Abstract
Zonula occludens proteins (ZO-1, ZO-2, ZO-3), which belong to the family of membrane-associated guanylate kinase (MAGUK) homologs, serve as molecular hubs for the assembly of multi-protein networks at the cytoplasmic surface of intercellular contacts in epithelial and endothelial cells. These multi-PDZ proteins exert crucial functions in the structural organization of intercellular contacts and in transducing intracellular signals from the plasma membrane to the nucleus. The junctional MAGUK protein ZO-2 not only associates with the C-terminal PDZ-binding motif of various transmembrane junctional proteins but also transiently targets to the nucleus and interacts with a number of nuclear proteins, thereby modulating gene expression and cell proliferation. Recent evidence suggests that ZO-2 is also involved in stress response and cytoprotective mechanisms, which further highlights the multi-faceted nature of this PDZ domain-containing protein.
This review focuses on ZO-2 acting as a molecular scaffold at the cytoplasmic aspect of tight junctions and within the nucleus and discusses additional aspects of its cellular activities. The multitude of proteins interacting with ZO-2 and the heterogeneity of proteins either influencing or being influenced by ZO-2 suggests an exceptional functional capacity of this protein far beyond merely serving as a structural component of cellular junctions.
Introduction
Direct interactions of neighboring cells are conferred by several types of cellular junctions. These can functionally be grouped into occluding (i.e., tight junctions), anchoring (i.e., adherens junctions, desmosomes and hemidesmosomes) and communicating junctions (i.e., gap junctions). Tight junctions (TJs) are characterized by dynamic and highly organized plasma membrane-bound structures, which seal the paracellular space in order to allow the establishment and maintenance of a distinct internal and external milieu in tissues and organs.Citation1-Citation3 The formation of such “ins” and “outs” is imperative for proper development and tissue function. For instance, the tight intercellular bonding between trophectoderm epithelial cells is necessary to create a suitable internal milieu for the developing mammalian embryo.Citation4 Or, the restrictive intercellular contacts between capillary endothelial cells, forming the so called blood-brain barrier, enable a physiological functioning of the brain within a homeostatic environment and protect the vulnerable neural tissue from the uncontrolled entrance of substances emanating from the systemic circulation.Citation5-Citation8 Next to regulating the paracellular flux of solutes and ions, tight junctions also create a “fence” in the plane of the plasma membrane, contributing to the establishment of cellular polarity.Citation9-Citation12 Epithelial and endothelial tight junctions consist of several classes of transmembrane proteins locating to the apical aspect of the cell.Citation13 The extracellular portions of the transmembrane proteins form a more or less tight seal between opposing cells, connecting the plasma membranes in a zipper-like fashion. Within the cell a highly organized cytoplasmic “plaque” consisting of an array of cytosolic proteins assembles in close vicinity to these contact sites, providing mechanical linkage between the plasma membrane and the cytoskeleton. A large number of junctional proteins have been identified during the last decade (reviewed inCitation13-Citation19) and soon it was realized that these complexes not only ensure the structural integrity of tight junctions,Citation20-Citation23 but also coordinate and transduce the signals impinging on and emanating from the apical plasma membrane. Consequently, next to serving as physical barriers tight junctions also influence pivotal processes such as morphogenesis, gene expression, cytoskeletal dynamics and cell proliferation.Citation14,Citation18,Citation24-Citation26
PDZ-Containing Scaffolding Proteins—The Molecular Basis for Junctional Plaque Assembly
The most prominent subgroup of scaffolding proteins localizing to the cytoplasmic face of the adhesion site is represented by the MAGUK (membrane-associated guanylate kinase) proteins,Citation27-Citation29 which can be classified into seven subfamilies based on their modular structure and sequence similarity.Citation30 The modular nature of MAGUK proteins was early recognized and comprises one or more PDZ domains next to other protein-protein binding domains including SH3 (Src homology 3), WW, L27 (MAGUK LIN-2 + LIN-7), CaMK (calcium/calmodulin-dependent protein kinase) domains and a GUK (guanylate kinase) domain,Citation28 a region which lacks enzymatic activity due to a deletion in its ATP- and GMP-binding sites but appears to enable protein–protein binding.Citation31 The most intensely studied junctional MAGUKs include the zonula occludens proteins (ZO-1, ZO-2 and ZO-3).Citation32-Citation35 Although originally described as TJ-specific proteins, ZO proteins also associate with other types of intercellular contacts, i.e., adherens and gap junctions (reviewed inCitation13,Citation14,Citation36,Citation37). Thus, ZO proteins appear to exert, at least in part, a basic and redundant role in establishing and/or maintaining intercellular adhesion and communication. Generally, ZO proteins serve as a multivalent binding platform for an ever growing list of junction-associated proteins. Next to the classical MAGUK domains, ZO proteins also contain a short binding region at the C-terminus (TEL), which represents an additional PDZ binding domain. Finally, a proline-rich region of varying length is present at the C-terminus of ZO-1 and ZO-2 and between the second and third PDZ domain of ZO-3 and has been shown to be targeted by actin-binding proteins and other cytoskeletal proteins.Citation38-Citation40 In recent years, increasing interest has focused on the role of variable regions, numbered U (unique)1 to U6, which are localized between the core domains listed further above. The best studied variable region, termed HOOK domain (U5), is a basic hinge region between the SH3 and GUK domain which is involved in oligomerization and ligand binding of MAGUKs.Citation41 The U6 motif is unique to ZO proteins and appears to be critical for junction assembly. For instance, expression of a mutant ZO-1, lacking the U6 region, induces ectopic junctional strands consisting of occludin and claudins but lacking most of the cytoplasmic plaque proteins.Citation41
Other junctional MAGUK proteins identified so far include MAGI-1, -2 and -3 (membrane-associated guanylate kinase with inverted orientation of protein–protein interaction domains-1, -2 and -3),Citation42-Citation44 and the calcium/calmodulin-dependent serine protein kinase (CASK/LIN-2).Citation45 In addition, the human homolog of the Drosophila discs large tumor suppressor hDlg/SAP97 has been shown to localize to regions of cell-cell contact.Citation46,Citation47 However the functional relevance of hDlg for TJ biology remains poorly characterized.
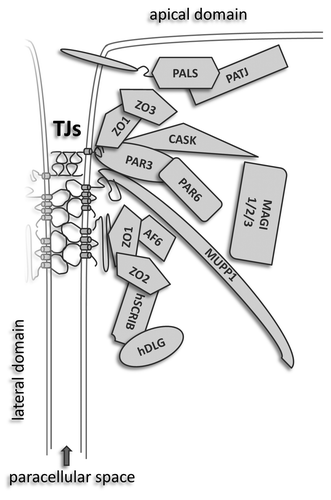
Next to the MAGUK proteins, several other proteins localizing to the junctional plaque harbor one or more PDZ domains. These protein-protein interaction modules, which often bind to short amino acid motifs at the C-termini of target proteins, are named after the first three proteins in which these domains have been identified: PSD-95 (post synaptic density protein-95), Dlg (the Drosophila discs large protein) and ZO-1 (zonula occludens-1).Citation48,Citation49 At TJs, the large 13 PDZ domain-containing protein MUPP1 (multi PDZ domain protein-1)Citation50 and ZO proteins directly bind the cytoplasmic domains of the TJ-specific transmembrane proteins occludin and claudins.Citation51-Citation53 In a similar manner, the Ig-like junction adhesion molecule 1 (JAM1) anchors Par-3,Citation54 and hence the Par6/aPKC (partitioning-defective protein-6/atypical protein kinase C) complex to the apical aspect of the plasma membrane.Citation55 PALS1 binds to a cytoplasmic portion of the transmembrane proteins CRB (crumbs)-1 and CRB-3, thereby targeting the evolutionary conserved polarity complex CRB1/3-PALS1-PATJ to the junction.Citation56 Further, the polarity protein Scribble, a large multi-domain protein involved in the maintenance of apical/basal polarity, migration and invasion,Citation57 was shown to associate with ZO-1 and has been demonstrated to be important for TJ assembly.Citation58 Finally, the Ras target AF-6/afadinCitation59 directly interacts with ZO-1 and localizes to TJs. Taken together, PDZ domain-mediated protein-protein interactions aid in the temporal and spatial assembly of multi-protein complexes at the cytoplasmic plaque and are pivotal for the functional and structural integrity of TJs (see ).
