Abstract
Tissue barriers play an integral role in the biology and pathobiology of mammary ductal epithelium. In normal breast physiology, tight and adherens junctions undergo dynamic changes in permeability in response to hormonal and other stimuli, while several of their proteins are directly involved in mammary tumorigenesis. This review describes first the structure of mammary ductal epithelial barriers and their role in normal mammary development, examining the cyclical changes in response to puberty, pregnancy, lactation and involution. It then examines the role of adherens and tight junctions and the participation of their constituent proteins in mammary tumorigenic functions such as migration, invasion and metastasis. Finally, it discusses the potential of these adhesion proteins as both prognostic biomarkers and potential therapeutic targets in breast cancer.
Introduction
Breast cancer is the commonest malignancy among women worldwide, with over 1.3 million new cases diagnosed and over 450,000 deaths per annum.Citation1 While the prognosis of breast cancer has improved significantly in recent years, its mortality remains significant. Despite much research into targeted treatment modalities, treatment remains focused on surgical excision, chemotherapy, radiotherapy and hormone therapy, with the anti-HER2 monoclonal antibody trastuzumab currently the only targeted therapy in routine clinical use. As many of these treatment modalities are associated with significant side effects, the quest continues to discover biomarkers identifying those likely to benefit most from adjuvant treatment, and novel targets for development of anti-breast cancer agents with minimal systemic side-effects.
Proteins located in tight and adherens junctions are obvious candidates for such biomarkers and/or therapeutic targets for a number of reasons: first, cellular junctions encourage physical cell-cell associations that must theoretically be overcome to allow tumor cell shedding and distal metastasis, and second, many of the proteins in tight and adherens junctions are also involved in pro-proliferative and pro-migratory signaling cascades relevant to cancer progression (for review seeCitation2). As the majority of breast cancer mortality is due to metastatic disease rather than the primary tumor, the development of strategies to prevent such distal sp.read is particularly crucial.
This review aims to examine the role of ductal adhesion complexes and their constituent proteins in both the normal and diseased breast. We will first discuss their structure and physiology under normal circumstances such as puberty, pregnancy and lactation. Next we will examine their role in abnormal breast conditions, both inflammatory and neoplastic, but will focus on breast cancer as the most significant and best studied pathological condition of the breast. We will discuss the role of cell junctions and their proteins in such neoplastic behaviors as dysregulated proliferation, migration, invasion and metastasis. Finally, we will examine the potential use of junctional proteins both as breast cancer prognostic biomarkers and as novel therapeutic targets.
Structure of Mammary Ductal Epithelium
The mammary gland () is a modified apocrine sweat gland that consists of multiple pyramidal lobes each subdivided into several lobules, which in turn consist of multiple acini. These drain via a complex network of branching ducts, eventually conveying milk to the nipple. The basic functional unit is the terminal duct-lobular unit (TDLU), consisting of a lobule and its draining duct, which is supported within a network of fat and connective tissue.Citation3
Figure 1. Normal physiology of the human breast.
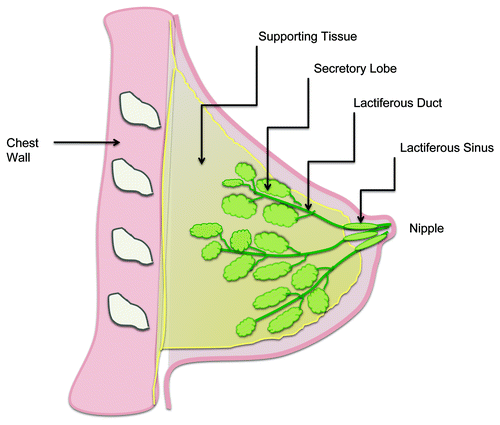
Microscopically, mammary ducts are lined by a single luminal layer of columnar or cuboidal epithelial cells surrounded by a discontinuous layer of contractile myoepithelial cells, in turn surrounded by basement membrane.Citation4 Like other epithelial layers, mammary ductal epithelial cells are joined at their cell-cell interfaces by junctional complexes; which include tight junctions, adherens junctions and desmosomes.
