Abstract
During spermatogenesis, spermatogonial stem cells, undifferentiated and differentiated spermatogonia, spermatocytes, spermatids and spermatozoa all express specific antigens, many of which transiently, yet the functions of many of these antigens remain unexplored. Studies in the past three decades have shown that many of these transiently expressed genes in developing germ cells during spermatogenesis are proto-oncogenes and oncogenes, which are expressed only in the testis and various types of cancers in both humans and rodents. As such, these antigens are designated cancer/testis antigens (CT antigens). Since the early 1980s, about 70 families of CT antigens have been identified with over 140 members are known to date. Due to their restricted expression in the testis and in various tumors in humans, they have been used as the target of immunotherapy, and multiple clinical trials at different phases are being conducted with some promising results. Interestingly, in a significant number of cancer patients, antibodies against some of these CT antigens were detected in their sera. However, antibodies against these CT antigens in humans under normal physiological conditions have yet to be reported even though many of these antigens are residing outside of the blood-testis barrier (BTB), such as in the basal compartment of the seminiferous epithelium and in the stem cell niche in the testis. In this review, we summarize latest findings in the field regarding several selected CT antigens which may be intimately related to spermatogenesis due to their unusual restricted expression during different discrete events of spermatogenesis, such as cell cycle progression, meiosis, and spermiogenesis. This information should be helpful to investigators in the field to study the roles of these oncogenes in spermatogenesis.
Introduction
Spermatogenesis is the major physiological event that occurs in the seminiferous tubules, the functional units of the testis that produce spermatozoa in men as well as in rodents.Citation1 Each man produces an upward of 100 million spermatozoa each day since puberty at ∼12–14 years of age through his entire adulthood under the influence of follicle stimulating hormone (FSH) and luteinizing hormone (LH) released from the pituitary gland.Citation1–Citation4 FSH exerts its effects on Sertoli cells in the seminiferous epithelium to maintain spermatogenesis via FSH receptors which are restricted to the Sertoli cell in the mammalian body.Citation1,Citation5–Citation7 On the other hand, LH binds to the LH receptors residing in Leydig cells in the interstitial space between seminiferous tubules to induce steroidogenesis for the production of testosterone and estradiol-17β.Citation2,Citation3,Citation8–Citation12 Since the release of these two pituitary hormones that regulate spermatogenesis is under the influence of the hypothalamus, the intricate hormonal and functional relationship between the hypothalamus, pituitary gland and the testis is known as the hypothalamic-pituitary-testicular axis. This axis is also the target of male hormonal contraception by using testosterone- or testosterone-progesterone-based injectables, pills, patches, and/or implants.Citation13–Citation18 While the hormonal events along the hypothalamic-pituitary-testicular axis and the detailed morphological events that occur in the seminiferous epithelium known as the seminiferous epithelial cycle have been eminently described and reported almost 70–80 years ago,Citation2,Citation3,Citation10,Citation12,Citation19–Citation23 the molecular and biochemical mechanism(s) that regulate spermatogenesis in the seminiferous epithelium remain largely unexplored. As a result, this poses major hurdles in developing safe and reliable male contraceptives in particular non-hormonal products.Citation24–Citation27
While spermatogenesis is composed of a series of highly complex cellular events, it can be broadly divided into several discrete events: (1) renewal of spermatogonial stem cells and spermatogonia via mitosis, (2) proliferation (via mitosis) and differentiation of spermatogonia through type A and type B, and eventually preleptotene spermatocytes, (3) meiosis, (4) spermiogenesis (transformation of round spermatids to elongated spermatids and spermatozoa) and (5) spermiation (the release of sperm from the epithelium into the tubule lumen).Citation1,Citation21,Citation28,Citation29 depicts the cross-section of an adult rat testis applicable to other mammalian testes, showing sections of several seminiferous tubules at various stages of the seminiferous epithelial cycle of spermatogenesis. This figure also illustrates germ cells at different stages of their development are intimately associated and supported by Sertoli cells, which, in turn, constitute the seminiferous epithelium overlying the tunica propria. The tunica propria is composed of an acellular zone of basement membrane (a modified form of extracellular matrix which is mainly composed of laminins and type IV collagenCitation30,Citation31), and type I collagen layer, underneath of which is a cellular zone composed of peritubular myoid cell layer and lymphatic vessel (). In addition, the seminiferous epithelium is anatomically divided into the apical (adluminal) and basal compartments by the blood-testis barrier (BTB), which is located near the basement membrane in the mammalian testis. The BTB is formed by specialized junctions at the adjacent Sertoli cells in the seminiferous epithelium, which are composed of coexisting tight junction (TJ), basal ES [basal ectoplasmic specialization, a testis-specific atypical adherens junction (AJ) typeCitation31–Citation33], gap junction and desmosome.Citation32,Citation34,Citation35 This ultrastructure segregates the events of meiosis I and II, spermiogenesis and spermiation from the host animal's immune system, so that all the post-meiotic spermatids are developed in a specialized microenvironment, namely the adluminal compartment, behind the BTB. The BTB, together with the secretory products from Sertoli cells, such as interleukins and interferons,Citation36–Citation38 thus confer an immune privilege status to the testis. This barrier is physiologically significant since during each of the five different cellular events that encompass spermatogenesis, numerous germ cell-specific antigens express transiently, some of which are residing on the cell surface. Thus, the BTB “seals” the exposure of these antigens to the host immune system to avoid the production of anti-sperm antibodies, which, if occurs, will lead to infertility in men. Additionally, Sertoli cells also secrete humoral factors (e.g., interleukins, interferons and cytokines) to confer an immune privilege status to the testis.Citation35,Citation39–Citation41
Interestingly, studies in past 15 years have shown that developing germ cells express a large number of genes, many of which were subsequently shown to be oncogenes and/or proto-oncogenes, which are either not expressed or expressed at exceedingly low level in normal mammalian cells unless they are induced and/or transformed to undergo carcinogenesis. However, normal testis per se is not a tumor, and many of the developing germ cells, including spermatogonia, spermatocytes, round and elongating spermatids only express these genes transiently, such as at specific steps of spermatids during spermiogenesis and differentiating spermatogonia, as well as differentiating spermatocytes that are preparing for meiosis. Also, greater than 75% of the developing germ cells undergo apoptosis to avoid overwhelming the limited number of Sertoli cells (estimated to be ∼25–40 million per testis in the rat.Citation42–Citation44 Sertoli cells cease to divide by about 17 days postpartum (dpp) in ratsCitation45 even though recent studies have shown that adult mouse and human Sertoli cells can become proliferative when cultured in vitro under certain conditionsCitation46,Citation47). Furthermore, some of the transiently expressed proteins have crucial functions to spermatogenesis (e.g., cell adhesion) and reproductive processes (e.g., sperm-egg interactions) (see sections below), yet they were subsequently found to be expressed abundantly only in metastatic or tumor cells but not in any other normal cells and/or tissues besides the testis and the placenta (and the ovary in some cases). Thus, these antigens are known as cancer/testis (CT) antigens.Citation48,Citation49 In short, the testis is not a tumor per se, but it does express many of the oncogenes that are restricted to tumorigenesis and metastasis. Herein, we review some of the recent findings regarding the roles of CT antigens in spermatogenesis and carcinogenesis. A better understanding on this group of proteins will not only unfold their roles in the seminiferous epithelial cycle of spermatogenesis but will also provide us with a clearer insight into how to target immunotherapy towards CT antigens in cancer.
Cancer/Testis (CT) antigens
The term “Cancer/Testis (CT) antigen (or CTA)” was first used in 1997.Citation50 CT antigens, also known as cancer germline antigens, refer to a growing list of antigens that were initially discovered in the 1980s–1990s,Citation50–Citation57 which specifically expressed in various tumors of different histological origins. CT antigens are absent in normal somatic cells in humans and rodents but expressed only in male germ cells (such as spermatogonial stem cells, spermatogonia, spermatocytes, spermatids and spermatozoa during spermatogenesis) in the testis (but not Sertoli and/or Leydig cells).Citation58–Citation63 In some cases, CT antigens are also expressed in ovary and in the placenta (usually in trophoblast cells), such as NY-ESO-1, a CT antigen originally identified from esophageal squamous cell carcinoma of a 58-year-old female cancer patient.Citation50,Citation58–Citation63 More importantly, the genes of CT antigens are activated and highly expressed in a number of malignant tumors and they are antigenic in tumor-bearing patients.Citation50,Citation60 Thus, many CT antigens were identified by using sera of cancer patients to screen human testis cDNA library or recombinant proteins.Citation50,Citation59,Citation64,Citation65
CT antigens are broadly divided into those that are encoded by genes residing on the X chromosomes, the X-CT antigens, and those that are not, the non-X-CT antigens.Citation59 However, X-CT antigens represent more than half of all CT antigens identified to date (). They are often organized in well-defined clusters to constitute multigene families along the X-chromosome, where different members of a single family are arranged into complex direct and inverted repeats.Citation48,Citation59,Citation66 The genes encoding non-X-CT antigens, however, are distributed throughout the genome and are mostly single-copy genesCitation59 (). While the functions of many of these CT antigens remain unknown, recent studies showed that these proteins are likely involved in cell cycle progression/regulation, transcriptional control, cell survival and apoptosis. Studies on the expression of CT antigens have also shown that epigenetic events, such as DNA methylation, are the primary mechanism regulating the expression of CT antigens in germ cells and transformed cells.Citation48 Also, CT antigens (e.g., MAGE proteins) are frequently mutated in cancers,Citation67 and many CT antigens were identified by using sera of cancer patients to screen human testis cDNA library or recombinant proteins,Citation50,Citation59,Citation64,Citation65 illustrating they are highly antigenic. In light of their restricted expression in tumor and cancer stem cells and their antigenic properties, several CT antigens have been the targets of cancer immunotherapy (e.g., vaccine therapy) and some of which are biomarkers of various malignancies, such as ovarian cancer, cervical cancer, breast cancer, lung cancer, urinary bladder cancer, osteosarcoma and testicular cancer.Citation48,Citation60,Citation68–Citation74 Unfortunately, some of the recently completed clinical trials revealed that even though the immunotherapy that targets CT antigens could shrink the tumor, it failed to increase the survival rate of the treated patients versus patients that received placebo,Citation48,Citation75 illustrating the complexity of CT antigens.Citation48 At present, there are ∼70 families of CT antigens and about 140 members have been identified, with their functions largely unknown.Citation48,Citation60 Since each cancer type is associated with multiple highly expressed CT antigens, an effective vaccine may require the presence of multiple CT antigens. In this review, we summarize some of the latest findings in the field regarding the roles of several best studied CT antigens during tumorigenesis. These selections were based on their intrinsic activities, mode of actions and/or immunohistochemical localization patterns in human testis, illustrating they are intimately related to spermatogenesis, such as their assoication with spermatogonia, spermatocytes, post-meiotic spermatids and spermatozoa. These findings thus illustrate the possible roles of CT antigens during spermatogenesis, and functional experiments can be designed in future studies ().
