maintain spermatogenesis. Testosterone stimulation of Sertoli cells co-cultured with germ cells from adult rats increased the numbers of germ cells attached to the Sertoli cells by 50%. However, the addition of inhibitors of Src or ERK kinase reduced germ cell attachment below basal levels.Citation40 Additional studies were performed in which AR-defective Sertoli cells were infected with adenovirus constructs expressing wild type AR or AR mutants that selectively activated only the classical pathway or the nonclassical pathway. In these studies, testosterone stimulation could only increase the attachment of germ cells to Sertoli cells expressing wild type AR or mutated AR that can stimulate the non-classical pathway.Citation40 These findings suggest that testosterone can act via the activation of Src and ERK kinases to facilitate Sertoli-germ cell attachment. It is possible that testosterone signaling that increases dramatically in stages VII–VIII of the cycle may be responsible for the remodeling of Sertoli-germ cell adhesion complexes that occurs during these stages when round spermatids begin to elongate. Furthermore, in testosterone deprived or AR deficient Sertoli cells, the lack of non-classical pathway-induced kinase activation may be responsible for the sloughing off and loss of spermatids that occurs during stages VII–VIII in the absence of testosterone signaling.
The release of mature sperm from Sertoli cells was also shown to be regulated by Src kinase that is activated by non-classical signaling. Seminiferous tubule fragments were micro dissected to isolate fragments containing only stages VII–VIII having mature elongated spermatids that were ready to be released. Culturing the seminiferous tubule fragments in the presence of a Src kinase inhibitor resulted in the release of 45% fewer sperm. These results are consistent with earlier studies showing that during stages VII–VIII when sperm are released, activated Src levels increase in the vicinity of the specialized Sertoli-elongated spematid adhesion complex called the ectoplasmic specialization (ES).Citation43–Citation45 Furthermore, Src is structurally associated with cell adhesion regulatory proteins at the ES.Citation46 Src also is known to phosphorylate focal adhesion kinase (FAK), β-catenin and N-cadherin proteins that contribute to the formation of the adhesion complexes between Sertoli cells and the mature elongated spermatids.Citation47–Citation49
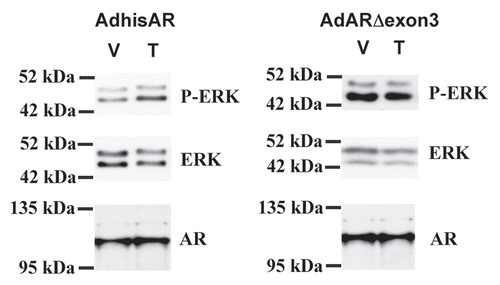
It has been proposed that only the classical pathway is required for spermatogenesis because spermatogenesis is halted during meiosis in transgenic mice in which exon 3 of the AR containing a portion of the DNA binding domain was removed.Citation50 However, the non-classical activity of the exon 3-deleted AR mutation was not tested in the study. Recreation of the AR mutant lacking exon 3 and analysis of non-classical testosterone signaling in a Sertoli cell line lacking AR activity revealed that the mutant did not permit the phosphorylation of ERK in response to testosterone stimulation (). This result indicates that the removal of more than 50 amino acids of AR in this model may alter the structure of AR to eliminate both non-classical and classical activity.
Applying Lessons Learned to Future Studies
Work is underway to better characterize the spermatogenesis processes in vivo that are regulated by the classical and non-classical pathways. Transgenic mouse models are being created in which the endogenous AR gene is removed while simultaneously initiating the expression of previously characterized mutant AR genes that are capable of only activating one of the testosterone signaling pathways (Walker WH, unpublished data). Analysis of these mouse models will determine the extent to which spermatogenesis progression is allowed by each of the pathways independently. Furthermore, the genes that are regulated by each pathway will be identified and the effects of each pathway on the maintenance of the BTB, germ cell adhesion and the release of mature sperm will be determined.