The tight junction forms a continuous band around the cell at the apical-most surface, effectively dividing it into apical and basolateral compartments and regulating the cellular barrier and paracellular transport.Citation5 Tight junction proteins can be broadly divided into integral transmembrane proteins such as occludin, claudins and junctional adhesion molecules (JAMs); peripheral or plaque adaptor proteins such as the Zona Occludens (ZO) proteins; and regulatory/signaling proteins such as cingulin and the Rho-GTPases.Citation2 Adherens junctions are located subjacent to tight junctions, forming a band around the cell and attaching the actin cytoskeleton to the plasma membrane.Citation6 Their proteins include the armadillo or armadillo-related proteins (β-catenin, plakoglobin, p120); cytoskeletal adaptor proteins like α−catenin; and the cadherins (E-cadherin in normal epithelial cells, N-cadherin in mesenchymal cells).Citation6-Citation8 Desmosomes appear as patches subjacent to adherens junctions. They anchor keratinous intermediate filaments to the plasma membrane, while their proteins include cadherins (desmogleins, desmocollins), armadillo proteins (plakoglobin, plakophilins) and desmoplakin.Citation8 The majority of this review will concentrate on the contribution of tight and adherens junction proteins to breast physiology and pathophysiology, principally because comparatively little has been published about breast desmosomal complexes. Nonetheless, desmosomal loss has recently been shown to be important for branching morphogenesis, the mammary remodeling process which ultimately permits lactation.Citation9 In fact desmosomal cadherins participate in epithelial-myoepithelial interactions in the normal mouse mammary gland, and selective desmosomal loss during the secretory phase of mammary developmentCitation10 likely underlies the development of myoepithelial discontinuities in lactating rat mammary glands.Citation11 The contribution of desmosomal alterations to pathologies such as cancer is also relatively understudied, though one might sp.eculate that any loss of mechanical adhesion could facilitate local invasion of single cells potentially facilitating metastasis. That said, mechanically-strong cell-cell attachments might also allow multi-cell clusters to move in so-called “Indian files,” a collective invasion phenomenon characteristic of some breast cancers. In support of the former sp.eculation, loss of at least one desmosomal protein, desmoplakin, has been implicated in breast tumor progression.Citation12 Accordingly, it has been noted that estrogen binding to ERα, a hallmark of well-differentiated / less-invasive breast cancers, promotes adhesion via morphological enhancement of desmosome numbers and upregulation of the desmosomal proteins desmocollin, γ-catenin, plakophilin and desmoplakin.Citation13 Since liganded ERα also promotes transcription of the retinoic acid receptor α gene,Citation14 itself a positive regulator of cell-matrix adhesion in breast cancer cells,Citation15 this supports a model whereby hormone receptor expression, in part by promoting a pro-adhesion state, is associated with favorable differentiation status in breast cancer.
As detailed below, several of the above adhesion structures are modulated dynamically in time with the normal reproductive and hormonal cycle, in addition to being altered in many pathological processes.
Models for Mammary Epithelial Research
As in most cell biology, research on mammary epithelial biology relies largely on in vivo and in vitro models. In vitro cell culture models, predominantly using commercially available epithelial cell lines, are the most common vehicle for researching mammary epithelial barriers. These offer the advantages of cost effective, readily available, uniform cell populations that are genetically well-characterized and grow readily and predictably in vitro. However, there are a small number of cell lines available, with only three lines (MCF 7, MDA-MB 231, T-47D) accounting for over two thirds of all abstracts reporting work on breast cancer derived cell lines appearing on Medline.Citation16 Given the reliance of a huge number of researchers on a small number of cell lines, cross-contamination between cell lines is an important problem, with one study suggesting 18% of cell lines may be affected.Citation17 Furthermore, an immortalized cell line selects only the clonal population that exhibits most proliferation under a given set of artificial conditions, most commonly derived from a metastatic cancer, and is thus inherently different from both the primary tumor and normal epithelium. Indeed, the in vitro environment, which usually consists of a clonal cell population adherent to a flat plastic surface, bathed in nutrient-rich, antibiotic-containing media; is quite different to the natural physiological milieu in which polyclonal cells proliferate in three dimensions, interacting both with other epithelial cells and with a complex supporting stroma.
In the in vivo setting, recent years have seen an explosion in the number and diversity of mouse mammary models of cancer, ranging from genetically-modified animals to mammary fatpad injectible tumor models to patient-derived xenograft studies in the emerging era of semi-personalized medicine (reviewed inCitation18). Enhanced by the recent adoption of non-invasive fluorescent tumor imaging technologies, these have collectively proven invaluable for unraveling aspects of the complex biology of breast cancer. For example, the human HER2 gene, whose amplification is linked with certain aggressive breast cancers, was originally identified as the pro-tumorigenic murine oncogene neuCitation19; and in fact murine models still represent the gold standard pre-clinical approach for testing potential anti-cancer drugs. Interestingly however, there has been a relative scarcity of models for studying the fundamental physiology (rather than pathophysiology) of the mammary epithelial barrier. This may relate in part to technical difficulties in experimentally manipulating mammary tissue; although the economic importance of estimating (for example) drug accumulation in the colostrum of lactating animals has been an agricultural issue for decades. Nonetheless, important work in rodent mammary barriers has revealed several useful model systems for physiological assessment of permeability. These include measuring the relative recovery of drugs in serum vs. breast milk following oral gavage of compounds of interest in lactating rats,Citation20 or the assessment of basal-apical transport of radiolabeled albumin following its injection into the bloodstream of a rodent.Citation21 The reverse approach has utilized injection of radiolabeled sucrose or fluorescent albumin directly into mammary ducts followed by periodic blood sampling in order to determine apical-basal tight junction permeability during hormonal events such as the lactogenic switch.Citation22 It is, however, unclear how sp.ecific permeability coefficients in such models might relate to those in the human setting, due to an absence of comprehensive studies on substance accumulation in the breastmilk of lactating human females.