CT Antigens: Their Roles in Spermatogenesis and Tumorigenesis
Sperm protein 17 (SP17).
SP17 was originally identified as an autoantigen in the mouse testis. It is highly expressed in mouseCitation76 and humanCitation77 spermatozoa and is restricted to the tail (in particular the fibrous sheathCitation78) and over the acrosomal region of the head of mature spermatozoaCitation76 (). The primary sequence of SP17 is composed of three domains: (1) a conserved N-terminal domain that shares homology with type IIα regulatory subunit of protein kinase A (PKA-RIIα) that binds A-kinase anchoring protein 3 (AKAP3) in sperm flagella with high affinity (note: AKAP3 is involved in regulating human sperm motility following its coupling to the regulatory subunit of PKA.Citation79,Citation80 AKAP3 is also a CT antigen restricted to human testis in normal tissue but highly expressed in tumors and its expression is correlated with a poor prognosis and low survival rate in ovarian cancer patientsCitation60) (2) a central sulfated carbohydrate (heparin)-binding domain which contains two putative heparin-binding motifs (BBXB, where B is any basic amino acid, X is any amino acid) of KREK (residues 49–52) and KKIR (residues 131–134), and (3) a Ca2+/calmodulin binding domain near its C-terminus.Citation81–Citation83 Its functions remain unknown except it was implicated to be: (1) involved in sperm-egg interactions between the acrosome-reacted sperm and zona pellucida of the ovum during fertilizationCitation84,Citation85 in light of its restricted localization at the acrosome of normal and capacitated spermatozoa undergoing acrosome reactionCitation76 and (2) involved in cell adhesion and/or cell migration possibly through the interaction of its heparin-binding motifs with heparan sulfate in extracellular matrix (e.g., in malignant lymphocytes).Citation86 Also, Sp17 can serve as a scaffolding protein to form a PKA-independent AKAP protein complex via its AKAP-binding motif to recruit additional signaling proteins to specific cellular site(s) in both germ cells and malignant cells.Citation87
A mouse homolog of SP17 was found to be highly expressed in metastatic cell lines derived from a mouse model of squamous cell carcinoma but not in the nonmetastatic parental line,Citation88 illustrating the potential of SP17 as a therapeutic target. Subsequent studies have shown that SP17 was highly expressed in cancers of unrelated histological origins, such as multiple myeloma, ovarian cancer, brain tumors, esophageal cancer, ovarian cancer, endometrial cancer and cervical cancer.Citation73,Citation89–Citation94 Thus, gynecological cancers appeared to be the best candidates for SP17 targeted immunotherapy because it is virtually absent in normal tissues in women.Citation95 Unfortunately, subsequent studies have identified that SP17, unlike most CT antigens, is expressed also in somatic cells including the ciliated epithelia of the respiratory airways and reproductive tract of both men and women,Citation96 synoviocytes from patients of rheumatoid arthritis,Citation97 and melanophages of cutaneous melanocytic lesions.Citation98 These findings thus raise a question as to whether SP17 can be a target for immunotherapy due to its wide-spread presence in other epithelial cells. Thus, no clinical trials have been performed using SP17 as an immunotarget. Interestingly, in a mouse model of ovarian cancer, vaccination of recombinant SP17 protein was found to provide long-term protective effect on reducing the tumor size and controlling tumor growth in mice.Citation99 Nonetheless, a better understanding of SP17 in the testis, in particular in spermatogenesis, such as its involvement in spermiogenesis, may offer hints on the roles and functions of this protein in tumorigenesis and/or metastasis.
NY-ESO-1.
NY-ESO-1, also known as CTAG1B (cancer/testis antigen 1B), is a 22 kDa protein that was originally found and isolated from a New York City 58-year-old female cancer patient of esophageal squamous cell carcinoma.Citation50 NY-ESO-1 together with LAGE-1 (also known as CTAG2, cancer/testis antigen 2Citation100 which shows no expression in normal tissues other than the testisCitation60) constitute the NY-ESO-1/LAGE family of CT antigensCitation59,Citation66 (). NY-ESO-1 is one of the best studied CT antigens and its mRNA is highly expressed in the testis [restricted mostly to spermatogonia and primary spermatocytes but not in post-meiotic spermatids nor Sertoli cells in humansCitation101,Citation102 ()], ovary (but not in any normal tissues) and human melanoma, breast cancer, bladder cancer, gastric cancer, lung cancer, sarcoma, ovarian cancer, prostate cancer and hepatocellular carcinoma.Citation50,Citation103,Citation104 Due to its unusual immunogenic nature to induce high titer of autologous antisera in patients,Citation50 NY-ESO-1 has been actively investigated to serve as a suitable target of immunotherapy in NY-ESO-1 expressing tumorsCitation63,Citation105 (). For instance, anti-NY-ESO-1 antibody was detected in the serum of ∼10–86% of patients of various cancers, which include thyroid cancers, lung cancers, ovarian cancers, breast cancers, bladder cancers, esophageal cancers and melanomas.Citation48 About 35 Phase 1 clinical trials have been or are being conducted with His-tagged recombinant NY-ESO-1 worldwide (including the US, Japan and Germany) using different adjuvants (e.g., saponin-based adjuvant, ISCOMATRIX) and in different cancer types (e.g., melanomas).Citation48 Despite detecting strong antibody responses, no favorable clinical response was noted in advanced melanoma patients in response to NY-ESO-1 ISCOMATRIX vaccine.Citation106 However, a study reported in 2008 has shown that ipilimumab [an anti-CTLA-4 (CTLA-4, cytotoxic T lymphocyte-associated antigen 4 found on cytotoxic T lymphocytes that plays a critical role in regulating natural immune responsesCitation107) monoclonal antibody (marketed as Yervoy by Bristol-Myers Squibb for advanced metastatic melanomas)] therapy combined with NY-ESO-1 vaccine can synergistically enhance the clinical effects in treating advanced metastatic melanomas, since ipilimumab was found to enhance polyfunctional NY-ESO-1-specific T-cell responses to combat metastatic cancer cells.Citation108 A recent study using a synthetic peptide based on NY-ESO-1 (ESO157–165, residues 157–165) was vaccinated in combination with Incomplete Freund's Adjuvant (Montanide ISA-51) and CpG7909 ODN (deoxycytidyl-deoxyguanosin oligodeoxy nucleotides) to a group of 14 patients with melanoma (n = 7), non-small cell lung cancer (n = 4), ovarian cancer (n = 1), sarcoma (n = 1) and breast cancer (n = 1) with confirmed metastatic and Stages III/IV malignancies. It was found that the vaccine is able to induce anti-NY-ESO-1 humoral and T-cell mediated response, and positively correlates with survival.Citation104 However, in a subsequent clinical study in which advanced esophageal (n = 8) and prostate cancer patients (n = 2) were vaccinated using a complex of cholesterol-bearing hydrophobized pullalan (CHP) and NY-ESO-1 protein (CHP-NY-ESO-1) to efficiently induce NY-ESO-1 antibody, which in turn activated CD4 and CD8 T cell responses, and associated with strong heteroclitic serological responses against 11 tumor antigens (including MAGE-A1, MAGE-A3, MAGE-A4, CT7/MAGEC1, CT10/MAGEC2, CT45, CT46/HORMAD1, SOX2, SSX2, XAGE1B and p53). However, the tumor size did not alter, and most subjects in the study eventually died, rendering the vaccine to be ineffective.Citation75 Interestingly, no functional study was found in the literature to explore the physiological role of NY-ESO-1 in spermatogenesis regarding its unusual high expression in spermatogonia and primary spermatocytes () even though its expression in the testis was reported ∼15 years ago.Citation50
Melanoma-associated antigen (MAGE) gene family.