Figure 1. PDZ proteins as molecular base of tight junctional plaques. Two groups of proteins are involved in the establishment and maintenance of TJs: (1) Transmembrane proteins which bridge the intercellular space and create a paracellular seal and (2) peripheral proteins, constituting the cytoplasmic “plaque” of tight junctions interacting directly or indirectly with the transmembrane junctional components. The formation of these complexes largely relies on PDZ-PDZ interactions. Only cytosolic proteins harboring one or more PDZ domains are illustrated.
Molecular Interactions of ZO-2 at the Junctional Site
Zonula occludens 2 (ZO-2) is a 160 kDa protein originally identified in a ZO-1 immunoprecipitate.Citation32 Proteins interacting with ZO-2 at cell-cell contact sites essentially can be subdivided into structural and non-structural proteins. depicts a schematic diagram of the ZO-2 interactome. Some of the protein-protein interactions described for ZO-2 are redundant and have been shown for all ZO proteins. Such interactions include the binding of the first PDZ domain to the C-termini of claudinsCitation52 or the homo- and hetero-dimerization of ZO-proteins via their second PDZ domain, which takes place by domain swapping of β1 and β2 strands.Citation60 These redundancies are not surprising as the ZO proteins share a high degree of homology among each other. Nevertheless, ZO proteins also fulfill a multitude of ZO-specific functions.
Figure 2. The ZO-2 (TJP2) interactome. Protein-protein interactions are symbolized by bold lines whereas outgoing dashed lines indicate that perturbations of ZO-2 expression affect the expression of the indicated proteins. Incoming dashed lines refer to proteins which impact upon the mRNA level of ZO-2. The graph was generated using the IPA software package (Ingenuity Systems) and gene symbols according to the H.G.N.C. nomenclature are shown.
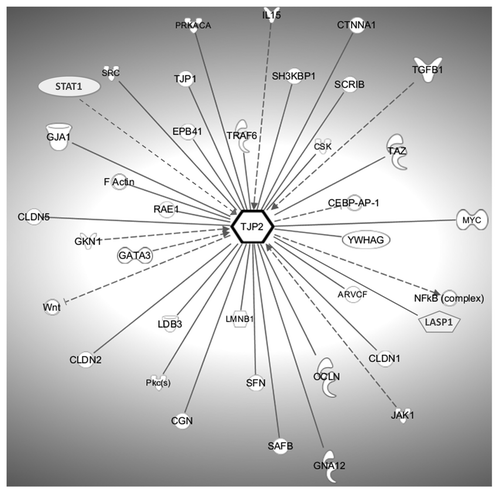
Transmembrane proteins interacting with ZO-2 include the tetra-span proteins claudins and occludin.Citation52,Citation61 Claudin-1 to -8 interact via their C-terminal region with the first PDZ domain,Citation52 while connexins (Cx), including Cx-36 and Cx-43, were found to associate with PDZ-1 or PDZ-2, respectively.Citation62,Citation63 Although the binding of occludin to a region at the SH3-hinge-GUK domain has only been demonstrated for ZO-1,Citation64 a similar interaction can be presumed for ZO-2, although this remains to be demonstrated.
The proline-rich C-terminal domain of ZO-2 has been shown to interact mainly with cytoskeleton-associated proteins, including protein 4.1R and F-actin.Citation38-Citation40 Cingulin, a 140 kDa phosphoprotein and regulator of Rho-A signaling at TJs interacts directly with ZO-2 and provides a direct link to the actomyosin cytoskeleton.Citation65,Citation66 Finally, data from a tandem proteomic approach suggests the direct or indirect interaction of ZO-2 with 14–3-3 binding proteins, which are known to be involved in cytoskeletal regulation.Citation67 ZO-2, next to ZO-1,Citation58 also associates with the basolateral polarity protein Scribble (huScribble) via a C-terminal motifCitation57 and the ZO-2/Scribble complex appears to be involved in TJ assembly and function.Citation58 Finally, protein kinases, signaling molecules and other regulatory proteins including cytokines which directly or indirectly target ZO-2 complete the dynamic network of the ZO-2 scaffold. These include various isoforms of protein kinase C,Citation68-Citation70 the tyrosine kinase c-Src and its negative regulator Csk (C-terminal Src kinase)Citation71 as well as the heterotrimeric G protein Galpha12,Citation72 which promotes Src phosphorylation of ZO-1 and ZO-2 resulting in disruption of TJs and an increase in paracellular permeability.Citation73 Interestingly, next to ZO-2 itself, several of the interacting proteins resemble “dual-residency” proteins, capable of shuttling between the cytoplasm and the nucleus. Therefore, ZO-2 serves as a molecular hub at the cytoplasmic aspect of TJs. In addition, the nuclear translocation of ZO-2 together with bound proteins substantially extends the physiological functions of this junctional MAGUK protein.
Nuclear ZO-2 and the Impact of ZO-2 on Transcription and Cell Proliferation
The presence of ZO-2 within the nucleus has been recognized more than a decade ago. All ZO proteins carry conserved nuclear localization and nuclear export sequences,Citation74 however only ZO-1 and ZO-2 have been shown to localize to cell nuclei of epithelial and endothelial cells.Citation75-Citation79 In sparse cultures, ZO-1 and ZO-2 are present in cell nuclei but become redistributed to the plasma membrane as soon as cells reach confluence.Citation76,Citation77 During mitosis and the concomitant disintegration of the nuclear membrane ZO-2 is evenly dispersed within the cytoplasm and during the late G1 phase ZO-2 accumulates in the nucleus. In quiescent cells ZO-2 is not present in the nucleus.Citation80 Further, it was demonstrated that ZO-2 preferentially targets the nuclei of epithelial cells during and upon chemical and heat stress.Citation79 Frequently, nuclear ZO-2 locates to nuclear speckles, which is dependent on the presence of an intact PDZ-1 domainCitation78 and potentially can be induced by the interaction with the heterogeneous nuclear ribonucleoprotein SAF-B/HET (scaffold attachment factor-B/HSP27 estrogen response element-TATA box-binding protein).Citation79 In addition, recruitment of ZO-2 to nuclear speckles requires binding to phosphatidylinositol 4,5-bisphosphate,Citation81 a phospholipid component of plasma membranes.