Aside from macromolecular permeability measurements, barrier function and ion transport characteristics of epithelial tissues (particularly intestine, skin and lung) have been classically assessed via electrical measurements in Ussing chambers.Citation23 Both semi-permeable filters (usually polycarbonate or polyester)Citation24,Citation25 and collagen gelsCitation26 have been successfully demonstrated as appropriate physical matrices for subsequent electrical measurements in primary or immortalized mammary cells. However it is interesting to note that mammary cells appear particularly sensitive to the culture microenvironment in terms of their ability to form tight, electrically-sealed barriers; as exemplified by a recent report that the non-transformed normal-like breast cell line MCF-10A can only develop measurable transepithelial electrical resistance in the absence of the adenylyl cyclase activator cholera toxin.Citation27 Our own unpublished observations have noted that these cells require a long period of “conditioning” to the cholera toxin-free environment (approximately one month) before a high resistance monolayer can be formed (Kieran Brennan, personal communication). These findings illustrate at least two essential points – first the complex crosstalk between electrical resistance and ion transport (via established chloride-secretory second messengers such as adenylyl cyclase), and second the importance of exercising caution in interpreting barrier function results from endocrine tissues such as the breast, whose very structure and function is heavily dependent on hormonal status and developmental stage.
Biology of Mammary Ductal Barriers
The physical structure of the mammary ductal epithelial barrier is almost identical to that of other epithelia, with the exception that intact epithelial monolayers lie on a bed of contractile myoepithelial cells () instead of being in direct contact with the basal lamina. The myoepithelial monolayer predominantly functions as a physiological tool to expel milk into the ducts during lactation, while in pathophysiological terms myoepithelial loss is an early hallmark of ductal carcinoma in situ. However it is in its biology, particularly the dynamic alteration of mammary barrier function in response to hormonal changes, that the mammary epithelium is unique. More than most organ systems, the breast undergoes frequent changes in response to puberty, the menstrual cycle, pregnancy, lactation and menopause; and many of these are modulated via alterations in ductal junctional complexes or junctional proteins directly. Signaling via the canonical Wnt pathway involving the adherens junction protein and transcription factor β-catenin has been implicated in virtually every stage of this cycle.Citation28 Wnt/ β-catenin signaling is vital for the formation of the embryonic mammary placode in miceCitation29 and β-catenin target genes are upregulated in murine embryonic ductal morphogenesis.Citation30 Murine in vivo studies have yielded indirect evidence of low level Wnt/ β-catenin signaling occurring during mammary development at puberty, with enrichment of Wnt5a and Wnt7b mRNAs in terminal end buds and that of Wnt2 in mammary stroma.Citation28,Citation31-Citation33 During pregnancy, progesterone-induced changes such as increased ductal branching are modulated via β-catenin signaling in a mouse model.Citation34
Figure 2. Cell-cell adhesion complexes in the breast epithelial barrier.
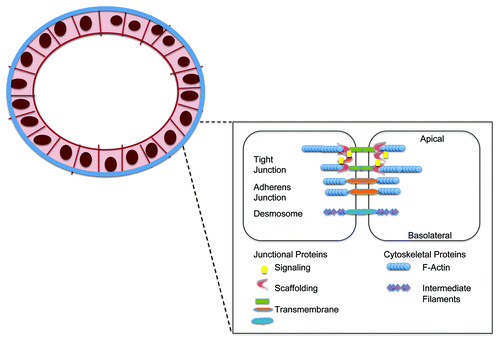
Most of our understanding of the effects of pregnancy and lactation on mammary junctions comes from the agricultural and dairy industry, with minimal research interest in human lactation. Pregnancy is characterized by increased leakiness of the mammary ductal tight junctions in particular, with numerous studies on both dairy goatsCitation35,Citation36 and miceCitation37 showing extravasation of large molecules from the pregnant duct, although to a greater extent in alveolar than ductal epithelium. This is reflected in the loss of trans-epithelial electrical resistance in the mammary epithelium of pregnant animals goats and miceCitation37,Citation38 and the altered composition of milk pre-partum in both humans and dairy animals, with higher concentrations of the interstitial ions sodium and chloride, as well as proteins.Citation35,Citation39,Citation40 In addition, morphological alterations in tight junctions have been observed in mammary epithelium derived from pregnant sheep, with lower numbers of strands and less branching complexity exhibited.Citation41,Citation42
Mammary epithelial tight junctions are altered by several hormones including glucocorticoids,Citation43 prolactin,Citation44 serotoninCitation45 and progesterone, and it is the sharp fall in the latter at parturition that allows tight junction closure to provide a leak-proof duct and restore trans-epithelial resistance, thus facilitating lactation.Citation22 Glucocorticoids such as cortisol are raised throughout pregnancy, and decline during lactation.Citation46,Citation47 There is evidence that glucocorticoid treatment reduces mammary tight junction permeability in both bovineCitation43,Citation44 and ratCitation48,Citation49 in vitro models, via downregulation of the small GTPase RhoACitation50 and phosphorylation of GSK3 by Akt.Citation51 Glucocorticoid treatment has been shown to prevent rat mammary involution in vivo.Citation52 An in vivo study on ovariectomized mice showed that although progesterone withdrawal was the primary trigger for mammary epithelial tight junction closure in late pregnancy (as demonstrated by progesterone antagonism), low to moderate levels of both cortisol and either placental lactogen or prolactin were required for this to happen.Citation22 Unlike progesterone and glucocorticoids, prolactin does not appear to prevent involution in mice,Citation52 although several authors report that prolactin maintains mammary epithelial impermeability in late lactation in rabbits, largely indirectly by preventing apoptosis.Citation53-Citation55 Interestingly, neutrophils are able to pass through these intercellular junctions to reach the lumen if necessary, with complete reconstitution of tight junctions occurring afterwards.Citation56