Since the identification of the first MAGE gene in a human melanoma called MAGE-A1 in 1991,Citation109 the MAGE family has expanded to include over 60 genes.Citation110,Citation111 MAGE genes, as the name implies, are highly expressed in melanoma in particular metastatic melanoma and by germ cells in the testis.Citation111,Citation112 All members of MAGE genes share a conserved stretch of sequence of about 200 amino acids known as the MAGE homolog domain (MHD), usually located close to the C-terminus.Citation111 Initial studies reported that MAGE genes (e.g., MAGE-A1) were silenced in normal adult tissues except male germ cells and, for some of them in the placenta. They have been targets for immunotherapy as they were highly expressed in different types of tumors (e.g., MAGE-A3).Citation63 However, similar to SP17, a few members of the MAGE family were subsequently found to be expressed in normal cells.Citation52,Citation113 Interestingly, MAGE-like genes are found in non-mammalian species, including Drosophila melanogaster (Fruit fly) and Danio retrio (Zebrafish), but not in Caenorhabditis elegans (Roundworm), Saccharomyces cerevisiae (Baker's yeast) or Schizosaccharomyces pombe (Fission yeast), indicating that MAGE genes might have evolutionarily conserved functions and recent studies have shown that MAGE sequences are also found in some flowering plants, such as Arabidopsis thaliana (Mouse-ear cress).Citation110
The MAGE family is now composed of two subfamilies MAGE-I and MAGE-II.Citation111 MAGE-I family consists of chromosome X-clustered genes including MAGE-A, MAGE-B and MAGE-C groups.Citation111 Most MAGE-I family members are CT antigens, rarely expressed in normal adult tissues except germ cells in the testis, but highly expressed in different types of cancer.Citation52,Citation59 MAGE-I proteins are crucial to tumorigenesis and cancer cell survival. For instance, MAGE-A3 was found to bind to procaspase-12 to block its activation to caspase-12. This disrupts the caspase-12-mediated cell apoptosis in response to drug-induced insults, resulting in enhanced survival of cancer cells as the result of increased expression of MAGE-A3.Citation114 Recent studies have also shown that several MAGE proteins interact with RING (Really Interesting New Gene, a protein structural domain of zinc finger type containing a Cys3HisCys4 amino acid motif that binds to two zinc cations which plays a key role in protein ubiquitination pathway and cell proliferationCitation115,Citation116) domain-containing proteins (e.g., E3 ubiquitin ligases) and can modify and adjust E3 ubiquitin ligase activity.Citation117 Since MAGE-A antigens are not expressed in normal tissues except germ cells in the testis, they are the targets of immunotherapy. Based on the promising findings using a MAGE-A3-based vaccine from a randomized Phase II study in patients with metastatic melanoma and a double-blinded placebo-controlled Phase II study in patients with resected lung cancer, two Phase III clinical trials have been initiated in patients with melanoma skin cancer and non-small cell lung cancer, which are known as DERMA (Adjuvant Immunotherapy with MAGE-A3 in melanoma) and MAGRIT (MAGE-A3 as Adjuvant Non-Small Cell Lung Cancer Immunotherapy), respectively ().Citation111,Citation118,Citation119 In these MAGE-A3 Antigen-Specific Cancer Immunotherapeutic (ASCI) trials, MAGE-A3 recombinant protein and a potent immunostimulant AS15 were used to formulate a vaccine and the results of these two Phase 3 trials will be known by the end of this decade.
On the other hand, members of MAGE-II family (e.g., MAGE-D proteins) are expressed in normal tissues and they are not related to cancer even though some of them are also located on X chromosome. Interestingly, each member also contains a MDH domain (MAGE Homology Domain, a signature domain for MAGE proteins) but the MDH domain is located more distant from the N-terminus.Citation111 MAGE-D1 gene, unlike members of MAGE-I family, is downregulated in tumor cells. In addition, protein of MAGE-D1 gene, Maged1, was found to play a role in anti-tumorigenesis in different cell types (and it was also shown to regulate circadian clock functions via its extensive network of interacting partner proteins).Citation120 Maged1 is also a binding partner of 75 kD low-affinity neurotrophin receptor (p75NTR), the apoptosis inhibitory protein XIAP, and Dix/MSX homeodomain protein, involving in blocking cell cycle progression and enhancing apoptosis.Citation121 Necdin, another MAGE-II family member, is a neuron-specific growth suppressor and binds to a cellular transcription factor E2F-1 (E2 promoter binding factor-1), which regulates cell cycle-specific genes.Citation122,Citation123 Necdin is known to interact with neurotrophic tyrosine kinase receptor type 1 (TrkA) and p75NTR, and its deletion on the chromosome 15 is associated with Prader-Willi syndrome (PWS, a neurobehavioral disorder and the genetic cause of life-threatening obesity in children).Citation124,Citation125 Necdin, similar to Maged1, is also downregulated in primary tumors suggesting a tumor suppressor role, which involves in hematopoietic stem cell quiescence and transcriptional repression.Citation124 In short, these findings illustrate that while MAGE proteins are CT antigens, members of the MAGE-II family are having anti-tumor properties.
SSX (synovial sarcoma X).
Human synovial sarcoma is an aggressive malignant soft tissue tumor with high recurrence and metastasis as the result of chromosomal translocation. It is characterized by its unique (X;18)(p11.2;q11.2) chromosomal translocation leading to the formation of the SS18-SSX fusion gene that causes tumorigenesis. In synovial sarcoma, the breakpoint-associated genes are fusion genes of SYT from chromosome 18 (also known as SS18), and SSX from the X chromosome.Citation126,Citation127 The resulting fusion protein product of SS18 (or SYT) and SSX genes encode nuclear proteins that exhibit opposite transcriptional regulatory activities, with the SS18 protein functions as a transcriptional activator and the SSX proteins as transcriptional repressors.Citation127,Citation128 It was shown that the downstream target of the SS18-SSX fusion protein is COM1, a regulator of cell proliferation, in which SS18-SSX downregulated COM1 in synovial sarcoma tissues and cell lines.Citation129 The SSX breakpoint multigene family is composed of at least 9 genes of SSX1-9, and all are located on chromosome X.Citation130 Among the 9 SSX genes, the mRNA transcripts of SSX1, 2, 3, 4, 5 and 7 (but not 6, 8 and 9) are expressed in normal human testes but not in other human tissues.Citation130 Among tumor tissues, SSX 1, 2 and 4 are frequently expressed while SSX 3, 5 and 6 are rarely expressed. For SSX 7, 8 and 9, they have yet to be detected.Citation130 Similar to other CT antigens, aberrant expression of SSX proteins during malignancy can elicit the production of antibodies. Anti-SSX1, 2, 3 and 4 antibodies have been demonstrated in sera from cancer patients with different tumors including melanoma, colon, breast and ovarian cancers.Citation131–Citation133 Thus, SSX proteins are potential targets of immunotherapy.Citation134,Citation135 However, no clinical trials have been conducted using SSX proteins as vaccines. By using a mouse monoclonal antibody that was found to cross-react with SSX2, 3 and 4 (but not SSX1), SSX proteins were found to be localized to the nucleus of spermatogonia and early spermatocytes (e.g., preleptotene, leptotene and zygotene) but not pachytene spermatocytes, round spermatids, elongating/elongated spermatids and spermatozoa in human testes.Citation136 This finding thus illustrated the possible involvement of SSX proteins in cell proliferation (e.g., self-renewal of spermatogonia stem cells and spermatogonia) and cell cycle progression. This localization pattern is also consistent with an earlier report which showed that the SS18-SSX fusion protein regulates cell proliferation via its effects on cell proliferation regulator COM1.Citation129
Synaptonemal complex protein 1 (SCP-1).
SCP-1, also known as HOM-TES-14 (), is selectively expressed during meiotic prophase of spermatocytes (from zygotene to diplotene)Citation137 in human testis at the synaptonemal complexes because it is an integrated component of the synaptonemal complexes.Citation137 It is involved in the pairing and recombination of homologous chromosomes during meiosis I.Citation137,Citation138 Unlike other CT antigens which are located on chromosome X, SCP-1 is localized on chromosome 1.Citation138 SCP-1 is also expressed in neoplastic tissues and tumor cell lines, including malignant gliomas, breast, renal cell and epithelial ovarian cancer.Citation138,Citation139 Its elevated expression in epithelial ovarian cancer (EOC) is associated with poor prognosis and reduced survival in EOC patients, and as such, it is being considered a target of immunotherapy.Citation140 Its restricted expression in pre-meiotic spermatocytes for chromosomal pairing illustrates its significance in meiosis. However, in cancer cells, SCP-1 displayed a cell cycle phase-independent nuclear expression in cancer cells.Citation138 Future studies should include studies to examine the role of SCP-1 in mitosis in tumor cells.
Piwil2.