The notion that junctional MAGUKs fulfill functions next to their prime role in aiding in the establishment of cell-cell junctions is not without precedence. A first clue came from studies with the Drosophila tumor suppressor protein Dlg, a protein relevant for the formation of invertebrate septate junctions and neuronal synaptic junctions. Genetic studies in Drospohila revealed that mutations in Dlg resulted in tumorous overgrowth of imaginal discs.Citation82,Citation83 Due to their high homology with Dlg it was speculated that the ZO family of MAGUK proteins, including ZO-2, are also involved in cell growth and proliferation. Indeed, several studies have demonstrated the interaction of ZO proteins with proteins involved in cell cycle progression and transcriptional regulation.Citation24,Citation26,Citation36 However, results from studies investigating the influence of ZO-2 on cellular proliferation remain controversial. Exogenous downregulation of ZO-2 resulted in functional perturbations of TJs in epithelial cells without altering cell proliferation or apoptosis rates.Citation84 Also, during mouse blastocyst formation repression of ZO-2 delayed the formation of the blastocoel cavity, without affecting cell proliferation.Citation4 In contrast, deletion of ZO-2 in mice is embryonic lethal shortly after implantation due to an arrest in early gastrulation. Interstingly, ZO-2 knockout embryos showed a decreased proliferation rate and an increase in apoptotic cells.Citation85 Finally, the nuclear accumulation of a ZO-2 variant tagged with exogenous SV40 nuclear localization signals led to increased cell proliferation of epithelial and endothelial cells.Citation86 In another study the overexpression of ZO-2 in epithelial cells resulted in binding to c-Myc triggering the downregulation of cyclin D1 and subsequent suppression of cell proliferation.Citation87 Later, Lechuga S. et al. provided evidence that the inhibitory effect of ZO-2 on cyclin D1 transcription is blocked by ZASP (ZO-2 associated speckle protein) through binding to the third PDZ domain of ZO-2.Citation88
Modulation of cell proliferation is also facilitated by binding of ZO-2 to the transcriptional activator YAP2 (Yes kinase-associated protein 2), an effector of the Hippo signaling pathway which controls the regulation of proliferation and apoptosis during mammalian organogenesis.Citation89 Interaction with ZO-2, which can be abrogated by deletion of the 1st PDZ domain, enhances the nuclear localization and pro-apoptotic function of YAP.Citation90 Next to ZO-2, also ZO-1 has been identified as a binding partner of YAP-2, also known as TAZ (transcriptional coactivator with PDZ-binding motif).Citation91 ZO-2 was also found to interact with the cytoskeletal protein LASP-1 (LIM-and-SH3-domain-protein-1), usually present in focal contacts, where it regulates cytoskeletal dynamics and cell migration. Phosphorylation of LASP-1 by PKA (protein kinase A) induces nuclear targeting of the LASP-1/ZO-2 complexCitation92 and nuclear accumulation of LASP-1 is thought to be relevant during cancer progression.
Several proteins have been found to interact with ZO-2 during nuclear shuttling or within the nucleus itself. For example, the interaction of the armadillo-repeat protein ARVCF (armadillo repeat gene deleted in velocardiofacial syndrome) depends on an N-terminal PDZ domain of ZO-2Citation93, a region which is also targeted by the non-receptor tyrosine kinase JAK1. Also, the transcription factors Jun, Fos and C/EBP were shown to associate with ZO-2 within the nucleus and at TJs in epithelial cells.Citation94 Next to directly associating with and/or influencing transcription factors, ZO-2 possibly impacts upon the transcription machinery via its association with the hnRNP SAF-B/HET.Citation79 SAF-B/HET is thought to serve as a molecular platform to assemble a transcription complex in the vicinity of actively transcribed genes,Citation95 a mechanism which appears to be highly conserved among several species.Citation96
However, only few studies demonstrate a direct or indirect impact on gene transcription following the perturbation of ZO-2 levels. For instance, in vascular smooth muscle cells (VSMCS) of coronary arteries the expression of STAT1 (signal transducers and activators of transcription)-specific genes is upregulated in response to ZO-2 silencing.Citation97 Another study demonstrates the association of ZO-2 with the non-receptor tyrosine kinase JAK1 (Janus kinase 1) in VMSCs mediating specific homotypic intercellular contacts.Citation98 Together, these observations suggest that ZO-2 is potentially involved in vascular remodeling and arteriogenesis. Overexpression of ZO-2 was also shown to increase β-catenin phosphorylation, thereby reducing β-catenin-mediated gene transcription of Wnt target genes.Citation99 In nasal epithelial cells, the experimentally induced upregulation of ZO-2 expression through activation of PKC signaling was shown to be inhibited by the transcriptional factor GATA-3.Citation70 Finally, in a human genome-wide siRNA screen ZO-2 has been identified as a novel NFκB activation pathway component.Citation100 However, for many of these observations the physiological ramifications remain poorly defined.
Taken together, one can postulate that not only peripheral, but also nuclear ZO-2 serves as a scaffold protein facilitating the transport of other proteins to the nucleus, aiding in the spatial and temporal assembly of nuclear “transcriptomes,” thereby directly or indirectly influencing cell proliferation and/or gene transcription.
The Role of ZO-2 in Stress Response and Health and Disease
ZO-2 has also been associated with several human pathologies. Clinical studies revealed that, e.g., familial hypercholanemia or nonsyndromic progressive hearing loss are associated with mutations of the ZO-2 gene and/or a change in ZO-2 protein levels.Citation101,Citation102 Due to the high degree of homology between the Drosophila tumor suppressor protein Dlg and ZO-1, together with the actions of ZO-2 on cell proliferation and gene transcription, it is also tempting to speculate that ZO-2 could act as a tumor suppressor. Indeed, in several tumors the mRNA levels of ZO-2 are reduced (reviewed inCitation103). However, most of these reports do not exclude that the reduction of ZO-2 and/or other junctional proteins is a consequence of the disease.
Recently, Bautista-Garcia et al. demonstrated a cytoprotective function of ZO-2. In a mouse model of adriacmycin-induced kidney dysfunction overexpression of ZO-2 prevents podocyte injury by increasing phosphorylated β-catenin and decreasing the expression of Wnt/β-catenin downstream genes.Citation99 The interference of ZO-2 with the Wnt/β-catenin signaling pathway predicts an unforeseeable array of functions in biological processes where a fine-tuned balance between differentiation, cell growth and cell-cell adhesion is required. This includes the modulation of all types of epithelial-mesenchymal transition (EMT) underlying development, tissue regeneration and disease.Citation104
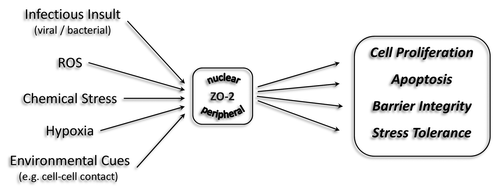
ZO-2 is also potentially involved in cellular stress responses. For instance, upon chemical stress and heat shock, ZO-2 accumulates in the nucleus of epithelial cells.Citation79 Since nuclear ZO-2 associates directly with SAF-B/HET, a negative regulator of the heat shock protein 27 (Hsp27) transcription,Citation105 one could extrapolate that ZO-2 is involved in cytoprotection by modulating Hsp27 expression levels (). Further, in vitro studies have shown that cerebral endothelial cells, stably overexpressing nuclear ZO-2, show a moderate decrease in MMP9 (matrix metalloproteinase 9) mRNA levels.Citation106 The underlying mechanism explaining the modulation of MMP9 levels by ZO-2 remains to be determined. Nevertheless, it has been demonstrated several-fold that the downregulation of MMP9 exerts a protective effect on the endothelial barrier, particularly the blood-brain barrier, following ischemic damage.Citation107-Citation111 Further, in human breast carcinoma cells MMP9 was shown to be suppressed by heme oxygenase 1 (HO-1), a target gene of the transcriptional repressor Bach1.Citation112,Citation113 Interestingly, via a yeast based Two-hybrid assay, we identified Bach1 to interact directly with ZO-2 (unpublished data). It will be of particular interest to determine whether and how this interaction modulates HO-1 levels and potentially contributes to the cytoprotective action of HO-1.
Figure 3. Functions of ZO-2 beyond cell-cell adhesion. The expression and intracellular distribution of the junctional MAGUK protein ZO-2 is influenced by various extrinsic and intrinsic cues, including cellular stress and cell-cell contact. In response ZO-2 affects diverse cellular processes, including cell proliferation, apoptosis, stress tolerance and barrier integrity by directly or indirectly influencing gene expression, ultimately feeding into various cell signaling pathways.