The neurotransmitter serotonin (5-HT) is an important regulator of lactation. It is locally synthesized in mouse mammary glands.Citation57 In vitro work on human mammary epithelial monolayers has demonstrated that serotonin influences transepithelial resistance only when added at the basolateral membrane,Citation58 and directly decreases expression of ZO-1 and ZO-2.Citation27 It exerts biphasic effects on the mammary tight junction, promoting tight junction integrity at low concentrations via protein kinase A; while sustained exposure to higher concentrations of serotonin disrupts tight junctions via p38 MAP kinase signaling, encouraging mammary involution.Citation45,Citation58 In addition, serotonin indirectly affects mammary tight junctions by stimulating prolactin secretion,Citation59 and is itself influenced by a prolactin-induced positive feedback mechanism.Citation57
Pathobiology of Mammary Ductal Barriers – Inflammation
As in many other organ systems, inflammation affecting the breast (mastitis), which most commonly occurs in the lactating gland, causes increased permeability of the ductal epithelium. This is evidenced by increased sodium and chloride content in the milk, as well as loss of transepithelial electrical resistance.Citation39,Citation60 While this may in part be due to direct epithelial injury, it has been shown in a variety of human and animal tissues that tight junction permeability is also increased as part of the inflammatory response, mediated by inflammatory cytokines such as tumor necrosis factor (TNF),Citation61,Citation62 histamineCitation63,Citation64 and interferon-γ in rat intestine.Citation65,Citation66 While these mechanisms have yet to be demonstrated in mammary tissue, it is quite likely that they are also involved in this process and may represent an important adaptive response to allow access by immune cells.
Pathobiology of Mammary Ductal Barriers – Breast Cancer
While some of the roles of ductal barriers in normal breast physiology and benign conditions have been described above, it is the role of mammary epithelial junctions and their proteins in the pathophysiology of breast cancer that has attracted by far the most attention. We will discuss below the role of ductal barriers and their constituent proteins in the pathophysiology of breast cancer, with particular focus on core markers of neoplastic behavior such as dedifferentiation, proliferation, migration, invasion and metastasis; before discussing the potential roles of junctional proteins as tumor biomarkers and drug targets.
Cell Polarity and Dedifferentiation
Tight junctions are vital to maintaining polarity of epithelial cells, delimiting their apical and basolateral aspects via the assembly of three complexes that maintain cell polarity: CRB, PAR and Scribble.
The CRB complex, the most apically-located, includes the proteins CRB3, PALS1 and PATJ and defines the apical region of polarized epithelial cells. CRB3 (Crumbs3) has been recognized as an important regulator of tight junction formation, its overexpression delaying tight junction formation in MDCK cells, the canine kidney cell line classically used as a highly-differentiated epithelium with well-developed junctional structures.Citation67 Forced expression of CRB3 in MCF10A cells, a mammary epithelial cell line that expresses little CRB3 and does not form true tight junctions in culture, has been shown to promote recruitment of ZO1, occludin and claudin-1 to sites of cell-cell contact and to induce the formation of true tight junctions.Citation68 PATJ stabilizes the CRB complex, having been demonstrated as vital for the proper localization of CRB3, as well as claudin-1, ZO-1, ZO-3, occludin and atypical PKC in mammalian epithelial tight junctions.Citation69-Citation71
The PAR complex consists of Par3, Par6, atypical PKC (aPKC) and Cdc42/Rac1. Par3 interacts with JAM-ACitation72 and PTEN,Citation73 which in turn allows it to interact with TIAM-1, stabilizing the junction.Citation74 Further binding of Cdc42/Rac1 recruits aPKC to the apical surface, maintaining the integrity of the apical cellular region.Citation75
The basolateral Scribble complex consists of the proteins Scribble, lethal giant larvae homolog (LGL) and discs large homolog (DLG).Citation76,Citation77 Scribble has been shown to co-localize with both the tight junction protein ZO-1Citation78 and with E-cadherin and DLG at adherens junctions,Citation79 suggesting its importance in the formation of both these junctions. Loss of Scribble function in Drosophila disrupts the localization of adherens junctions and apical proteinsCitation80 while its knockdown in MDCK cells disrupts adhesion, delaying tight junction formation and allowing more rapid migration without impairing cell polarity.Citation81
Loss of cell polarity is a crucial step in epithelial-mesenchymal transformation (EMT), the process whereby epithelial-derived cancers, including breast, progressively develop an invasive mesenchymal signature and phenotype.Citation82 Many proteins involved in cell polarity are dysregulated in breast cancer. The transcription factor ZEB1, which is upregulated in some breast cancers and is implicated in EMT, inhibits the expression of CRB3 and PATJ.Citation83 Overexpression of Par6 in breast epithelial cells induces increased proliferation while maintaining cell polarity,Citation84 and activation of ErbB2 (the gene encoding HER2, the oncogenic receptor tyrosine kinase overexpressed in a population of aggressive breast cancers) disrupts apical-basal polarity by associating with Par6 and aPKC.Citation85 Scribble expression invasive lobular carcinoma sp.ecimens has been shown to be quantitatively reduced,Citation79 while it is redistributed from the membrane to the cytoplasm in several invasive ductal carcinoma lines.Citation86 In addition, its loss in mammary epithelial cells results in abnormal morphogenesis both in vivo and in vitro and inhibits c-myc-induced apoptosis in vitro.Citation86