Piwi (P-element induced wimpy testis was first identified in DrosophilaCitation141,Citation142) is a class of genes encoding regulatory proteins responsible for maintaining differentiation and self-renewal in stem cells, RNA silencing and translational regulation.Citation142–Citation145 They are members of the Argonaute family which are highly conserved among speciesCitation146 and are the catalytic components of RISC (RNA-induced silencing complex) which is responsible for RNAi to silence gene in normal eukaryotic cells to maintain cell homeostasis.Citation147 Argonaute proteins bind small non-coding RNAs such as microRNAs (miRNAs), small interfering RNAs (siRNAs) and Piwi-interacting RNAs (piRNAs).Citation145,Citation148–Citation151 Miwi is the murine homolog of Piwi (and Hili is the human homolog of Piwi) and it was found to encode a cytoplasmic protein specifically expressed in spermatocytes and spermatids that is essential for spermatogenesis, since Miwi−/− mice displayed spermatogenic arrest at round spermatid, disrupting spermiogenesis.Citation152 Among the genes of the Piwi family, they all contain the conversed PAZ domain (a ∼110-amino acid stretch of sequence found in Piwi, Argo and Zwille/Pinhead proteins in C. elegans, Drosophila and humans) and Piwi domain (Piwi is a protein domain homologous to piwi proteins that bind and cleave RNA).Citation153 A stem-cell protein Piwil2 (Piwi-like 2) was found to be specifically expressed in the testis of humans and mice, restricted to spermatogonia and early spermatocytes, with enhanced expression in human testicular seminomas (but not in testicular non-seminomatous tumors) and also in different tumors, including prostate, breast, gastrointestinal, ovarian and endometrial cancer of humans.Citation154 As such, Piwil2 is a CT antigen and an oncogene.Citation60 Piwil2 exerts its effects by inhibiting cell apoptosis and promoting cell proliferation via the Stat3/Bcl-XL signaling pathway.Citation154 A recent study has shown that Piwil2 regulates chromatin modifications such as chromatin condensation that affects DNA repair and its overexpression in cancer cells could lead to cellular cisplatin resistance.Citation155 Several Piwil2-like (PL2L) proteins have recently been found in various human tumors that promote tumor cell survival and proliferation via the Stat3/Bcl2 pathway.Citation156 Thus, by blocking the expression of this gene during tumorigenesis would reduce the uncontrolled expansion of malignant cells and since it is a CT antigen, it can become a target of immunotherapy and/or diagnostic/prognosis biomarker.
ACRBP (acrosin binding protein).
ACRBP, also known as OY-TES-1 or CT13 (cancer/testis antigen 23), is an acrosin binding protein located on chromosome 12 ().Citation157 OY-TES-1 is the human homolog of proacrosin binding protein sp32 precursor first found in pig and mouse testis.Citation158 Besides highly expressed in the testis and localized to the mature spermatozoa at the acrosomal region and not other somatic cells (), it is expressed at high levels in malignant tissues including bladder, breast, lung, liver, colon and ovarian cancers.Citation157,Citation159,Citation160 Anti-OY-TES-1 antibody was detected in ∼10% of the ovarian cancer patients whose tumors expressed this antigen.Citation160 Its elevated expression in ovarian cancers was found to associate with reduced survival time and faster relapse in ovarian cancer patients,Citation159 illustrating OY-TES-1 can be a target of immunotherapy. A recent study has shown that the mitotic spindle protein NuMA is the binding partner of this acrosin binding protein, and as such, OY-TES-1 affects mitotic spindle assembly during tumor cell proliferation.Citation159
CT antigen expression during spermatogenesis.
The role of CT antigens in spermatogenesis remain largely obscure (with the unusual exception of SCP-1 which is known to be involved in synaptonemal complex formation) since virtually no functional studies are found in the literature to explore their roles in spermatogenesis except for several studies that examine their immunohistochemical localization in human testes (or normal rodent testes) versus patients' samples having different cancers. These findings are briefly summarized in and discussed herein. However, based on these limited findings, some obvious experiments can be conducted in future studies to understand the role of these transiently expressed cancer genes during spermatogenesis, in particular stem cell proliferation, cell cycle progression, meiosis and spermiogenesis. These studies will assist the understanding on the involvement of CT antigens in these cellular events but also enhance our understanding of the molecular mechanism(s) underlying tumorigenesis and metastasis when these CA antigens are highly expressed and/or activated.
A mouse monoclonal antibody, designated 57B, that was developed against recombinant MAGE-3 protein (i.e., an anti-MAGE-3 antibody, but it was found to cross-react also with MAGE-1, -4, -6 and -12,Citation52 which are members of the MAGE-A groupCitation161 in MAGE-I subfamily) has been used for immunohistochemistry analysis to examine the association of MAGE-3 with specific germ cell types.Citation52 MAGE-A was found to be highly expressed in spermatocytes, early spermatocytes (e.g., leptotene, zygotene and diplotene spermatocytes), but somewhat diminished in pachytene spermatocytes and round spermatids, and appeared to be localized more to the nucleus than the cytoplasm, but not expressed in elongating spermatids and spermatozoa, nor in Sertoli and Leydig cellsCitation52 (). Interestingly, this pattern of staining was almost identical in a subsequent study using a specific anti-MAGE-1 monoclonal antibody designated MA454 except that the localization of MAGE-1 is more prominent in the cytosol and at the perinuclear region of spermatogonia, spermatocytes and round spermatids.Citation162 Collectively, these findingsCitation52,Citation162 show that MAGE-3, -4, -6 and -12 may have slightly different cellular distribution, which are nuclear bound, versus MAGE-1 which was localized more to the cytoplasm. Furthermore, MAGE proteins may be intimately related to cell cycle progression, to prepare germ cells for meiosis I and II, and early phase of spermiogenesis but not involved in spermiation.
Magphinins, a new group of MAGE proteins composed of three isoforms of magphinin-α, -β and -γ, are found in the mouse testis and ovary, restricted to the male and female germ cells, regulating cell proliferation during gametogenesis.Citation163 However, magphinin-α and -β, unlike other CT antigens, are also detected in the epididymis and brain.Citation163 Overexpression of magphinin-α in GC1spg cells (GC1spg cell line was established by immortalizing spermatogenic cells with SV40 antigenCitation164) or overexpression of magphinin-α, -β or -γ in Saos-2 cells (a non-transformed cell line derived from primary osteosarcomaCitation165) were found to inhibit cell proliferation by promoting cell cycle arrest,Citation163 illustrating their involvement in cell cycle progression, and perhaps SSC/spermatogonia proliferation for their self-renewal to maintain the proper SSC population at the stem cell niche. When the localization of magphilins in the seminiferous epithelium of adult mouse testis were examined, magphinin-β was found to be abundantly expressed by spermatogonia and spermatocytes in the cytosol. On the other hand, both cytosol and nucleus were stained with magphinin-β in secondary spermatocytes and round spermatids, which persisted through elongated spermatids and spermatozoa at spermiationCitation163 (see ). Studies by immunohistochemistry have shown that NY-ESO-1 was prominently expressed by spermatogonia, early spermatocytes through pachytene spermatocytes but not in any post-meiotic spermatogenic cells including spermatids and spermatozoa in human testis, and also not in Sertoli and Leydig cellsCitation101,Citation102 (). Furthermore, this pattern of expression is somewhat similar to the expression of SCP-1Citation137 and Piwil2Citation154 based on in situ hybridization (and immunogold electron microscopy) and immunohistochemistry analysis, respectively. For SCP-1, it is a structural component of synaptonemal complexes involving in pairing of the homologous chromosomes in zygote through diplotene spermatocytesm and it is restricted mostly to these spermatocytes,Citation137 whereas Piwil2 is involved in spermatogonia and SSC renewal and differentiation and is therefore restricted to these cellsCitation154 in the human testis.
However, several other CT antigens are restrictively expressed and localized to spermatozoa in the testis such as SP17, and OY-TES-1 is restricted to elongating/elongated spermatids and spermatozoa instead of the less advanced germ cell types (). These findings regarding the expression and localization of several CT antigens in germ cells during spermatogenesis clearly illustrate that much work is needed to delineate the function of these proteins in spermatogenesis. These findings, if available, should provide insightful information regarding their role and their aberrant expression during tumorigenesis.
Concluding Remarks and Future Perspectives
As summarized above, while the physiological function for virtually all the CT antigens during spermatogenesis remain unknown, several of these CT antigens are emerging as proteins intimately related to: (1) spermatogonial stem cell/spermatogonial self-renewal via mitosis (e.g., Piwil2), (2) transcription regulation (e.g., SS18-SSX proteins), (3) chromosomal pairing in spermatocytes to prepare for meiosis I (e.g., SCP-1), (4) germ cell apoptosis (e.g., MAGE proteins. Piwil2), (5) spermiogenesis (e.g., OY-TES-1) and (6) sperm motility (e.g., SP17). In short, this information, as summarized in , thus illustrates that CT antigens are involved in virtually all the different phases of cellular events pertinent to spermatogenesis. With the recent advances in molecular and cell biology, and biochemical and molecular techniques to manipulate the expression of different genes and/or proteins such as by RNAi utilizing siRNA duplexes, shRNA, and/or lentiviruses/adenoviruses, we will gain a lot of insightful information regarding the molecular mechanism(s) by which these CT antigens regulate spermatogenesis. These findings should be helpful to design better chemotherapeutic drugs to treat different tumors in humans.