As mentioned before, ZO-2 has also been shown to directly or indirectly act as a negative regulator of STAT-1 expression in VSMCs.Citation95 Interestingly, the JAK/STAT pathway mediates HIV-induced adhesion and transmigration of monocytes across the blood-brain barrier in vitro through increased expression of IL-6 and IL-8.Citation114 It will be interesting to determine whether or not ZO-2 suppresses STAT-1 expression in cerebral endothelial cells as well, thereby exerting a protective effect in HIV-induced neuroinflammation by decreasing the JAK/STAT-mediated leukocyte entry into the brain.
Many TJ proteins have been shown to be directly targeted by viruses with oncogenic and non-oncogenic potential, often resulting in a perturbation of the junctional barrier of their target cells.Citation115,Citation116 As a result, the dissemination and transmission efficiency of the viruses is often increased.
Oxidative stress also alters ZO protein expression and localization in a number of pathologies, including hypoxia, bacterial and viral infections, inflammation and age-related diseases.Citation117 However, most of the studies have focused on ZO-1 and on increased permeability of epithelial and endothelial barriers and information concerning redundant and non-redundant functions of ZO proteins upon oxidative stress is still missing. Of particular interest in this context is the role of nuclear ZO-2 for the cellular stress response.
In summary, TJ-associated proteins in general and ZO-2 in particular have been associated with a multitude of physiological and pathological conditions other than forming and maintaining the tight junction permeability barrier.
Summary
Historically, the multi PDZ domain MAGUK protein ZO-2 was believed to serve as a simple structural component of the “cytoplasmic plaque” of TJs. However, the recognition that ZO-2 transiently targets the nucleus of epithelial and endothelial cells has fuelled intense interest over the years and has led to the discovery of a myriad of functions extending beyond the assembly of TJs, including the modulation of gene transcription, cell growth and proliferation. ZO-1/2 protein expression and localization also has been shown to be altered during cellular stress responses (i.e., oxidative stress or hypoxiaCitation117) and various other pathologies. More recently, ZO-2 has further been shown to exert a cytoprotective effect by perturbing the Wnt/β-catenin pathway, preventing podocyte injury in a mouse model. Much remains to be learned and additional studies dissecting the molecular mechanism(s) underlying the proposed ZO-2-mediated cytoprotective response pathways are required, providing ample opportunities for future research.
Acknowledgments
H.B. and H.C.B. are supported by the EU 7th FP (Neurobid). I.K. and I.W. are supported by OTKA K100807, PD100958 and HURO/1101/173/2.2.1
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
References
- Balda MS, Matter K. Tight junctions at a glance. J Cell Sci 2008; 121:3677 - 82; http://dx.doi.org/10.1242/jcs.023887; PMID: 18987354
- Furuse M. Molecular basis of the core structure of tight junctions. Cold Spring Harb Perspect Biol 2010; 2:a002907; http://dx.doi.org/10.1101/cshperspect.a002907; PMID: 20182608
- Luissint AC, Artus C, Glacial F, Ganeshamoorthy K, Couraud PO. Tight junctions at the blood brain barrier: physiological architecture and disease-associated dysregulation. Fluids Barriers CNS 2012; 9:23; http://dx.doi.org/10.1186/2045-8118-9-23; PMID: 23140302
- Sheth B, Nowak RL, Anderson R, Kwong WY, Papenbrock T, Fleming TP. Tight junction protein ZO-2 expression and relative function of ZO-1 and ZO-2 during mouse blastocyst formation. Exp Cell Res 2008; 314:3356 - 68; http://dx.doi.org/10.1016/j.yexcr.2008.08.021; PMID: 18817772
- Abbott NJ, Friedman A. Overview and introduction: the blood-brain barrier in health and disease. Epilepsia 2012; 53:Suppl 6 1 - 6; http://dx.doi.org/10.1111/j.1528-1167.2012.03696.x; PMID: 23134489
- Daneman R. The blood-brain barrier in health and disease. Ann Neurol 2012; 72:648 - 72; http://dx.doi.org/10.1002/ana.23648; PMID: 23280789
- Ek CJ, Dziegielewska KM, Habgood MD, Saunders NR. Barriers in the developing brain and Neurotoxicology. Neurotoxicology 2012; 33:586 - 604; http://dx.doi.org/10.1016/j.neuro.2011.12.009; PMID: 22198708
- Liu WY, Wang ZB, Zhang LC, Wei X, Li L. Tight junction in blood-brain barrier: an overview of structure, regulation, and regulator substances. CNS Neurosci Ther 2012; 18:609 - 15; http://dx.doi.org/10.1111/j.1755-5949.2012.00340.x; PMID: 22686334
- Mandel LJ, Bacallao R, Zampighi G. Uncoupling of the molecular ‘fence’ and paracellular ‘gate’ functions in epithelial tight junctions. Nature 1993; 361:552 - 5; http://dx.doi.org/10.1038/361552a0; PMID: 8429911
- Matter K, Balda MS. Functional analysis of tight junctions. Methods 2003; 30:228 - 34; http://dx.doi.org/10.1016/S1046-2023(03)00029-X; PMID: 12798137
- van Meer G, Gumbiner B, Simons K. The tight junction does not allow lipid molecules to diffuse from one epithelial cell to the next. Nature 1986; 322:639 - 41; http://dx.doi.org/10.1038/322639a0; PMID: 3748143
- van Meer G, Simons K. The function of tight junctions in maintaining differences in lipid composition between the apical and the basolateral cell surface domains of MDCK cells. EMBO J 1986; 5:1455 - 64; PMID: 3743548
- Ebnet K. Organization of multiprotein complexes at cell-cell junctions. Histochem Cell Biol 2008; 130:1 - 20; http://dx.doi.org/10.1007/s00418-008-0418-7; PMID: 18365233
- Bauer HC, Traweger A, Zweimueller-Mayer J, Lehner C, Tempfer H, Krizbai I, et al. New aspects of the molecular constituents of tissue barriers. J Neural Transm 2011; 118:7 - 21; http://dx.doi.org/10.1007/s00702-010-0484-6; PMID: 20865434
- Giepmans BN, van Ijzendoorn SC. Epithelial cell-cell junctions and plasma membrane domains. Biochim Biophys Acta 2009; 1788:820 - 31; http://dx.doi.org/10.1016/j.bbamem.2008.07.015; PMID: 18706883
- Gonzalez-Mariscal L, Nava P. Tight junctions, from tight intercellular seals to sophisticated protein complexes involved in drug delivery, pathogens interaction and cell proliferation. Adv Drug Deliv Rev 2005; 57:811 - 4; http://dx.doi.org/10.1016/j.addr.2005.01.004; PMID: 15820554
- González-Mariscal L, Tapia R, Chamorro D. Crosstalk of tight junction components with signaling pathways. Biochim Biophys Acta 2008; 1778:729 - 56; http://dx.doi.org/10.1016/j.bbamem.2007.08.018; PMID: 17950242
- Steed E, Balda MS, Matter K. Dynamics and functions of tight junctions. Trends Cell Biol 2010; 20:142 - 9; http://dx.doi.org/10.1016/j.tcb.2009.12.002; PMID: 20061152