It is likely that both pathways will be found to contribute important independent regulatory actions required to maintain spermatogenesis. Signals from the two pathways also may act in concert or synergy. Data obtained from studies of progesterone, glucocorticoid and estrogen receptor function suggest that the classical and non-classical pathways cross-talk and interact in their target cells. Specifically, stimulation of cells with the steroid hormones resulted in rapid phosphorylation of their cognate receptors, which permitted the receptor to recruit co-factors resulting in the increased stimulation of specific endogenous target genes.Citation51,Citation52 Also, rapid signaling through AR has been shown to phosphorylate paxillin, which was found to contribute to testosterone-mediated transcription in prostate cells.Citation53 It is possible that the phosphorylation and activation of kinases by the nonclassical pathway in Sertoli cells may contribute to gene expression regulation via the classical pathway. In addition, important gene targets may be regulated independently downstream of the kinases that are activated via the non-classical pathway as exemplified by the androgen-mediated activation of the CREB transcription factor that is required for spermatogenesis.Citation42,Citation54
If the non-classical pathway is found to be required to maintain spermatogenesis, then it is expected that new targets for the regulation of spermatogenesis will be identified. One potential target for contraceptive development could be the testosteroneinduced interaction of AR and Src kinase that initiates the nonclassical pathway. Peptides that have already been identified to block AR-Src interactionsCitation55,Citation56 and corresponding peptidomimetic molecules are being assessed for blocking the non-classical pathway and spermatogenesis (Walker WH, unpublished data). It is possible that the partnering of factors that block the non-classical pathway with a Sertoli cell-specific delivery system could provide a hormone independent, reversible male contraceptive.
Conclusion
Although testosterone has been known to be essential for male fertility for at least 70 years,Citation57,Citation58 the molecular mechanisms by which testosterone acts to support spermatogenesis are only now being identified. The identification of testosterone-regulated genes and kinases in Sertoli cells has allowed for the discovery of the precise targets of testosterone action and a better understanding of the how testosterone regulates the process of spermatogenesis. As the molecular mechanisms of testosterone signaling continue to be revealed, we will accumulate the intellectual resources required to produce therapies for specific male infertility conditions and targets for contraceptive development.
Figures and Tables
Figure 1 Testosterone signaling pathways. (1) The classical testosterone signaling pathway. Testosterone diffuses through the plasma membrane and binds with the AR. The AR undergoes an alteration in conformation allowing it to be released from heat shock proteins in the cytoplasm. AR then is able to translocate to the nucleus where it binds to specific DNA sequences called androgen response elements (AREs). AR binding to an ARE allows the recruitment of co-activator and co-repressor proteins that alter the expression of genes to alter cellular function. (2) The non-classical kinase activation pathway: testosterone interacts with the classical AR that then is able to recruit and activate Src, which causes the activation of the EGF receptor via an intracellular pathway. The EGF receptor then activates the MAP kinase cascade most likely through Ras resulting in the sequential activation of RAF and MEK and then ERK that activates p90Rsk-kinase, which is known to phosphorylate CREB on serine 133. As a result, CREB-regulated genes such as lactate dehydrogenase A (LDH-A) and early growth response 1 (Egr1) and CREB can be induced by testosterone.Citation42 (3) The non-classical Ca2+ influx pathway: Testosterone interacts with a receptor in the plasma membrane that has characteristics of a Gq coupled G-protein coupled receptor (GPCR). Phospholipase C (PLC) is activated to cleave PIP2 into IP3 and DAG. Lower concentrations of PIP2 inhibit K+ATP channels causing membrane depolarization and Ca2+ entry via L-type Ca2+ channels.
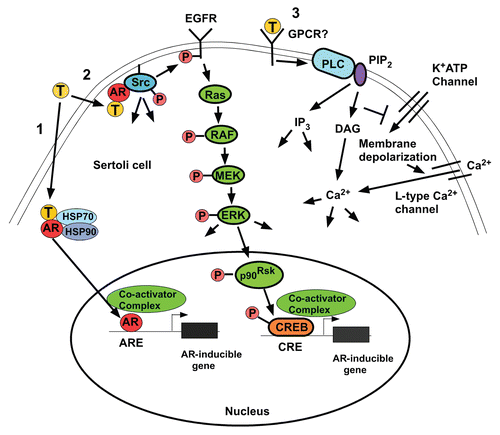
Figure 2 Deletion of exon 3 eliminates non-classical AR activity. 15P-1 Sertoli cells were infected with adenovirus constructs expressing wild type AR (AdhisAR) or exon 3-deleted AR (AdARΔexon 3). P-ER K, ER K and AR levels were determined by western blot after a 10 min stimulation in serum free media with vehicle (V) or 100 nM testosterone (T).
Acknowledgments
Research in the author's laboratory is supported by NIH grant RO1-HD43143.