Figures and Tables
Figure 1 Morphological and cellular features of spermatogenesis in the mammalian testis. The anatomical features of the rat testis shown here (A) share many of the feature similar to other mammalian testes including humans, and the schematic drawing shown in (B) depicts the major cellular events of spermatogenesis, namely spermatogonia/SSC self-renewal via mitosis and differentiation to type B spermatogonia, which in turn transform to preleptotene spermatocytes. Preleptotene spermatocytes are the germ cells residing in the basal compartment traverse the blood-testis barrier (BTB) at stage VIII of the epithelial cycle that enter the adluminal (apical) compartment while differentiate into leptotene, zygote and then pachytene spermatocytes in a process known as cell cycle progression. Diplotene spermatocytes eventually enter meiosis I to be immediately followed by meiosis II at stage XIV as shown in (A) to form haploid round spermatids (step I spermatids). Spermatids undergo spermiogenesis to form elongating and elongated spermatids involving 19 and 16 steps in rats and mice, respectively. Elongated spermatids transform into spermatozoa following the shredding of the residual body to be phagocytosed by the Sertoli cells to allow the release of sperm at spermiation. The fine morphological features of some of these germ cells are also depicted in (B). It is noted that the seminiferous epithelium is resting on the tunica propria, which is composed of an acellular zone namely basement membrane (a modified form of extracellular matrix) and type I collagen layer, and a cellular zone of peritubular myoid cell layer and the lymphatic microvessel. It is noted that the seminiferous epithelium is composed of only Sertoli and germ cells without the presence of any blood vessels and nerve fibers since all microvessels are restricted to the interstitial space between seminiferous tubules. The BTB also physically divides the seminiferous epithelium into the basal and adluminal (apical) compartments. Different cancer/testis (CT) antigens are expressed throughout spermatogenesis with unique patterns of cellular expression among different types of germ cells. PLS, preleptotene spermatocyte; RS, round spermatid.
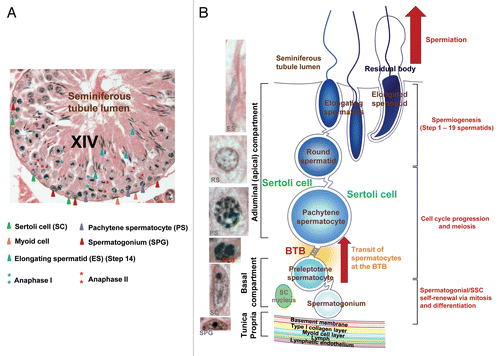
Figure 2 A schematic drawing illustrating the restricted expression of different CT antigens in germ cells during spermatogenesis. The part on the right depicts different germ cell types during spermatogenesis, and the left part illustrates the unique expression patterns of seven different CT antigens in different germ cell types, illustrating their possible involvement in different phases of spermatogenesis. These CT antigens, besides their restricted expression in the testis but not other normal human cells/tissues, they are highly expressed in cancer cells. As such, many of these antigens are the targets of immunotherapy and vaccine development for cancer treatment. Yet their functional significance in the testis remains largely unexplored.
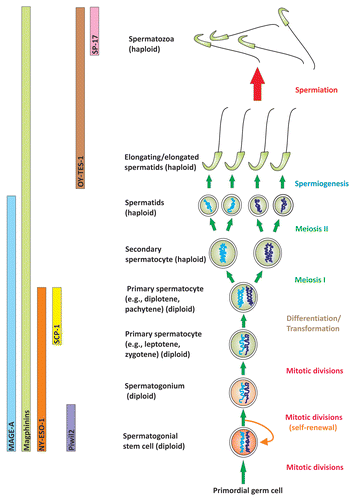
Table 1 Members of the CT antigen familyTable Footnote*
Table 2 Selected clinical trials of immunotherapy by targeting CT antigensTable Footnote*
Acknowledgments
This work was supported in part by grants from the National Institutes of Health (NICHD, U54 HD029990 Project 5 to C.Y.C.; R01 HD056034 to C.Y.C.).
References
- de Kretser D, Kerr J. Knobil E, Neill J, Ewing L, Greenwald G, Markert C, Pfaff D. The cytology of the testis. The Physiology of Reproduction 1988; 1:New York Raven Press 837 - 932
- O'Donnell L, Meachem SJ, Stanton PG, McLachlan RI. Neill JD. Endocrine regulation of spermatogenesis. Physiology of Reproduction 2006; 3:Amsterdam Elsevier 1017 - 1069
- Sharpe RM. Knobil E, Neill JD. Regulation of spermatogenesis. The Physiology of Reproduction 1994; New York Raven Press 1363 - 1434
- Bardin CW, Gunsalus GL, Cheng CY. Desjardins C, Ewing L. The cell biology of the Sertoli cell. Cell and Molecular Biology of the Testis 1993; New York Oxford University Press 189 - 219
- Griswold MD, Heckert L, Linder C. The molecular biology of the FSH receptor. J Steroid Biochem Mol Biol 1995; 53:215 - 218
- Heckert L, Griswold M. Structural organization of the follicle stimulating hormone receptor gene. Mol Endocrinol 1992; 6:70 - 80
- Bardin CW, Cheng CY, Musto NA, Gunsalus GL. Knobil E, Neill J, Ewing L, Greenwald G, Markert C, Pfaff D. The Sertoli cell. The Physiology of Reproduction 1988; New York Raven Press 933 - 974
- Hall PF. Knobil E, Neill JD, Ewing LL, Greenwald GS, Markert CL, Pfaff DW. Testicular steroid synthesis: organization and regulation. The Physiology of Reproduction 1988; 1:New York Raven Press 975 - 998
- Ge RS, Chen G, Hardy MP. The role of the Leydig cell in spermatogenic function. Adv Exp Med Biol 2008; 636:255 - 269
- Carreau S, Hess RA. Oestrogens and spermatogenesis. Philos Trans R Soc Lond B Biol Sci 2010; 365:1517 - 1535
- Carreau S, Wolczynski S, Galeraud-Denis I. Aromatase, estrogens and human male reproduction. Phil Trans R Soc Lond B Biol Sci 2010; 365:1571 - 1579
- O'Donnell L, Robertson KM, Jones ME, Simpson ER. Estrogen and spermatogenesis. Endocr Rev 2001; 22:289 - 318
- Page ST, Amory JK, Bremner WJ. Advances in male contraception. Endocr Rev 2008; 29:465 - 493
- Anderson R, Baird D. Male contraception. Endocr Rev 2002; 23:735 - 762
- Nieschlag E. The struggle for male hormonal contraception. Best Pract Res Clin Endocrinol Metab 2011; 25:369 - 375
- Liu PY, Swerdloff RS, Wang C. Recent methodological advances in male hormonal contraception. Contraception 2010; 82:471 - 475
- Cheng CY, Mruk DD. Cell junction dynamics in the testis: Sertoli-germ cell interactions and male contraceptive development. Physiol Rev 2002; 82:825 - 874
- Nieschlag E. Male hormonal contraception. Handb Exp Pharmacol 2010; 198:197 - 223
- Clermont Y. Kinetics of spermatogenesis in mammals: seminiferous epithelium cycle and spermatogonial renewal. Physiol Rev 1972; 52:198 - 235
- Parvinen M. Regulation of the seminiferous epithelium. Endocr Rev 1982; 3:404 - 417
- Hess RA, de Franca LR. Spermatogenesis and cycle of the seminiferous epithelium. Adv Exp Med Biol 2008; 636:1 - 15
- Winters SJ, Moore JP. Paracrine control of gonadotrophs. Semin Reprod Med 2007; 25:379 - 387
- Franca LR, Ogawa T, Avarbock MR, Brinster RL, Russell LD. Germ cell genotype controls cell cycle during spermatogenesis in the rat. Biol Reprod 1998; 59:1371 - 1377
- Atiken RJ, Baker MA, Doncel GF, Matzuk MM, Mauck CK, Harper MJ. As the world grows: contraception in the 21st century. J Clin Invest 2008; 118:1330 - 1343
- Cheng CY, Mruk DD. New frontiers in non-hormonal male contraception. Contraception 2010; 82:476 - 482
- Mruk DD, Cheng CY. Delivering non-hormonal contraceptives to men: advances and obstacles. Trends Biotechnol 2008; 26:90 - 99
- Mruk DD. New perspectives in non-hormonal male contraception. Trends Endocrinol Metab 2008; 19:57 - 64
- Cheng CY, Mruk DD. The biology of spermatogenesis: the past, present and future. Philos Trans R Soc Lond B Biol Sci 2010; 365:1459 - 1463
- O'Donnell L, Nicholls PK, O'Bryan MK, McLachlan RI, Stanton PG. Spermiation: the process of sperm release. Spermatogenesis 2011; 1:14 - 35
- Dym M. Basement membrane regulation of Sertoli cells. Endocr Rev 1994; 15:102 - 115
- Siu MKY, Cheng CY. Dynamic cross-talk between cells and the extracellular matrix in the testis. BioEssays 2004; 26:978 - 992
- Wong EWP, Mruk DD, Cheng CY. Biology and regulation of ectoplasmic specialization, an atypical adherens junction type, in the testis. Biochem Biophys Acta 2008; 1778:692 - 708
- Yan HHN, Mruk DD, Lee WM, Cheng CY. Ectoplasmic specialization: a friend or a foe of spermatogenesis?. BioEssays 2007; 29:36 - 48