It is tempting to sp.eculate that alterations in the expression levels of tight junction proteins, or indeed their inappropriate localization, may also play a role in breast cellular dedifferentiation. Histological observations of deficits in the strict polarization of the breast ducts are frequently one of the earliest indicators of malignancy. Occludin is downregulated in breast cancer, and its forced expression promotes senescence in murine mammary carcinoma cells,Citation87 while expression of its interacting protein ZO-1 is reduced in poorly differentiated breast cancer sp.ecimens.Citation88 JAM-A plays an important role in regulating cell morphology by modulating activity of the integrin-activating small GTPase Rap1.Citation89 Despite initial conflicts between reports of its expressional correlation with the malignant breast phenotype,Citation90,Citation91 the balance of evidence from our laboratory and others now favors a model whereby JAM-A overexpression in breast cancer associates with poor prognosis.Citation91-Citation94 This will be further discussed in the Migration and Invasion section.
The cadherin switch is an important precursor of EMT in breast cancer. It involves a progressive dedifferentiation, switching from expression primarily of epithelial markers such as E-cadherin and cytokeratins to mesenchymal markers such as N-cadherin, vimentin and fibronectin.Citation95 This may be a normal component of processes such as wound healing and development of structures such as tubules,Citation96 however it occurs in a dysregulated fashion in cancer. E-cadherin is underexpressed in a number of breast cancer cell lines including the highly-invasive triple negative MDA-MB 231, although E- and N-cadherin status is not fully predictive of invasiveness.Citation97 Immunohistochemical staining of breast cancer sp.ecimens suggests that E-cadherin is lost in the majority of lobular, but not ductal, breast carcinoma,Citation98-Citation100 and occurs as early as the in situ stage in lobular carcinoma.Citation101 Numerous mechanisms can result in the loss or downregulation of E-cadherin, of which loss of heterozygosity and mutations in the CDH1 gene which encodes it is commonly seen in lobular carcinoma.Citation102,Citation103
A number of transcription factors can coordinate the shift from E- to N-cadherin expression, largely controlled by the transcription suppressor Snail1. The latter protein activates Zeb1, which binds to the E-cadherin promoter and blocks transcription of E-cadherin.Citation104,Citation105 Further repressors that have been implicated include Slug, Twist, Zeb2, E12/47, SIP1 and δEF1, in addition to hypermethylation of E-cadherin promoters.Citation106-Citation109 Furthermore, E-cadherin can be targeted for endocytosis and degradation secondary to the actions of tyrosine kinases such as Src, EGFR, FGF receptor, c-Met and IGF-1R in epithelial cellsCitation110; and can be directly degraded by matrix metalloproteinases.Citation95
The protein Twist, itself regulated by canonical Wnt1 signaling,Citation111 downregulates E-cadherin expression, concurrently upregulates N-cadherin expression and can induce EMT. Twist overexpression is associated with dysregulated cell growth in murine mammary tumors,Citation111 and with multi drug resistance in human breast cancer cells.Citation112,Citation113 N-cadherin promotes fibroblast growth factor (FGF) signaling in breast cancer cells by binding to and preventing the internalization of its receptor, thus sustaining its pro-migratory and invasive effects via MAP kinase activation and matrix metalloproteinase 9 secretion.Citation114
Dysregulation of Proliferation
Cancer can essentially be considered a disease of dysregulated cell growth and proliferation. A crucial point is the loss of regulation of the cell cycle, resulting in uncontrolled cell division and abnormal growth.Citation115 Loss of E-cadherin expression, in addition to facilitating cell detachment through its mechanical effect at adherens junctions, directly induces a number of pro-proliferative signaling pathways in breast cancer. E-cadherin interacts with the epidermal growth factor (EGF) receptor to modulate proliferation by suppressing pro-proliferative tyrosine kinase signaling.Citation116 E-cadherin also binds and sequesters β-catenin in adherens junctions, thus its downregulation frees β-catenin to enter the nucleus and participate in pro-proliferative canonical Wnt signaling. Increased levels of cytosolic and nuclear β-catenin have been reported in up to 68% of breast cancer patient sp.ecimens (77% of invasive lobular carcinoma, 64% of invasive ductal carcinoma), and implies a poor prognosis.Citation117-Citation120 In addition, upregulation of other β-catenin signaling promoters such as Disheveled,Citation121 LRP 6Citation122 and a mutant LRP5,Citation123 as well as downregulation of the β-catenin inhibitor Wnt5a,Citation124 are commonly seen in breast cancer sp.ecimens. Several alternative pathways known to increase β-catenin expression are activated in breast cancer, including the NF-κB pathway in ER-negative, HER2-positive tumorsCitation125; Pin1 upregulation in breast cancer patient sp.ecimens is proportional to increasing tumor grade and associated with increased β-catenin expressionCitation126,Citation127; while loss of PTEN activates Akt and β-catenin resulting in increased proliferation in breast cells.Citation128,Citation129
Placental (P-) cadherin, a junctional protein usually expressed by mammary myoepithelial cells, is expressed in approx 30% of breast cancer cell linesCitation97 and up to 50% of invasive ductal carcinoma sp.ecimens.Citation130 It is strongly expressed in the basal (classically ER, PR and HER2 triple negative) and HER2-overexpressing subtypes of breast cancer on patient tissue microarrays,Citation131 and has been suggested as a routine biomarker for basal-like cancers.Citation132,Citation133 It is associated with increased proliferation, a reduction in ER-α signaling, increased p53 and HER2 expression; and a poorer prognosis.Citation131 The cadherin switch from E- to P- expression is described in embryonic development,Citation133 with little evidence of its occurrence in breast carcinoma, where P-cadherin is more commonly co-expressed with E-cadherin in both human and murine studies.Citation134,Citation135