References
- Sharpe RM. Knobil E, Neil JD. Regulation of spermatogenesis. The Physiology of Reproduction 1994; New York Raven Press 1363 - 1434
- McLachlan RI, O'Donnell L, Meachem SJ, Stanton PG, De Kretser DM, Pratis K, Robertson DM. Identification of specific sites of hormonal regulation in spermatogenesis in rats, monkeys and man. Recent Prog Horm Res 2002; 57:149 - 179
- Haywood M, Spaliviero J, Jimemez M, King NJ, Handelsman DJ, Allan CM. Sertoli and germ cell development in hypogonadal (hpg) mice expressing transgenic follicle-stimulating hormone alone or in combination with testosterone. Endocrinology 2003; 144:509 - 517
- Chang C, Chen YT, Yeh SD, Xu Q, Wang RS, Guillou F, et al. Infertility with defective spermatogenesis and hypotestosteronemia in male mice lacking the androgen receptor in Sertoli cells. Proc Natl Acad Sci USA 2004; 101:6876 - 6881
- De Gendt K, Swinnen JV, Saunders PT, Schoonjans L, Dewerchin M, Devos A, et al. A Sertoli cell-selective knockout of the androgen receptor causes spermatogenic arrest in meiosis. Proc Natl Acad Sci USA 2004; 101:1327 - 1332
- Comhaire FH, Vermeulen A. Testosterone concentration in the fluids of seminiferous tubules, the interstitium and the rete testis of the rat. J Endocrinol 1976; 70:229 - 235
- Turner TT, Jones CE, Howards SS, Ewing LL, Zegeye B, Gunsalus GL. On the androgen microenvironment of maturing spermatozoa. Endocrinology 1984; 115:1925 - 1932
- Awoniyi CA, Santulli R, Sprando RL, Ewing LL, Zirkin BR. Restoration of advanced spermatogenic cells in the experimentally regressed rat testis: quantitative relationship to testosterone concentration within the testis. Endocrinology 1989; 124:1217 - 1223
- Maddocks S, Hargreave TB, Reddie K, Fraser HM, Kerr JB, Sharpe RM. Intratesticular hormone levels and the route of secretion of hormones from the testis of the rat, guinea pig, monkey and human. Int J Androl 1993; 16:272 - 278
- Jarow JP, Chen H, Rosner TW, Trentacoste S, Zirkint BR. Assessment of the androgen environment within the human testis: minimally invasive method to obtain intratesticular fluid. J Androl 2001; 22:640 - 645
- Zirkin BR, Santulli R, Awoniyi CA, Ewing LL. Maintenance of advanced spermatogenic cells in the adult rat testis: quantitative relationship to testosterone concentration within the testis. Endocrinology 1989; 124:3043 - 3049
- Wang RS, Yeh S, Tzeng CR, Chang C. Androgen receptor roles in spermatogenesis and fertility: lessons from testicular cell-specific androgen receptor knockout mice. Endocr Rev 2009; 30:119 - 132
- Lyon MF, Glenister PH, Lamoreux ML. Normal spermatozoa from androgen-resistant germ cells of chimaeric mice and the role of androgen in spermatogenesis. Nature 1975; 258:620 - 622
- Johnston DS, Russell LD, Friel PJ, Griswold MD. Murine germ cells do not require functional androgen receptors to complete spermatogenesis following spermatogonial stem cell transplantation. Endocrinology 2001; 142:2405 - 2408
- Tsai MY, Yeh SD, Wang RS, Yeh S, Zhang C, Lin HY, et al. Differential effects of spermatogenesis and fertility in mice lacking androgen receptor in individual testis cells. Proc Natl Acad Sci USA 2006; 103:18975 - 18980
- Griswold MD. The central role of Sertoli cells in spermatogenesis. Semin Cell Dev Biol 1998; 9:411 - 416
- Griswold MD. Griswold MD. Perspective on the function of Sertoli cells. Sertoli Cell Biology 2005; San Diego Elsevier Science 15 - 18
- Bremner WJ, Millar MR, Sharpe RM, Saunders PTK. Immunohistochemical localization of androgen receptors in the rat testis: evidence for stage-dependent expression and regulation by androgens. Endocrinology 1994; 135:1227 - 1234
- Suarez-Quian CA, Martinez-Garcia F, Nistal M, Regadera J. Androgen receptor distribution in adult human testis. J Clin Endocrin Metab 1999; 84:350 - 358