- Lie PPY, Cheng CY, Mruk DD. The biology of the desmosome-like junction: A versatile anchoring junction and signal transducer in the seminiferous epithelium. Int Rev Cell Mol Biol 2011; 286:223 - 269
- Cheng CY, Mruk DD. The blood-testis barrier and its implication in male contraception. Pharmacol Rev 2011; In press http://dx.doi.org/10.1124/pr110002790
- Jegou B, Cudicini C, Gomez E, Stephan J. Interleukin-1, interleukin-6 and the germ cell-Sertoli cell cross-talk. Reprod Fertil Dev 1995; 7:723 - 730
- Jegou B, Pineau C. Current aspects of autocrine and paracrine regulation of spermatogenesis. Adv Exp Med Biol 1995; 377:67 - 86
- Mruk DD, Cheng CY. Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocr Rev 2004; 25:747 - 806
- Dufour JM, Dass B, Halley KR, Korbutt GS, Dixon DE, Rajotte RV. Sertoli cell line lacks the immunoprotective properties associated with primary Sertoli cells. Cell Transplant 2008; 17:525 - 534
- Fijak M, Bhushan S, Meinhardt A. Immunoprivileged sites: the testis. Methods Mol Biol 2011; 677:459 - 470
- Meinhardt A, Hedger MP. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol Cell Endocrinol 2011; 335:60 - 68
- Berndtson WE, Thompson TL. Changing relationships between testis size, Sertoli cell number and spermatogenesis in Sprague-Dawley rats. J Androl 1990; 11:429 - 435
- Auharek SA, Franca LR, McKinnell C, Jobling MS, Scott HM, Sharpe RM. Prenatal plus postnatal exposure to Di(n-Buttyl) phthalate and/or flutamide markedly reduces final Sertoli cell number in the rat. Endocrinology 2010; 151:2868 - 2875
- Atanassova NN, Walker M, McKinnell C, Fisher JS, Sharpe RM. Evidence that androgens and oestrogens, as well as follicle-stimulating hormone, can alter Sertoli cell number in the neonatal rat. J Endocr 2005; 184:107 - 117
- Orth JM. Proliferation of Sertoli cells in fetal and post-natal rats: A quantitative autoradiographic study. Anat Rec 1982; 203:485 - 492
- Chui K, Trivedi A, Cheng CY, Cherbavaz DB, Dazin PF, Huynh ALT, et al. Characterization and functionality of proliferative human Sertoli cells. Cell Transplant 2011; 20:619 - 635
- Ahmed EA, Barten-van Rijbroek AD, Kal HB, Sadri-Ardekani H, Mizrak SC, van Pelt AM, et al. Proliferative activity in vitro and DNA repair indicate that adult mouse and human Sertoli cells are not terminally differentiated, quiescent cells. Biol Reprod 2009; 80:1084 - 1091
- Fratta E, Coral S, Covre A, Parisi G, Colizzi F, Danielli R, et al. The biology of cancer testis antigens: putative function, regulation and therapeutic potential. Mol Oncol 2011; 5:164 - 182
- Akers SN, Odunsi K, Karpf AR. Regulation of cancer germline antigen gene expression: implications for cancer immunotherapy. Future Oncol 2010; 6:717 - 732
- Chen YT, Scanlan MJ, Sahin U, Tureci O, Gure AO, Tsang S, et al. A testicular antigen aberrantly expressed in human cancers detected by autologous antibody screening. Proc Natl Acad Sci USA 1997; 94:1914 - 1918
- Chen YT, Stockert E, Chen Y, Garin-Chesa P, Rettig WJ, van der Bruggen P, et al. Identification of the MAGE-1 gene product by monoclonal and polyclonal antibodies. Proc Natl Acad Sci USA 1994; 91:1004 - 1008
- Jungbluth AA, Busam KJ, Kolb D, Iversen K, Coplan K, Chen YT, et al. Expression of MAGE-antigens in normal tissus and cancer. Int J Cancer 2000; 85:460 - 465
- Boel P, Wildmann C, Sensi Ml, Brasseur R, Renauld J, Coulie P, et al. BAGE: a new gene encoding an antigen recognized on human melanomas by cytolytic lymphocytes. Immunity 1995; 2:167 - 175
- van der Bruggen P, Traversari C, Chomez P, Lurquin C, De Plaen E, Van den Eynde B, et al. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science 1991; 254:1643 - 1647
- Van den Eynde B, Peeters O, De Backer O, Gaugler B, Lucas S, Boon T. A new family of genes coding for an antigen recognized by autologous cytolytic T lymphocytes on a human melanoma. J Exp Med 1995; 182:689 - 698
- Gaugler B, Van den Eynde B, van der Bruggen P, Romero P, Gaforio JJ, de Plaen E, et al. Human gene MAGE-3 codes for an antigen recognizedon a melanoma by autologous cytolytic T lymphocytes. J Exp Med 1994; 179:921 - 930
- Chen YT, Boyer AD, Viars CS, Tsang S, Old LJ, Arden KC. Genomic cloning and localization of CTAG, a gene encoding an autoimmunogenic cancer-tesis antigen NY-ESO-1, to human chromosome Xq28. Cytogenet Cell Genet 1997; 79:237 - 240
- Scanlan MJ, Gure AO, Jungbluth AA, Old LJ, Chen YT. Cancer/testis antigens: an expanding family of targets for cancer immunotherapy. Immunol Rev 2002; 188:22 - 32
- Simpson AJG, Caballero OL, Jungbluth A, Chen YT, Old LJ. Cancer/testis antigens, gametogenesis and cancer. Nat Rev Cancer 2005; 5:615 - 625
- Mirandola L, Cannon MJ, Cobos E, Bernardini G, Jenkins MR, Kast WM, et al. Cancer testis antigens: novel biomarkers and targetable proteins for ovarian cancer. Int Rev Immunol 2011; 30:127 - 137
- Cebon J. Cancer vaccines: Where are we going?. Asia Pac J Clin Oncol 2010; 6:9 - 15
- Akers SN, Odunsi K, Karpf AR. Regulation of cancer germline antigen gene expression: implications for cnacer immunotherapy. Futre Oncol 2010; 6:717 - 732
- Caballero OL, Chen YT. Cancer/testis (CT) antigens: Potential targets for immunotherapy. Cancer Sci 2009; 100:2014 - 2021
- Song MH, Ha JC, Lee SM, Park YM, Lee SY. Identification of BCP-20 (FBXO39) as a cancer/testis antigen from colon cancer patients by SEREX. Biochem Biophys Res Commun 2011; 408:195 - 201
- Chen YT, Chadbum A, Lee P, Hsu MR E, Chiu A, Gnjatic S, et al. Expression of cancer testis antigen CT45 in classical Hodgkin lymphoma and other B-cell lymphomas. Proc Natl Acad Sci USA 2010; 107:3093 - 3098
- Zendman AJ, Ruiter DJ, van Muijen GN. Cancer/testis-associated genes: identification, experssion profile and putative function. J Cell Physiol 2003; 194:272 - 288
- Caballero OL, Zhao Q, Rimoldi D, Stevenson BJ, Svobodova S, Devalle S, et al. Frequent MAGE mutations in human melanoma. PLoS One 2010; 5:12773
- Chen YT, Ross DS, Chiu R, Zhou XK, Chen YY, Lee P, et al. Multiple cancer/testis antigens are preferentially expressed in hormone-receptor negative and high-grade brease cancers. PLoS One 2011; 6:17876
- Boss DS, Glen H, Beijnen JH, de Jong D, Wanders J, Evans TR, et al. Serum β-HCG and CA-125 as tumor markers in a patient with osteosarcoma: case report. Tumori 2011; 97:109 - 114
- Silina K, Zayakin P, Kalnina Z, Ivanova L, Meistere I, Endzelins E, et al. Sperm-associated antigens as targets for cancer immunotheraphy: expression pattern and humoral immune response in cancer patients. J Immunother 2011; 34:28 - 44
- Maruyama M, Yoshitake HTH, Takamori K, Araki Y. Molecular expression of Ly6k, a putative glycosylphosphatidyl-inositol-anchored membrane protein on the mouse testicular germ cells. Biochem Biophys Res Commun 2010; 402:75 - 81
- Matsuda R, Enokida H, Chiyomaru T, Kikkawa N, Sugimoto T, Kawakami K, et al. LY6K is a novel molecular target in blader cancer on basis of integrate genome-wide profiling. Br J Cancer 2011; 104:376 - 386
- Li F, Liu Q, Han Y, Wu B, Yin H. Sperm protein 17 is highly expressed in endometrial and cervical cancers. BMC Cancer 2010; 10:429
- Demento SL, Siefert AL, Bandyopadhyay A, Sharp FA, Fahmy TM. Pathogen-associated molecular patterns on biomaterials: a paradigm for engineering new vaccines. Trends Biotechnol 2011; 29:294 - 306
- Kawada J, Wada H, Isobe M, Gnjatic S, Nishikawa H, Jungbluth AA, et al. Heteroclitic serological response in esophageal and prostate cancer patients after NY-ESO-1 protein vaccination. Int J Cancer 2011; In press http://dx.doi.org/10.1002/ijc26074
- Kong M, Richardson RT, Widgren EE, O'Rand MG. Sequence and localization of the mouse sperm autoantigenic proteins, Sp17. Biol Reprod 1995; 53:579 - 590