Although the bulk of mechanistic work implicating dysregulated adherens junction signaling in cancer has been performed in cell line models, concordance between cell line and animal data appears to be high and thus valuable insights from a wealth of genetic ablation studies in non-breast tissues must not be overlooked. While conventional knockout animals featuring germline loss of sp.ecific adherens junction proteins are often embryonic-lethal, recent advances in tissue-targeted loss of components such as p120-cateninCitation136,Citation137 have revealed interesting tendencies toward an increased risk of cancer. The precise contribution of altered adhesion vs. altered signaling to such tumorigenic events remains elusive, however contrived separation of these two functions may be unphysiological and unrealistic. As a comprehensive discussion of genetic models of adherens junction perturbation is beyond the scope of this article, interested readers are directed to a recent review on the topic.Citation138
Migration and Invasion
A further hallmark of malignancy that facilitates tumor sp.read and thus survival is dysregulated migration and invasion. Broadly sp.eaking, cell migration consists of five cyclical steps, reviewed in.Citation139 It begins with the formation of protrusions known as pseudopodia at the leading edge, driven by actin polymerization controlled by the Rho GTPase Cdc42 and several downstream effectors. Small transient adhesions to extracellular matrix near the leading edge are formed by β1, β2 and α2β1 integrins and other adaptor and signaling proteins, interacting with actin; and these adhesions further develop in response to tension applied by stress fibers. Proteinases are recruited to these focal adhesions, which then cleave extracellular matrix barriers.Citation140,Citation141 The cell body translocates forward driven by myosin bundles sliding along actin filaments in an energy-dependent process.Citation139 Release of adhesions at the rear of the cell and retraction of the rear complete the cycle.Citation142 Normal epithelium and well-differentiated carcinomas tend to exhibit collective migration and invasion, whereby cell-cell interactions are retained and migration occurs in single sheets or strands.Citation143 In contrast, inflammatory cells, mesenchymally-derived tumor cells and poorly-differentiated carcinomas with loss of strong cell-cell contacts tend to migrate individually.Citation144
The tight junctional protein JAM-A, a Type I transmembrane protein belonging to the immunoglobulin superfamily and a known regulator of cell adhesion and cell migration (reviewed inCitation145), has a somewhat controversial role in breast cancer. While early evidence suggested that low JAM-A expression correlated with a less migratory and invasive breast cancer phenotype,Citation90 an increasing body of evidence from our group and others would suggest otherwise. Specifically, JAM-A overexpression in breast cancer sp.ecimens correlates with poorer patient prognosis,Citation91 and its expression has been shown to correlate with HER2 expression,Citation92 ER negativity, higher grade, and aggressive luminal B, HER2 and basal subtypes of breast cancer. Knockdown or antagonism of JAM-A reduces migration and invasion in JAM-A-expressing breast cancer cells,Citation146 and JAM-A has been shown to act as an upstream regulator of various signaling pathways relevant to the promotion of migratory behavior. One such pathway has revealed a direct relationship between JAM-A expression and that of the migratory protein β1-integrin in both colonicCitation89 and breastCitation91,Citation146 epithelial cells, and accordingly JAM-A dimerization signaling has been shown to regulate expression levels and activation status of the β1-integrin-activating small GTPase Rap1.Citation89,Citation91,Citation146 A second emerging pathway implicates JAM-A as a novel regulator of the expression levels (and therefore signaling potential) of oncogenic HER2 in breast cancer cell lines, via a mechanism regulating the proteasomal stability of HER2.Citation92 It is tempting to sp.eculate that JAM-A might also regulate the signaling functions of other oncogenes or receptor tyrosine kinases by influencing their proteasomal stability, but as yet this field of investigation is in its infancy. Regarding the initial controversy over whether or not JAM-A expression levels have a positive or a negative correlation with aggressive and migratory tumor behavior,Citation90,Citation91 the balance of studies supports the notion of JAM-A as a positive regulator of cancer progression at least in the breast.Citation94 Nonetheless it is possible that seemingly contradictory effects could be explained by JAM-A under-expression impairing cell adhesion and polarity, favoring early malignant change; while its overexpression in later stages of cancer might favor tumor progression via (among others) integrin-mediated pro-migratory signaling.Citation2
Another key family of tight junction molecules is the 27-member claudin family of membrane tetra-sp.anning proteins (reviewed inCitation147). Although originally best known for ultrastructurally driving tight junction strand formation and consequently exerting differential control over charge selectivity through various epithelia (reviewed inCitation148), claudins have recently also been implicated in the control of cell migration and reported to be up- or downregulated in a variety of cancers. Claudin 4 appears to regulate migration in both normal and malignant breast cells,Citation149 and its expression correlates with higher grade and worse prognosis in breast cancer sp.ecimens.Citation150 Similarly, claudin 5 expression in breast tumor sp.ecimens correlates with worse survival and its forced expression increases breast cancer cell motility in vitro.Citation151 In contrast, forced overexpression of claudins -6Citation152 and -16Citation153 have been shown to decrease breast cancer cell migration