- Meng J, Holdcraft RW, Shima JE, Griswold MD, Braun RE. Androgens regulate the permeability of the blood-testis barrier. Proc Natl Acad Sci USA 2005; 102:16696 - 16700
- Holdcraft RW, Braun RE. Androgen receptor function is required in Sertoli cells for the terminal differentiation of haploid spermatids. Development 2004; 131:459 - 467
- Willems A, Batlouni SR, Esnal A, Swinnen JV, Saunders PT, Sharpe RM, et al. Selective ablation of the androgen receptor in mouse Sertoli cells affects Sertoli cell maturation, barrier formation and cytoskeletal development. PLoS One 2010; 5:14168
- Meng J, Greenlee AR, Taub CJ, Braun RE. Sertoli Cell-Specific Deletion of the Androgen Receptor Compromises Testicular Immune Privilege in Mice. Biol Reprod 2011; http://dx.doi.org/10.1095/biolreprod.110.090621
- O'Donnell L, McLachlan RI, Wreford NG, Robertson DM. Testosterone promotes the conversion of round spermatids between stages VII and VIII of the rat spermatogenic cycle. Endocrinology 1994; 135:2608 - 2614
- O'Donnell L, McLachlan RI, Wreford NG, de Kretser DM, Robertson DM. Testosterone withdrawal promotes stage-specific detachment of round spermatids from the rat seminiferous epithelium. Biol Reprod 1996; 55:895 - 901
- Wang RS, Yeh S, Chen LM, Lin HY, Zhang C, Ni J, et al. Androgen receptor in Sertoli cell is essential for germ cell nursery and junctional complex formation in mouse testes. Endocrinology 2006; 147:5624 - 5633
- Yan HH, Mruk DD, Lee WM, Cheng CY. Blood-testis barrier dynamics are regulated by testosterone and cytokines via their differential effects on the kinetics of protein endocytosis and recycling in Sertoli cells. Faseb J 2008; 22:1945 - 1959
- Zhou W, Wang G, Small CL, Liu Z, Weng CC, Yang L, et al. Gene expression alterations by conditional knockout of androgen receptor in adult Sertoli cells of Utp14b jsd/jsd (jsd) mice. Biol Reprod 2011; 84:400 - 408
- Wang G, Weng CC, Shao SH, Zhou W, de Gendt K, Braun RE, et al. Androgen receptor in Sertoli cells is not required for testosterone-induced suppression of spermatogenesis, but contributes to Sertoli cell organization in Utp14b jsd mice. J Androl 2009; 30:338 - 348
- Walker WH. Molecular mechanisms of testosterone action in spermatogenesis. Steroids 2009; 74:602 - 607
- Walker WH. Non-classical actions of testosterone and spermatogenesis. Philos Trans R Soc Lond B Biol Sci 2010; 365:1557 - 1569
- Shang Y, Myers M, Brown M. Formation of the androgen receptor transcription complex. Mol Cell 2002; 9:601 - 610
- Verhoeven G, Willems A, Denolet E, Swinnen JV, De Gendt K. Androgens and spermatogenesis: lessons from transgenic mouse models. Philos Trans R Soc Lond B Biol Sci 2010; 365:1537 - 1556
- Lindsey JS, Wilkinson MF. Pem: a testosterone- and LH-regulated homeobox gene expressed in mouse Sertoli cells and epididymus. Dev Biol 1996; 179:471 - 484
- Willems A, De Gendt K, Deboel L, Swinnen JV, Verhoeven G. Emerging New Models to Study Androgen Action in Adult Spermatogenesis. Poster presentation, North American Testis Workshop 2011;
- Gorczynska E, Handelsman DJ. Androgens rapidly increase the cytosolic calcium concentration in Sertoli cells. Endocrinology 1995; 136:2052 - 2059
- Lyng FM, Jones GR, Rommerts FFG. Rapid androgen actions on calcium signaling in rat Sertoli cells and two human prostatic cell lines: Similar biphasic responses between 1 picomolar and 100 nanomolar concentrations. Biol Reprod 2000; 63:736 - 747
- Von Ledebur EI, Almeida JP, Loss ES, Wassermann GF. Rapid effect of testosterone on rat Sertoli cell membrane potential. Relationship with K+ATP channels. Horm Metab Res 2002; 34:550 - 555
- Rahman F, Christian HC. Non-classical actions of testosterone: an update. Trends Endocrinol Metab 2007; 18:371 - 378
- Cheng J, Watkins SC, Walker WH. Testosterone Activates MAP Kinase Via Src Kinase and the EGF Receptor in Sertoli Cells. Endocrinology 2007; 148:2066 - 2074