- Wang Q, Margolis B. Apical junctional complexes and cell polarity. Kidney Int 2007; 72:1448 - 58; http://dx.doi.org/10.1038/sj.ki.5002579; PMID: 17914350
- Aijaz S, Balda MS, Matter K. Tight junctions: molecular architecture and function. Int Rev Cytol 2006; 248:261 - 98; http://dx.doi.org/10.1016/S0074-7696(06)48005-0; PMID: 16487793
- Hartsock A, Nelson WJ. Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochim Biophys Acta 2008; 1778:660 - 9; http://dx.doi.org/10.1016/j.bbamem.2007.07.012; PMID: 17854762
- Miyoshi J, Takai Y. Structural and functional associations of apical junctions with cytoskeleton. Biochim Biophys Acta 2008; 1778:670 - 91; http://dx.doi.org/10.1016/j.bbamem.2007.12.014; PMID: 18201548
- Paris L, Tonutti L, Vannini C, Bazzoni G. Structural organization of the tight junctions. Biochim Biophys Acta 2008; 1778:646 - 59; http://dx.doi.org/10.1016/j.bbamem.2007.08.004; PMID: 17945185
- Farkas AE, Capaldo CT, Nusrat A. Regulation of epithelial proliferation by tight junction proteins. Ann N Y Acad Sci 2012; 1258:115 - 24; http://dx.doi.org/10.1111/j.1749-6632.2012.06556.x; PMID: 22731724
- Shen L. Tight junctions on the move: molecular mechanisms for epithelial barrier regulation. Ann N Y Acad Sci 2012; 1258:9 - 18; http://dx.doi.org/10.1111/j.1749-6632.2012.06613.x; PMID: 22731710
- Spadaro D, Tapia R, Pulimeno P, Citi S. The control of gene expression and cell proliferation by the epithelial apical junctional complex. Essays Biochem 2012; 53:83 - 93; http://dx.doi.org/10.1042/bse0530083; PMID: 22928510
- Dimitratos SD, Woods DF, Stathakis DG, Bryant PJ. Signaling pathways are focused at specialized regions of the plasma membrane by scaffolding proteins of the MAGUK family. Bioessays 1999; 21:912 - 21; http://dx.doi.org/10.1002/(SICI)1521-1878(199911)21:11<912::AID-BIES3>3.0.CO;2-Z; PMID: 10517864
- Funke L, Dakoji S, Bredt DS. Membrane-associated guanylate kinases regulate adhesion and plasticity at cell junctions. Annu Rev Biochem 2005; 74:219 - 45; http://dx.doi.org/10.1146/annurev.biochem.74.082803.133339; PMID: 15952887
- González-Mariscal L, Betanzos A, Avila-Flores A. MAGUK proteins: structure and role in the tight junction. Semin Cell Dev Biol 2000; 11:315 - 24; http://dx.doi.org/10.1006/scdb.2000.0178; PMID: 10966866
- te Velthuis AJ, Admiraal JF, Bagowski CP. Molecular evolution of the MAGUK family in metazoan genomes. BMC Evol Biol 2007; 7:129; http://dx.doi.org/10.1186/1471-2148-7-129; PMID: 17678554
- Olsen O, Bredt DS. Functional analysis of the nucleotide binding domain of membrane-associated guanylate kinases. J Biol Chem 2003; 278:6873 - 8; http://dx.doi.org/10.1074/jbc.M210165200; PMID: 12482754
- Gumbiner B, Lowenkopf T, Apatira D. Identification of a 160-kDa polypeptide that binds to the tight junction protein ZO-1. Proc Natl Acad Sci U S A 1991; 88:3460 - 4; http://dx.doi.org/10.1073/pnas.88.8.3460; PMID: 2014265
- Haskins J, Gu L, Wittchen ES, Hibbard J, Stevenson BR. ZO-3, a novel member of the MAGUK protein family found at the tight junction, interacts with ZO-1 and occludin. J Cell Biol 1998; 141:199 - 208; http://dx.doi.org/10.1083/jcb.141.1.199; PMID: 9531559
- Jesaitis LA, Goodenough DA. Molecular characterization and tissue distribution of ZO-2, a tight junction protein homologous to ZO-1 and the Drosophila discs-large tumor suppressor protein. J Cell Biol 1994; 124:949 - 61; http://dx.doi.org/10.1083/jcb.124.6.949; PMID: 8132716
- Stevenson BR, Siliciano JD, Mooseker MS, Goodenough DA. Identification of ZO-1: a high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J Cell Biol 1986; 103:755 - 66; http://dx.doi.org/10.1083/jcb.103.3.755; PMID: 3528172
- Bauer H, Zweimueller-Mayer J, Steinbacher P, Lametschwandtner A, Bauer HC. The dual role of zonula occludens (ZO) proteins. J Biomed Biotechnol 2010; 2010:402593; http://dx.doi.org/10.1155/2010/402593; PMID: 20224657
- Fanning AS, Anderson JM. Zonula occludens-1 and -2 are cytosolic scaffolds that regulate the assembly of cellular junctions. Ann N Y Acad Sci 2009; 1165:113 - 20; http://dx.doi.org/10.1111/j.1749-6632.2009.04440.x; PMID: 19538295
- Fanning AS, Ma TY, Anderson JM. Isolation and functional characterization of the actin binding region in the tight junction protein ZO-1. FASEB J 2002; 16:1835 - 7; PMID: 12354695
- Mattagajasingh SN, Huang SC, Hartenstein JS, Benz EJ Jr.. Characterization of the interaction between protein 4.1R and ZO-2. A possible link between the tight junction and the actin cytoskeleton. J Biol Chem 2000; 275:30573 - 85; http://dx.doi.org/10.1074/jbc.M004578200; PMID: 10874042
- Wittchen ES, Haskins J, Stevenson BR. Protein interactions at the tight junction. Actin has multiple binding partners, and ZO-1 forms independent complexes with ZO-2 and ZO-3. J Biol Chem 1999; 274:35179 - 85; http://dx.doi.org/10.1074/jbc.274.49.35179; PMID: 10575001
- Fanning AS, Little BP, Rahner C, Utepbergenov D, Walther Z, Anderson JM. The unique-5 and -6 motifs of ZO-1 regulate tight junction strand localization and scaffolding properties. Mol Biol Cell 2007; 18:721 - 31; http://dx.doi.org/10.1091/mbc.E06-08-0764; PMID: 17182847
- Adamsky K, Arnold K, Sabanay H, Peles E. Junctional protein MAGI-3 interacts with receptor tyrosine phosphatase beta (RPTP beta) and tyrosine-phosphorylated proteins. J Cell Sci 2003; 116:1279 - 89; http://dx.doi.org/10.1242/jcs.00302; PMID: 12615970
- Dobrosotskaya I, Guy RK, James GL. MAGI-1, a membrane-associated guanylate kinase with a unique arrangement of protein-protein interaction domains. J Biol Chem 1997; 272:31589 - 97; http://dx.doi.org/10.1074/jbc.272.50.31589; PMID: 9395497
- Wood JD, Yuan J, Margolis RL, Colomer V, Duan K, Kushi J, et al. Atrophin-1, the DRPLA gene product, interacts with two families of WW domain-containing proteins. Mol Cell Neurosci 1998; 11:149 - 60; http://dx.doi.org/10.1006/mcne.1998.0677; PMID: 9647693
- Caruana G. Genetic studies define MAGUK proteins as regulators of epithelial cell polarity. Int J Dev Biol 2002; 46:511 - 8; PMID: 12141438
- Van Campenhout CA, Eitelhuber A, Gloeckner CJ, Giallonardo P, Gegg M, Oller H, et al. Dlg3 trafficking and apical tight junction formation is regulated by nedd4 and nedd4-2 e3 ubiquitin ligases. Dev Cell 2011; 21:479 - 91; http://dx.doi.org/10.1016/j.devcel.2011.08.003; PMID: 21920314
- Rivera C, Simonson SJ, Yamben IF, Shatadal S, Nguyen MM, Beurg M, et al. Requirement for Dlgh-1 in planar cell polarity and skeletogenesis during vertebrate development. PLoS One 2013; 8:e54410; http://dx.doi.org/10.1371/journal.pone.0054410; PMID: 23349879
- Chi CN, Bach A, Strømgaard K, Gianni S, Jemth P. Ligand binding by PDZ domains. Biofactors 2012; 38:338 - 48; http://dx.doi.org/10.1002/biof.1031; PMID: 22674855