and invasion in vitro, and loss of claudin 6 conversely promotes anchorage-independent survival of MCF 7 breast cancer cells.Citation154 Claudin 1 under-expression has been demonstrated in breast cancer sp.ecimens,Citation155 while conversely high levels in breast cancers have been shown to correlate with the aggressive basal-like phenotype.Citation156 Interestingly, a new molecular subtype of breast cancer, “claudin low” claudin low, has been recently described, in which tumors are characterized by low gene expression of claudins 3,4 and 7 in conjunction with an aggressive, basal-like phenotype.Citation157 However there is a paucity of information on potential cross-talk mechanisms between claudins and established drivers of tumorigenicity. Intriguing recent data have demonstrated a physical interaction between claudin-7 and the potential oncogene EpCAM in various gastrointestinal cells, tissues and tumors,Citation158,Citation159 while emerging evidence suggests that EpCAM regulates the lysosomal degradation of claudin-7 in addition to its localization.Citation160 It is likely that pursuit of knowledge regarding the mechanistic involvement of various claudins in cancer cell migration and tumor metastasis will be a lucrative area of research in coming years.
Adherens junctional proteins are also strongly implicated in breast cancer migration and invasion. The cadherin switch to expression of mesenchymal cadherins most likely facilitates migration and invasion both by increasing tumor cells’ ability to detach from their normal surrounding epithelial cells, and by inducing inappropriate pro-motility signaling.Citation161 Transfection of N-cadherin into E-cadherin-expressing breast cancer cells induces invasion and motility,Citation97 while transfection of E-cadherin into highly invasive mesenchymal-like MDA-MB 231 cells reduces invasion and migration.Citation162
Metastasis
Migration and invasion are particularly important for the systemic sp.read of tumors, acting as a key early step in the multi-step cascade known as metastasis. Accordingly, cell junctions and their proteins have been reported to play an intrinsic role in preventing breast cancer metastasis. The primary event in metastasis involves detachment of cells from the primary tumor and invasion into the bloodstream, followed by extravasation at the site of metastasis. This is somewhat similar to extravasation of leukocytes in the immune response and consists primarily of three steps; first loose attachment and rolling on the endothelial surface, second tighter attachment of the tumor cells to the endothelium, and third diapedesis or transmigration through the endothelium. The latter can occur either by the transcellular or paracellular route. While the loose attachment, rolling and tighter attachment steps in tumor cells are similar to leukocytes, diapedesis transmigration of tumor cells differs from that of leukocytes (termed diapedesis) in that it permanently alters endothelial morphology,Citation163 resulting in retraction of endothelial cells and in some cases apoptosis, possibly due to loss of cell-cell contacts.Citation164-Citation166 N-cadherin interactions between tumor and vascular endothelial cells appear to partly mediate tumor cell-endothelial attachment and extravasation.Citation167,Citation168
Claudin-2 expression has been shown to be increased in breast cancers that metastasize to the liver. Its ability to mediate tumor cell- hepatocyte interaction is thought to facilitate arrest in this organ.Citation169,Citation170 A further study in mice has reported downregulation of claudin 4, claudin 7 and γ-catenin in liver metastases originating from breast cell lines, in conjunction with altered γ-catenin cellular localization. Interestingly, claudin 7 was also expressed by macrophage-like cells surrounding the liver metastases, and was re-expressed in large tumors, suggesting a possible interaction of with the microenvironment to promote metastasis.Citation171
Breast cancer commonly metastasizes to brain, with a prevalence of approximately 30% at autopsy.Citation172,Citation173 Risk factors include high grade and stage, young age, estrogen receptor negativity and HER2 overexpression.Citation174,Citation175 Brain metastasis requires breach of the blood brain barrier (BBB), a unique non-fenestrated endothelial structure that prevents passage of large molecules and cells. Tight and adherens junctions are integral to the barrier function of brain microvascular endothelial cells (BMECs).Citation176,Citation177 BMECs display higher transepithelial electrical resistance and lower solute permeability than other endothelial cells, while their tight junctions are more complex and passage of polar solutes via the paracellular pathway is greatly reduced. The basement membrane is relatively thicker, and the underlying astrocytes regulate flow across the barrier. Among the proteins implicated in susceptibility to brain metastasis, loss of claudins 3 and 5 is associated with increased leakiness of the BBB in vivo.Citation178,Citation179 Stromal cell derived factor-1α (SDF-1 α) is a chemokine expressed by several organs including the CNS, and expression of its receptor CXCR4 in breast cancer cells may facilitate BBB penetration. SDF-1 α treatment increases permeability of BMEC monolayers to breast tumor cell invasion, activating the PI-3K/AKT signaling pathway and causing endothelial cell retraction.Citation180 HER2/Neu has been shown to upregulate CXCR4 expression.Citation181 Matrix metalloproteinases also play a role in breast to brain metastasis; with MMP-2 and -3 activity increased in vivoCitation182,Citation183; and that of MMP-1 and -9 increased in vitro.Citation184
Having discussed many of the functional neoplastic processes relevant to breast cancer progression involving junctions, we will now examine some of the many proteins and membrane domains that have been implicated in junction-based signaling in breast cancer. As an exhaustive examination of all implicated proteins and domains would be beyond the scope of this short review, we will focus on a select few relevant to our own research.