- Lea Ia, Richardson RT, Widgren EE, O'Rand MG. Cloning and sequencing of cDNAs encoding the human sperm protein, Sp17. Biochem Biophys Acta 1996; 1307:263 - 266
- Chiriva-Internati M, Gagliano N, Donetti E, Costa F, Grizzi F, Franceschini B, et al. Sperm protein 17 is expressed in the sperm fibrous sheath. J Transl Med 2009; 7:61
- Luconi M, Baldi E. How do sperm swim? Molecular mechanisms underlying sperm motility. Cell Mol Biol 2003; 49:357 - 369
- Vijayaraghavan S, Liberty GA, Mohan J, Winfrey VP, Olson GE, Carr DW. Isolation and molecular characterization of AKAP110, a novel, sperm-specific protein kinase A-anchoring protein. Mol Endocrinol 1999; 13:705 - 717
- Wen Y, Richardson RT, O'Rand MG. Processing of the sperm protein Sp17 during the acrosome reaction and characterization as a calmodulin binding protein. Dev Biol 1999; 206:113 - 122
- Lea IA, Widgren EE, O'Rand MG. Association of sperm protein 17 with A-kinase anchoring protein 3 in flagella. Reprod Biol Endocrinol 2004; 2:57
- Wen Y, Richardson RT, Widgren EE, O'Rand MG. Characterization of Sp17: a ubiquitous three domain protein that binds heparin. Biochem J 2001; 357:25 - 31
- McLeskey SB, Dowds C, Carballada R, White RR, Saling PM. Molecules involved in mammalian sperm-egg interaction. Int Rev Cytol 1997; 177
- Primakoff P, Myles DG. Penetration, adhesion and fusion in mammalian sperm-egg interaction. Science 2002; 296:2183 - 2185
- Lacy M, Sanderson R. Sperm protein Sp17 is expressed on normal and malignant lymphocyts and promotes heparan sulfate-mediated cell-cell adhesion. Blood 2001; 98:2160 - 2165
- Frayne J, Hall L. A re-evaluation of sperm protein 17 (Sp17) indicates a regulatory role in an A-kinase anchoring protein complex, rather than a unique role in sperm-zona pellucida binding. Reproduction 2002; 124:767 - 774
- Dong G, Loukinova E, Smith CW, Chen Z, Van Waes C. Genes differentially expressed with malignant transformation and metastatic tumor progression of murine squamous cell carcinoma. J Cell Biochem 1997; 28:90 - 100
- Lim SH, Wang ZQ, Chiriva-Internati M, Xue Y. Sperm protein 17 is a novel cancer-testis antigen in multiple myeloma. Blood 2001; 978:1508 - 1510
- Straugh JMJ, Shaw DR, Guerrero A, Bhoola SM, Racelis A, Wang ZQ, et al. Expression of sperm protein 17 (Sp17) in ovarian cancer. Int J Cancer 2004; 108:805 - 811
- De Jong ABR, Robbins D. Characterization of sperm protein 17 in human somatic and neoplastic tissue. Cancer Lett 2002; 186:201 - 209
- Grizzi F, Gaetani P, Franceschini B, Di leva A, Colombo P, Ceva-Grimaldi G, et al. Sperm protein 17 is expressed in human nverous system tumours. MBC Cancer 2006; 6:23 - 29
- Straughn JMJ, Shaw DR, Guerrero A, Bhoola SM, Racelis A, Wang ZQ, et al. Expression of sperm protein 17 (Sp17) in ovarian cancer. Int J Cancer 2004; 108:805 - 811
- Gupta G, Sharma R, Chattopadhyay TK, Gupta SD, Ralhan R. Clinical significance of sperm protein 17 expression and immunogenicity in esophageal cancer. Int J Cancer 2007; 120:1739 - 1747
- Chiriva-Internati M, Weidanz JA, Yu Y, Frezza EE, Jenkins MR, Kennedy RC, et al. Sperm protein 17 is a suitable target for adoptive T-cell-based immunotheraphy in human ovarian cancer. J Immunother 2008; 31:693 - 703
- Grizzi F, Chiriva-Internati M, Fraceschini B, Bumm K, Colombo P, Ciccarelli M, et al. Sperm protein 17 is expressed in human somatic ciliated epithelia. J Histochem Cytochem 2004; 52:549 - 554
- Takeoka Y, Kenny TP, yago H, Naiki M, Gershwin ME, Robbins DL. Developmental considerations of sperm protein 17 gene expression in rheumatoid arthritis synoviocytes. Dev Immunol 2002; 9:97 - 102
- Franceschini B, Grizzi F, Colombo P, Soda G, Bumm K, Hermonat PL, et al. Experssion of human sperm protein 17 in melanophages of cutaneous melanocytic lesions. Br J Dermatol 2004; 150:780 - 782
- Chiriva-Internati M, Yu Y, Mirandola L, Jenkins MR, Chapman C, Cannon M, et al. Cancer testis antigen vaccination affords long-term protection in a murine modle of ovarian cancer. PLoS One 2010; 5:10471
- Lethe B, Lucas S, Michaux L, De Smet C, Godelaine D, Serrano A, et al. LAGE-1, a new gene with tumor specificity. Int J Cancer 1998; 76:903 - 908
- Jungbluth AA, Chen YT, Stockert E, Busam KJ, Kolb D, Iversen K, et al. Immunohistochemical analsyis of NY-ESO-1 antigen expression in normal and malignant human tissues. Int J Cancer 2001; 92:856 - 860
- Satie AP, Rajpert-De Meyts E, Spagnoli GC, Henno S, Olivo L, Jacobsen GK, et al. The cancer-testis gene, NY-ESO-1, is expressed in normal fetal and adult testes and in spermatocytic seminomas and testicular carcinoma in situ. Lab Invest 2002; 82:775 - 780
- Wang Y, Wu XJ, Zhao AL, Yuan YH, Chen YT, Jungbluth AA, et al. Cancer/tesis antigen expression and autologous humoral immunity to NY-ESO-1 in gastric cancer. Cancer Immunol 2004; 4:11
- Karbach J, Gnjatic S, Bender A, Neumann A, Weidmann E, Yuan J, et al. Tumor-reactive CD8+ T-cell responses after vaccination with NY-ESO-1 peptide, CpG7909 and Montanide ISA-51: association with survival. Int J Cancer 2010; 126:909 - 918
- Gnjatic S, Nishikawa H, Jungbluth AA, Gure AO, Ritter G, Jager E, et al. NY-ESO-1: review of an immunogenic tumor antigen. Adv Cancer Res 2006; 95:1 - 30
- Nicholaou T, Ebert LM, Davis ID, McArthur GA, Jackson H, Dimopoulos N, et al. Regulatory T-cell-mediated attenuation of T-cell responses to the NY-ESO-1 ISCOMATRIX vaccine in patients with advanced malignant melanoma. Clin Cancer Res 2009; 15:2166 - 2173
- Tarhini AA, Iqbal F. CTLA-4 blockade: therapeutic potential in cancer treatments. Onco Targets Ther 2010; 3:15 - 25
- Yuan J, Gnjatic S, Li H, Powel S, Gallardo HF, Ritter E, et al. CTLA-4 blockade enhances polyfunctional NY-ESO-1 specific T cell responses in metastatic melanoma patients with clinical benefit. Proc Natl Acad Sci USA 2008; 105:20410 - 20415
- van der Bruggen P, Traversari C, Chomez P, Lurguin C, De Plaen E, Van den Eynde B, et al. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science 1991; 254:1643 - 1647
- Chomez P, De Backer O, Bertrand M, De Plaen E, Boon T, Lucas S. An overview of the MAGE gene family with the identification of all human members of the family. Cancer Res 2001; 61:5544 - 5551
- Sang M, Wang L, Ding C, Zhou X, Wang B, Wang L, et al. Melanoma-associated antigen genes—an update. Cancer Letts 2011; 302:85 - 90
- Zandman AJ, de Wit NJ, van Kraats AA, Weidle UH, Ruiter DJ, van Muijen GN. Expression profile of genes coding for melanoma differentiation antigens and cancer/testis antigens in metastatic lesions of human cutaneous melanoma. Res 2001; 11:451 - 459
- Osterlund C, Tohonen V, Forslund KO, Nordqvist K. Mage-b4, a novel melanoma antigen (MAGE) gene specifically expressed during germ cell differentiation. Cancer Res 2000; 60:1054 - 1061
- Morishima N, Namanishi K, Takenouchi H, Shibata T, Tasuhiko Y. An endoplasmic reticulum stress-specific caspase cascade in apoptosis. Cytochrome c-independent activation of caspase-9 by caspase-12. J Biol Chem 2002; 277:34287 - 34294
- Borden KL, Freemont PS. The RING finger domain: a recent example of a sequence-structure family. Curr Opin Struct Biol 1996; 6:395 - 401
- Jin X, Cheng HC, Chen J, Zhu D. RNF13: an emerging RING finger ubiquitin ligase important in cell proliferation. FEBS J 2011; 278:78 - 84
- Eng Y, Gao J, Yang M. When MAGE meets RING: insights into biological functions of MAGE proteins. Protein Cell 2011; 2:7 - 12
- Tyagi P, Mirakhur B. MAGRIT: the largest-ever phase III lung cancer trial aims to establish a novel tumor-specific approach to theraphy. Clin Lung Cancer 2009; 10:371 - 374
- Trerfzer U, Jouary T, Robert C, Santinami M, Levchenko E, Smithers B, et al. DERMA Phase III trial of MAGE-A3 immunotherapy as adjuvant treatment in stage III melanoma: MAGE-A3 gene expression frequency and baseline demographics. Melanoma Res 2010; 20:34 - 35