- Jeleń F, Oleksy A, Smietana K, Otlewski J. PDZ domains - common players in the cell signaling. Acta Biochim Pol 2003; 50:985 - 1017; PMID: 14739991
- Hamazaki Y, Itoh M, Sasaki H, Furuse M, Tsukita S. Multi-PDZ domain protein 1 (MUPP1) is concentrated at tight junctions through its possible interaction with claudin-1 and junctional adhesion molecule. J Biol Chem 2002; 277:455 - 61; http://dx.doi.org/10.1074/jbc.M109005200; PMID: 11689568
- Furuse M, Itoh M, Hirase T, Nagafuchi A, Yonemura S, Tsukita S, et al. Direct association of occludin with ZO-1 and its possible involvement in the localization of occludin at tight junctions. J Cell Biol 1994; 127:1617 - 26; http://dx.doi.org/10.1083/jcb.127.6.1617; PMID: 7798316
- Itoh M, Furuse M, Morita K, Kubota K, Saitou M, Tsukita S. Direct binding of three tight junction-associated MAGUKs, ZO-1, ZO-2, and ZO-3, with the COOH termini of claudins. J Cell Biol 1999; 147:1351 - 63; http://dx.doi.org/10.1083/jcb.147.6.1351; PMID: 10601346
- Jeansonne B, Lu Q, Goodenough DA, Chen YH. Claudin-8 interacts with multi-PDZ domain protein 1 (MUPP1) and reduces paracellular conductance in epithelial cells. Cell Mol Biol (Noisy-le-grand) 2003; 49:13 - 21; PMID: 12839333
- Ebnet K, Suzuki A, Horikoshi Y, Hirose T, Meyer Zu Brickwedde MK, Ohno S, et al. The cell polarity protein ASIP/PAR-3 directly associates with junctional adhesion molecule (JAM). EMBO J 2001; 20:3738 - 48; http://dx.doi.org/10.1093/emboj/20.14.3738; PMID: 11447115
- Itoh M, Sasaki H, Furuse M, Ozaki H, Kita T, Tsukita S. Junctional adhesion molecule (JAM) binds to PAR-3: a possible mechanism for the recruitment of PAR-3 to tight junctions. J Cell Biol 2001; 154:491 - 7; http://dx.doi.org/10.1083/jcb.200103047; PMID: 11489913
- Roh MH, Makarova O, Liu CJ, Shin K, Lee S, Laurinec S, et al. The Maguk protein, Pals1, functions as an adapter, linking mammalian homologues of Crumbs and Discs Lost. J Cell Biol 2002; 157:161 - 72; http://dx.doi.org/10.1083/jcb.200109010; PMID: 11927608
- Métais JY, Navarro C, Santoni MJ, Audebert S, Borg JP. hScrib interacts with ZO-2 at the cell-cell junctions of epithelial cells. FEBS Lett 2005; 579:3725 - 30; http://dx.doi.org/10.1016/j.febslet.2005.05.062; PMID: 15975580
- Ivanov AI, Young C, Den Beste K, Capaldo CT, Humbert PO, Brennwald P, et al. Tumor suppressor scribble regulates assembly of tight junctions in the intestinal epithelium. Am J Pathol 2010; 176:134 - 45; http://dx.doi.org/10.2353/ajpath.2010.090220; PMID: 19959811
- Yamamoto T, Harada N, Kano K, Taya S, Canaani E, Matsuura Y, et al. The Ras target AF-6 interacts with ZO-1 and serves as a peripheral component of tight junctions in epithelial cells. J Cell Biol 1997; 139:785 - 95; http://dx.doi.org/10.1083/jcb.139.3.785; PMID: 9348294
- Fanning AS, Lye MF, Anderson JM, Lavie A. Domain swapping within PDZ2 is responsible for dimerization of ZO proteins. J Biol Chem 2007; 282:37710 - 6; http://dx.doi.org/10.1074/jbc.M707255200; PMID: 17928286
- Itoh M, Morita K, Tsukita S. Characterization of ZO-2 as a MAGUK family member associated with tight as well as adherens junctions with a binding affinity to occludin and alpha catenin. J Biol Chem 1999; 274:5981 - 6; http://dx.doi.org/10.1074/jbc.274.9.5981; PMID: 10026224
- Li X, Lu S, Nagy JI. Direct association of connexin36 with zonula occludens-2 and zonula occludens-3. Neurochem Int 2009; 54:393 - 402; http://dx.doi.org/10.1016/j.neuint.2009.01.003; PMID: 19418635
- Singh D, Solan JL, Taffet SM, Javier R, Lampe PD. Connexin 43 interacts with zona occludens-1 and -2 proteins in a cell cycle stage-specific manner. J Biol Chem 2005; 280:30416 - 21; http://dx.doi.org/10.1074/jbc.M506799200; PMID: 15980428
- Schmidt A, Utepbergenov DI, Mueller SL, Beyermann M, Schneider-Mergener J, Krause G, et al. Occludin binds to the SH3-hinge-GuK unit of zonula occludens protein 1: potential mechanism of tight junction regulation. Cell Mol Life Sci 2004; 61:1354 - 65; http://dx.doi.org/10.1007/s00018-004-4010-6; PMID: 15170513
- Citi S, Paschoud S, Pulimeno P, Timolati F, De Robertis F, Jond L, et al. The tight junction protein cingulin regulates gene expression and RhoA signaling. Ann N Y Acad Sci 2009; 1165:88 - 98; http://dx.doi.org/10.1111/j.1749-6632.2009.04053.x; PMID: 19538293
- Cordenonsi M, D’Atri F, Hammar E, Parry DA, Kendrick-Jones J, Shore D, et al. Cingulin contains globular and coiled-coil domains and interacts with ZO-1, ZO-2, ZO-3, and myosin. J Cell Biol 1999; 147:1569 - 82; http://dx.doi.org/10.1083/jcb.147.7.1569; PMID: 10613913
- Jin J, Smith FD, Stark C, Wells CD, Fawcett JP, Kulkarni S, et al. Proteomic, functional, and domain-based analysis of in vivo 14-3-3 binding proteins involved in cytoskeletal regulation and cellular organization. Curr Biol 2004; 14:1436 - 50; http://dx.doi.org/10.1016/j.cub.2004.07.051; PMID: 15324660
- Avila-Flores A, Rendón-Huerta E, Moreno J, Islas S, Betanzos A, Robles-Flores M, et al. Tight-junction protein zonula occludens 2 is a target of phosphorylation by protein kinase C. Biochem J 2001; 360:295 - 304; http://dx.doi.org/10.1042/0264-6021:3600295; PMID: 11716757
- Chamorro D, Alarcón L, Ponce A, Tapia R, González-Aguilar H, Robles-Flores M, et al. Phosphorylation of zona occludens-2 by protein kinase C epsilon regulates its nuclear exportation. Mol Biol Cell 2009; 20:4120 - 9; http://dx.doi.org/10.1091/mbc.E08-11-1129; PMID: 19625451
- Koizumi J, Kojima T, Ogasawara N, Kamekura R, Kurose M, Go M, et al. Protein kinase C enhances tight junction barrier function of human nasal epithelial cells in primary culture by transcriptional regulation. Mol Pharmacol 2008; 74:432 - 42; http://dx.doi.org/10.1124/mol.107.043711; PMID: 18477669
- Saito K, Enya K, Oneyama C, Hikita T, Okada M. Proteomic identification of ZO-1/2 as a novel scaffold for Src/Csk regulatory circuit. Biochem Biophys Res Commun 2008; 366:969 - 75; http://dx.doi.org/10.1016/j.bbrc.2007.12.055; PMID: 18086565
- Meyer TN, Schwesinger C, Denker BM. Zonula occludens-1 is a scaffolding protein for signaling molecules. Galpha(12) directly binds to the Src homology 3 domain and regulates paracellular permeability in epithelial cells. J Biol Chem 2002; 277:24855 - 8; http://dx.doi.org/10.1074/jbc.C200240200; PMID: 12023272
- Sabath E, Negoro H, Beaudry S, Paniagua M, Angelow S, Shah J, et al. Galpha12 regulates protein interactions within the MDCK cell tight junction and inhibits tight-junction assembly. J Cell Sci 2008; 121:814 - 24; http://dx.doi.org/10.1242/jcs.014878; PMID: 18285450
- Lopez-Bayghen E, Jaramillo B, Huerta M, Betanzos A, Gonzalez-Mariscal L. TJ Proteins That Make Round Trips to the Nucleus. Tight Junctions: Springer US, 2006:76-100.