- Migliaccio A, Castoria G, Di Domenico M, de Falco A, Barone MV, Ametrano D, et al. Steroid-induced androgen receptor-oestradial receptor β-Src complex triggers prostate cancer cell proliferation. EMBO J 2000; 20:5406 - 5417
- Fix C, Jordan C, Cano P, Walker WH. Testosterone activates mitogen-activated protein kinase and the cAMP response element binding protein transcription factor in Sertoli cells. Proc Natl Acad Sci USA 2004; 101:10919 - 10924
- Chapin RE, Wine RN, Harris MW, Borchers CH, Haseman JK. Structure and control of a cell-cell adhesion complex associated with spermiation in rat seminiferous epithelium. J Androl 2001; 22:1030 - 1052
- Zhang J, Wong CH, Xia W, Mruk DD, Lee NP, Lee WM, Cheng CY. Regulation of Sertoli-germ cell adherens junction dynamics via changes in protein-protein interactions of the N-cadherin-beta-catenin protein complex which are possibly mediated by c-Src and myotubularin-related protein 2: an in vivo study using an androgen suppression model. Endocrinology 2005; 146:1268 - 1284
- Wang WNWR, Chapin RE. Rat testicular src: Normal distribution and involvement in ethylene glycol monomethyl ether-induced apoptosis. Toxicology and Applied Pharmacology 2000; 163:125 - 134
- Lee NP, Cheng CY. Protein kinases and adherens junction dynamics in the seminiferous epithelium of the rat testis. J Cell Physiol 2005; 202:344 - 360
- Roura S, Miravet S, Piedra J, Garcia de Herreros A, Dunach M. Regulation of E-cadherin/Catenin association by tyrosine phosphorylation. J Biol Chem 1999; 274:36734 - 36740
- Kinch MS, Clark GJ, Der CJ, Burridge K. Tyrosine phosphorylation regulates the adhesions of ras-transformed breast epithelia. J Cell Biol 1995; 130:461 - 471
- Xia W, Cheng CY. TGFbeta3 regulates anchoring junction dynamics in the seminiferous epithelium of the rat testis via the Ras/ERK signaling pathway: An in vivo study. Dev Biol 2005; 280:321 - 343
- Lim P, Robson M, Spaliviero J, McTavish KJ, Jimenez M, Zajac JD, et al. Sertoli cell androgen receptor DNA binding domain is essential for the completion of spermatogenesis. Endocrinology 2009; 150:4755 - 4765
- Chen W, Dang T, Blind RD, Wang Z, Cavasotto CN, Hittelman AB, et al. Glucocorticoid receptor phosphorylation differentially affects target gene expression. Mol Endocrinol 2008; 22:1754 - 1766
- Hagan CR, Regan TM, Dressing GE, Lange CA. ck2-Dependent Phosphorylation of Progesterone Receptors (PR) on Ser81 Regulates PR-B Isoform-Specific Target Gene Expression in Breast Cancer Cells. Mol Cell Biol 2011; 31:2439 - 2452
- Sen A, O'Malley K, Wang Z, Raj GV, Defranco DB, Hammes SR. Paxillin regulates androgen- and epidermal growth factor-induced MAPK signaling and cell proliferation in prostate cancer cells. J Biol Chem 2010; 285:28787 - 28795
- Scobey MJ, Bertera S, Somers JP, Watkins SC, Zeleznik AJ, Walker WH. Delivery of a cyclic adenosine 3′,5′-monophosphate response element binding protein (CREB) to seminiferous tubules results in impaired spermatogenesis. Endocrinology 2001; 142:948 - 954
- Migliaccio A, Varricchio L, De Falco A, Castoria G, Arra C, Yamaguchi H, et al. Inhibition of the SH3 domain-mediated binding of Src to the androgen receptor and its effect on tumor growth. Oncogene 2007; 26:6619 - 6629
- Auricchio F, Migliaccio A, Castoria G. Sex-steroid hormones and EGF signalling in breast and prostate cancer cells: targeting the association of Src with steroid receptors. Steroids 2008; 73:880 - 884
- Walsh E, Cuyler W, McCullagh D. Physiologic maintenance of male sex glands: effect of androtin on hypophysectomized rats. Am J Physiol Lung Cell Mol Physiol 1934; 107:508 - 512
- Nelson W, Merkel C. Maintenance of spermatogenesis in hypophysectomized mice with androgenic substances. Proc Soc Exp Biol Med 1938; 38:737 - 740