- Wang X, Gao X, Liu L, Xu Y. MAGED1: molecular insights and clinical implications. Ann Med 2011; 43:347 - 355
- Barker PA, Salehi A. The MAGE proteins: emerging roles in cell cycle progression, apoptosis and neurogenetic disease. J Neurosci Res 2002; 67:705 - 712
- Harbour JW, Dean DC. The Rb/E2F pathway: expanding roles and emerging paradigms. Genes Dev 2000; 14:2393 - 2409
- Taniura H, Taniguchi N, Hara M, Yoshikawa K. Necdin, a postmitotic neuron-specific growth suppressor, interacts with viral transforming proteins and cellular transcription factor E2F1. J Biol Chem 1998; 273:720 - 728
- Chapman EJ, Knowles MA. Necdin: a multi functional protein withpotential tumor suppressor role?. Mol Carcinog 2009; 48:975 - 981
- Ingraham CA, Wertalik L, Schor NF. Necdin and neurotrophin receptors: interactors of relevance for neuronal resistance to oxidant stress. Pediatr Res 2011; 69:279 - 284
- Naka N, Takenaka S, Araki N, Miwa T, Hashimoto N, Yoshioka K, et al. Synovial sarcoma is a stem cell malignancy. Stem Cells 2010; 28:1119 - 1131
- dos Santos NR, de Bruijn DR, van Kessel AG. Molecular mechanisms underlying human synovial sarcoma development. Genes Chromosomes Cancer 2001; 30:1 - 14
- de Bruijn DR, Nap JP, Van Kessel AG. The (epi)genetics of human synovial sarcoma. Genes Chromosomes Cancer 2007; 46:107 - 117
- Ishida M, Miyamoto M, Naitoh S, Tatsuda D, Hasegawa T, Nemoto T, et al. The SYT-SSX fusion protein downregulates the cell proliferation regulator COM1 in t(x;18) synovial sarcoma. Mol Cell Biol 2007; 27:1348 - 1355
- Gure AO, Wei JJ, Old LJ, Chen YT. The SSX gene family: characterization of 9 complete genes. Int J Cancer 2002; 101:448 - 453
- Gure AO, Tureci O, Sahin U, Tsang S, Scanlan MJ, Jager E, et al. SSX: a multigene family with severael members transcribed in normal tesis and human cancer. Int J Cancer 1997; 72:965 - 971
- Jager D, Stockert E, Scanlan MJ, Gure AO, Jager E, Knuth A, et al. Cancer-testis antigens and ING1 tumor suppressor gene production are breast cancer antigens: characterization of tissue-specific ING1 transcripts and a homologue gene. Cancer Res 1999; 59:6197 - 6204
- Michel JJ, Scott JD. AKAP mediated signal transduction. Annu Rev Pharmacol Toxicol 2002; 42:235 - 257
- Smith HA, McNeel DG. The SSX family of cancer-testis antigens as target proteins for tumor therapy. Clin Dev Immunol 2010; 2010:150591
- Bodey B. Cancer-testis antigens: promising targets for antigen directed antineoplastic immunotherapy. Expert Opin Biol Ther 2002; 2:577 - 584
- Dos Santos NR, Torensma R, de Vries TJ, Schreurs MWJ, de Bruijn DRH, Kater-Baats E, et al. Heterogeneous expression of the SSX cancer/testis antigens in human melanoma lesions and cell lines. Cancer Res 2000; 60:1654 - 1662
- Meuwissen RLJ, Offenberg HH, Dietrich AJJ, Riesewijk A, van Iersel M, Heyting C. A coiled-coil related specific for synapsed regions of meiotic prophase chromosomes. EMBO J 1992; 11:5091 - 5100
- Tureci O, Sahin U, Zwick C, Koslowski M, Seitz G, Pfreundschuh M. Identification of a meiosis-specific protein as a member of the class of cancer/testis antigens. Proc Natl Acad Sci USA 1998; 95:5211 - 5216
- Tammela J, Jungbluth AA, Qian F, Santiago D, Scanlan MJ, Keitz B, et al. SCP-1 cancer/testis antigen is a prognostic indicator and a candidate target for immunotherapy in epithelial ovarian cancer. Cancer Immun 2004; 4:1 - 10
- Cannistra SA. Cancer of the ovary. N Engl J Med 2004; 351:2519 - 2529
- Saito K, Nishida KM, Mori T, Kawamura Y, Miyoshi K, Nagami T, et al. Specific association of Piwi with rasiRNAs derived from retrotransposon and hetero-chromatic regions in the Drosophila genome. Genes Dev 2006; 20:2214 - 2222
- Katahira J, Yoneda Y. Nucleocytoplasmic transport of microRNAs and related small RNAs. Traffic 2011; In press http://dx.doi.org/10.1111/j1600-0854201101211x
- Cox DN, Chao A, Lin HY. piwi encodes a nucleoplasmic factor whose activity modulates the number and division rate of germline stem cells. Development 2000; 127:503 - 514
- Thomson T, Lin H. The biogenesis and function of PIWI proteins and piRNAs: progress and prospect. Annu Rev Cell Dev Biol 2009; 25:355 - 376
- Saxe JP, Lin H. Small noncoding RNAs in the germline. Cold Spring Harb Perspect Biol 2011; In press http://dx.doi.org/10.1101/cshperspecta002717
- Cox DN, Chao A, Baker J, Chang L, Qiao D, Lin H. A novel class of evolutionarily conserved genes defined by piwi aer essential for stem cell self-renewal. Genes Dev 1998; 12:3715 - 3727
- Cenik ES, Zamore PD. Argonaute proteins. Curr Biol 2011; 21:446 - 449
- Ghildiya lM, Zamore PD. Small silencing RNAs: an expanding universe. Nat Rev Genet 2009; 10:94 - 108
- Li L, Liu Y. Diverse small non-coding RNAs in RNA interference pathways. Methods Mol Biol 2011; 764:169 - 182
- Czech B, Hannon GJ. Small RNA sorting: matchmaking for Argonautes. Nat Rev Genet 2011; 12:19 - 31
- Saito K, Siomi MC. Small RNA-mediated quiescence of transposable elements in animals. Dev Cell 2010; 19:687 - 697
- Deng W. Lin H. miwi, a murine homolog of piwi, encodes a cytoplasmic protein essential for spermatogenesis. Dev Cell 2002; 2:819 - 830
- Cerutti L, Mian N, Bateman A. Domains in gene silencing and cell differentiation proteins: the novel PAZ domain and redefinition of the Piwi domain. Trends Biochem Sci 2000; 25:481 - 482
- Lee JH, Schutte D, Wulf G, Fuzesi L, Radzun HJ, Schweyer S, et al. Stem-cell protein Piwil2 is widely expressed in tumors and inhibits apoptosis through activation of Stat3/Bcl-XL pathway. Hum Mol Genet 2006; 15:201 - 211
- Wang QE, Han C, Milum K, Wani AA. Stem cell protein Piwil2 modulates chromatin modifications upon cisplatin treatment. Mutat Res 2011; 708:59 - 68
- Ye Y, Yin DTCL, Zhou Q, Shen R, He G, Yan Q, et al. Identification of Piwil2-like (PL2L) proteins that promote tumorigenesis. PLoS One 2010; 5:13406
- Ono T, Kurashige T, Harada N, Noguchi Y, Saika T, Niikawa N, et al. Identification of proacrosin binding protein sp32 precursor as a human cern/testis antigen. Proc Natl Acad Sci USA 2001; 98:3282 - 3287
- Baba T, Azuman S, Kashiwabara S, Toyoda Y. Sperm from mice carrying a targeted mutation of the acrosin gene can penetrate the oocyte zona pellucida and effect fertilization. J Biol Chem 1994; 269:31845 - 31849
- Whitehurst AW, Xie Y, Purinton SC, Cappell KM, Swanik JT, Larson B, et al. Tumor antigen acrosin binding protein normalizes mitotic spindle function to promote cancer cell proliferation. Cancer Res 2010; 70:7652 - 7661
- Tammela J, Uenaka A, Ono T, Noguchi Y, Jungbluth AA, Mhawech-Fauceglia PQF, et al. OY-TES-1 expression and serum immunoreactivity in epithelial ovarian cancer. Int J Oncol 2006; 29:903 - 910
- Rimoldi D, Salvi S, Schultz TE, Spagnoli GC, Cerottini JC. Anti-MAGE-3 antibody 57B and anti-MAGE-1 antibody 6C1 can be used to study different proteins of the MAGE-A family. Int J Cancer 2000; 86:749 - 751
- Jungbluth AA, Stockert E, Chen YT, Kolb D, Iversen K, Coplan K, et al. Monoclonal antibody MA454 reveals a heterogeneous expression pattern of MAGE-1 antigen in formalin-fixed paraffinembedded lung tumours. Br J Cancer 2000; 83:493 - 497
- Saburi S, Nadano D, Akama TO, Hirama K, Yamanouchi K, Naito K, et al. The trophinin gene encodes a novel group of MAGE proteins, magphinins and regulates cell proliferation during gametogenesis in the mouse. J Biol Chem 2001; 276:49378 - 49389
- Hofmann MC, Narisawa S, Hess RA, Millan JL. Immortalization of germ cells and somatic testicular cells using the SV40 large T antigen. Exp Cell Res 1992; 201:417 - 435
- Fogh J, Fogh Jm, Orfeo T. One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice. J Natl Cacer Inst 1977; 59:221 - 226