- González-Mariscal L, Ponce A, Alarcón L, Jaramillo BE. The tight junction protein ZO-2 has several functional nuclear export signals. Exp Cell Res 2006; 312:3323 - 35; http://dx.doi.org/10.1016/j.yexcr.2006.07.006; PMID: 16920099
- Gottardi CJ, Arpin M, Fanning AS, Louvard D. The junction-associated protein, zonula occludens-1, localizes to the nucleus before the maturation and during the remodeling of cell-cell contacts. Proc Natl Acad Sci U S A 1996; 93:10779 - 84; http://dx.doi.org/10.1073/pnas.93.20.10779; PMID: 8855257
- Islas S, Vega J, Ponce L, González-Mariscal L. Nuclear localization of the tight junction protein ZO-2 in epithelial cells. Exp Cell Res 2002; 274:138 - 48; http://dx.doi.org/10.1006/excr.2001.5457; PMID: 11855865
- Jaramillo BE, Ponce A, Moreno J, Betanzos A, Huerta M, Lopez-Bayghen E, et al. Characterization of the tight junction protein ZO-2 localized at the nucleus of epithelial cells. Exp Cell Res 2004; 297:247 - 58; http://dx.doi.org/10.1016/j.yexcr.2004.03.021; PMID: 15194440
- Traweger A, Fuchs R, Krizbai IA, Weiger TM, Bauer HC, Bauer H. The tight junction protein ZO-2 localizes to the nucleus and interacts with the heterogeneous nuclear ribonucleoprotein scaffold attachment factor-B. J Biol Chem 2003; 278:2692 - 700; http://dx.doi.org/10.1074/jbc.M206821200; PMID: 12403786
- Tapia R, Huerta M, Islas S, Avila-Flores A, Lopez-Bayghen E, Weiske J, et al. Zona occludens-2 inhibits cyclin D1 expression and cell proliferation and exhibits changes in localization along the cell cycle. Mol Biol Cell 2009; 20:1102 - 17; http://dx.doi.org/10.1091/mbc.E08-03-0277; PMID: 19056685
- Meerschaert K, Tun MP, Remue E, De Ganck A, Boucherie C, Vanloo B, et al. The PDZ2 domain of zonula occludens-1 and -2 is a phosphoinositide binding domain. Cell Mol Life Sci 2009; 66:3951 - 66; http://dx.doi.org/10.1007/s00018-009-0156-6; PMID: 19784548
- Willott E, Balda MS, Fanning AS, Jameson B, Van Itallie C, Anderson JM. The tight junction protein ZO-1 is homologous to the Drosophila discs-large tumor suppressor protein of septate junctions. Proc Natl Acad Sci U S A 1993; 90:7834 - 8; http://dx.doi.org/10.1073/pnas.90.16.7834; PMID: 8395056
- Woods DF, Bryant PJ. The discs-large tumor suppressor gene of Drosophila encodes a guanylate kinase homolog localized at septate junctions. Cell 1991; 66:451 - 64; http://dx.doi.org/10.1016/0092-8674(81)90009-X; PMID: 1651169
- Hernandez S, Chavez Munguia B, Gonzalez-Mariscal L. ZO-2 silencing in epithelial cells perturbs the gate and fence function of tight junctions and leads to an atypical monolayer architecture. Exp Cell Res 2007; 313:1533 - 47; http://dx.doi.org/10.1016/j.yexcr.2007.01.026; PMID: 17374535
- Xu J, Kausalya PJ, Phua DC, Ali SM, Hossain Z, Hunziker W. Early embryonic lethality of mice lacking ZO-2, but Not ZO-3, reveals critical and nonredundant roles for individual zonula occludens proteins in mammalian development. Mol Cell Biol 2008; 28:1669 - 78; http://dx.doi.org/10.1128/MCB.00891-07; PMID: 18172007
- Traweger A, Lehner C, Farkas A, Krizbai IA, Tempfer H, Klement E, et al. Nuclear Zonula occludens-2 alters gene expression and junctional stability in epithelial and endothelial cells. Differentiation 2008; 76:99 - 106; PMID: 17973926
- Huerta M, Muñoz R, Tapia R, Soto-Reyes E, Ramírez L, Recillas-Targa F, et al. Cyclin D1 is transcriptionally down-regulated by ZO-2 via an E box and the transcription factor c-Myc. Mol Biol Cell 2007; 18:4826 - 36; http://dx.doi.org/10.1091/mbc.E07-02-0109; PMID: 17881732
- Lechuga S, Alarcón L, Solano J, Huerta M, Lopez-Bayghen E, González-Mariscal L. Identification of ZASP, a novel protein associated to Zona occludens-2. Exp Cell Res 2010; 316:3124 - 39; http://dx.doi.org/10.1016/j.yexcr.2010.09.008; PMID: 20868680
- Oka T, Remue E, Meerschaert K, Vanloo B, Boucherie C, Gfeller D, et al. Functional complexes between YAP2 and ZO-2 are PDZ domain-dependent, and regulate YAP2 nuclear localization and signalling. Biochem J 2010; 432:461 - 72; http://dx.doi.org/10.1042/BJ20100870; PMID: 20868367
- Oka T, Sudol M. Nuclear localization and pro-apoptotic signaling of YAP2 require intact PDZ-binding motif. Genes Cells 2009; 14:607 - 15; http://dx.doi.org/10.1111/j.1365-2443.2009.01292.x; PMID: 19371381
- Remue E, Meerschaert K, Oka T, Boucherie C, Vandekerckhove J, Sudol M, et al. TAZ interacts with zonula occludens-1 and -2 proteins in a PDZ-1 dependent manner. FEBS Lett 2010; 584:4175 - 80; http://dx.doi.org/10.1016/j.febslet.2010.09.020; PMID: 20850437
- Mihlan S, Reiss C, Thalheimer P, Herterich S, Gaetzner S, Kremerskothen J, et al. Nuclear import of LASP-1 is regulated by phosphorylation and dynamic protein-protein interactions. Oncogene 2012; 32:2107 - 13; PMID: 22665060
- Kausalya PJ, Phua DC, Hunziker W. Association of ARVCF with zonula occludens (ZO)-1 and ZO-2: binding to PDZ-domain proteins and cell-cell adhesion regulate plasma membrane and nuclear localization of ARVCF. Mol Biol Cell 2004; 15:5503 - 15; http://dx.doi.org/10.1091/mbc.E04-04-0350; PMID: 15456900
- Betanzos A, Huerta M, Lopez-Bayghen E, Azuara E, Amerena J, González-Mariscal L. The tight junction protein ZO-2 associates with Jun, Fos and C/EBP transcription factors in epithelial cells. Exp Cell Res 2004; 292:51 - 66; http://dx.doi.org/10.1016/j.yexcr.2003.08.007; PMID: 14720506
- Renz A, Fackelmayer FO. Purification and molecular cloning of the scaffold attachment factor B (SAF-B), a novel human nuclear protein that specifically binds to S/MAR-DNA. Nucleic Acids Res 1996; 24:843 - 9; http://dx.doi.org/10.1093/nar/24.5.843; PMID: 8600450
- Alfonso-Parra C, Maggert KA. Drosophila SAF-B links the nuclear matrix, chromosomes, and transcriptional activity. PLoS One 2010; 5:e10248; http://dx.doi.org/10.1371/journal.pone.0010248; PMID: 20422039
- Kusch A, Tkachuk S, Tkachuk N, Patecki M, Park JK, Dietz R, et al. The tight junction protein ZO-2 mediates proliferation of vascular smooth muscle cells via regulation of Stat1. Cardiovasc Res 2009; 83:115 - 22; http://dx.doi.org/10.1093/cvr/cvp117; PMID: 19380416
- Tkachuk N, Tkachuk S, Patecki M, Kusch A, Korenbaum E, Haller H, et al. The tight junction protein ZO-2 and Janus kinase 1 mediate intercellular communications in vascular smooth muscle cells. Biochem Biophys Res Commun 2011; 410:531 - 6; http://dx.doi.org/10.1016/j.bbrc.2011.06.017; PMID: 21679692
- Bautista-García P, Reyes JL, Martín D, Namorado MC, Chavez-Munguía B, Soria-Castro E, et al. Zona occludens-2 protects against podocyte dysfunction induced by ADR in mice. Am J Physiol Renal Physiol 2013; 304:F77 - 87; http://dx.doi.org/10.1152/ajprenal.00089.2012; PMID: 23034938
- Gewurz BE, Towfic F, Mar JC, Shinners NP, Takasaki K, Zhao B, et al. Genome-wide siRNA screen for mediators of NF-κB activation. Proc Natl Acad Sci U S A 2012; 109:2467 - 72; http://dx.doi.org/10.1073/pnas.1120542109; PMID: 22308454
- Op de Beeck K, Schacht J, Van Camp G. Apoptosis in acquired and genetic hearing impairment: the programmed death of the hair cell. Hear Res 2011; 281:18 - 27; http://dx.doi.org/10.1016/j.heares.2011.07.002; PMID: 21782914
- Walsh T, Pierce SB, Lenz DR, Brownstein Z, Dagan-Rosenfeld O, Shahin H, et al. Genomic duplication and overexpression of TJP2/ZO-2 leads to altered expression of apoptosis genes in progressive nonsyndromic hearing loss DFNA51. Am J Hum Genet 2010; 87:101 - 9; http://dx.doi.org/10.1016/j.ajhg.2010.05.011; PMID: 20602916
- Gonzalez-Mariscal L, Bautista P, Lechuga S, Quiros M. ZO-2, a tight junction scaffold protein involved in the regulation of cell proliferation and apoptosis. Ann N Y Acad Sci 2012; 1257:133 - 41; http://dx.doi.org/10.1111/j.1749-6632.2012.06537.x; PMID: 22671599
- Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest 2009; 119:1420 - 8; http://dx.doi.org/10.1172/JCI39104; PMID: 19487818
- Oesterreich S, Lee AV, Sullivan TM, Samuel SK, Davie JR, Fuqua SA. Novel nuclear matrix protein HET binds to and influences activity of the HSP27 promoter in human breast cancer cells. J Cell Biochem 1997; 67:275 - 86; http://dx.doi.org/10.1002/(SICI)1097-4644(19971101)67:2<275::AID-JCB13>3.0.CO;2-E; PMID: 9328833
- Lehner C, Gehwolf R, Tempfer H, Krizbai I, Hennig B, Bauer HC, et al. Oxidative stress and blood-brain barrier dysfunction under particular consideration of matrix metalloproteinases. Antioxid Redox Signal 2011; 15:1305 - 23; http://dx.doi.org/10.1089/ars.2011.3923; PMID: 21294658
- Cui J, Chen S, Zhang C, Meng F, Wu W, Hu R, et al. Inhibition of MMP-9 by a selective gelatinase inhibitor protects neurovasculature from embolic focal cerebral ischemia. Mol Neurodegener 2012; 7:21; http://dx.doi.org/10.1186/1750-1326-7-21; PMID: 22587708
- Liu W, Hendren J, Qin XJ, Shen J, Liu KJ. Normobaric hyperoxia attenuates early blood-brain barrier disruption by inhibiting MMP-9-mediated occludin degradation in focal cerebral ischemia. J Neurochem 2009; 108:811 - 20; http://dx.doi.org/10.1111/j.1471-4159.2008.05821.x; PMID: 19187098
- Romanic AM, White RF, Arleth AJ, Ohlstein EH, Barone FC. Matrix metalloproteinase expression increases after cerebral focal ischemia in rats: inhibition of matrix metalloproteinase-9 reduces infarct size. Stroke 1998; 29:1020 - 30; http://dx.doi.org/10.1161/01.STR.29.5.1020; PMID: 9596253
- Rosenberg GA, Estrada EY, Dencoff JE. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke 1998; 29:2189 - 95; http://dx.doi.org/10.1161/01.STR.29.10.2189; PMID: 9756602
- Yang Y, Estrada EY, Thompson JF, Liu W, Rosenberg GA. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J Cereb Blood Flow Metab 2007; 27:697 - 709; PMID: 16850029
- Kitamuro T, Takahashi K, Ogawa K, Udono-Fujimori R, Takeda K, Furuyama K, et al. Bach1 functions as a hypoxia-inducible repressor for the heme oxygenase-1 gene in human cells. J Biol Chem 2003; 278:9125 - 33; http://dx.doi.org/10.1074/jbc.M209939200; PMID: 12511571
- Lin CW, Shen SC, Hou WC, Yang LY, Chen YC. Heme oxygenase-1 inhibits breast cancer invasion via suppressing the expression of matrix metalloproteinase-9. Mol Cancer Ther 2008; 7:1195 - 206; http://dx.doi.org/10.1158/1535-7163.MCT-07-2199; PMID: 18483307
- Yang B, Akhter S, Chaudhuri A, Kanmogne GD. HIV-1 gp120 induces cytokine expression, leukocyte adhesion, and transmigration across the blood-brain barrier: modulatory effects of STAT1 signaling. Microvasc Res 2009; 77:212 - 9; http://dx.doi.org/10.1016/j.mvr.2008.11.003; PMID: 19103208
- Javier RT, Rice AP. Emerging theme: cellular PDZ proteins as common targets of pathogenic viruses. J Virol 2011; 85:11544 - 56; http://dx.doi.org/10.1128/JVI.05410-11; PMID: 21775458
- Latorre IJ, Roh MH, Frese KK, Weiss RS, Margolis B, Javier RT. Viral oncoprotein-induced mislocalization of select PDZ proteins disrupts tight junctions and causes polarity defects in epithelial cells. J Cell Sci 2005; 118:4283 - 93; http://dx.doi.org/10.1242/jcs.02560; PMID: 16141229
- González-Mariscal L, Quirós M, Díaz-Coránguez M. ZO proteins and redox-dependent processes. Antioxid Redox Signal 2011; 15:1235 - 53; http://dx.doi.org/10.1089/ars.2011.3913; PMID: 21294657