Lipid Rafts and Adhesion Proteins
While the classical model of the plasma membrane describes the membrane as a liquid-disordered phospholipid bilayer with molecules such as proteins and cholesterol randomly interspersed,Citation185 this model is now viewed as an oversimplification. Specifically it has been recognized that cholesterol and several proteins involved in dynamic cellular processes are non-randomly segregated into liquid-ordered membrane microdomains known as lipid rafts, which form as a consequence of tight sp.atial packing of membrane domains predominantly composed of sp.hingolipids, as opposed to phospholipids. This was initially postulated based on such observations as the ability of differing phases to co-exist in lipid bilayersCitation186,Citation187; differential distribution and clustering of membrane lipidsCitation188-Citation190; and the presence of sp.hingolipid-enriched detergent-resistant regions of cell membranes.Citation191 Simons et al. were among the first to define the concept of lipid rafts,Citation192 which have been defined as “small (10–200 nm), heterogeneous, highly dynamic, sterol- and sphingolipid-enriched domains that compartmentalize cellular processes”Citation193.
Several adhesion proteins have been described to be associated with lipid raft domains, and it has even been suggested that tight junctions themselves constitute a subtype of raft domain.Citation194 Biochemical enrichment of occludin and ZO-1 have been demonstrated in detergent-resistant domains in humanCitation194,Citation195 intestinal cells, while claudins 1, 3, 4, 5, 7, 8Citation195-Citation197 and JAM-ACitation197 have been retrieved from detergent-resistant domains in various epithelial cells. Cholesterol depletion with methyl β cyclodextrin has been shown to redistribute claudins 3, 4 and 7, JAM-A and occludin, but not claudin 1, out of these domains in intestinal epithelial cells.Citation197 The tyrosine kinases Src and EGFR both localize in breast cancer cell lipid raftsCitation198-Citation200; both regulate cadherin/catenin interactions,Citation201,Citation202 and both have been strongly implicated in pro-malignant signaling in breast cancer.Citation202,Citation203 An inhibitor sp.ecifically targeting raft-associated Src has been shown to inhibit cell cycle progression and cell adhesion in breast cancer cells, while the absence of a raft-targeting sequence in the inhibitor eliminated these effects.Citation200 Pharmacological disruption of lipid rafts using lovastatin to interfere with cholesterol biosynthesis increases the growth inhibitory effects of the EGFR inhibitor gefitinib in breast cancer cell lines that normally are resistant to this agent.Citation199 In addition, components of the Wnt signaling pathway may be lipid raft associated.Citation204,Citation205 It has been suggested that cholesterol in lipid rafts stabilizes the protein complexes in tight junction strands.Citation197
While junctions play an important role in cell migration, it has also been shown via a pharmacological raft-disruption approach that lipid rafts modulate front-rear polarity in migrating MCF7 breast cancer cells.Citation206 Work from our group has revealed shuttling of the hyaluronan receptor CD44 in and out of intact lipid raft domains of breast epithelial cells according to migratory status,Citation207 and recent site-directed mutagenesis work has revealed that genetic targeting of biochemical motifs which drive raft affiliation of CD44 is sufficient to force an EMT-like state in breast cancer cells (Babina, Donatello, Hill and Hopkins, manuscript under review). Similarly, unpublished work from our group suggests shuttling of Na+ K+ ATPase in and out of raft domains in breast cancer cells lines, in a hormone receptor status-dependent manner, in response to treatment with anti-proliferative doses of the potential anti-cancer drugs cardiac glycosides (Owens and Hopkins, manuscript in preparation). It is intriguing to sp.eculate that similar changes may occur in breast cancer with raft-associated junctional proteins in response to such junction-dependent processes as cell migration. However it is advisable to interpret the collective literature on lipid rafts in physiological/pathophysiological processes with caution due to a relative over-dependence on the pharmacological lipid raft disrupting tool, methyl-β-cyclodextrin, which has recognized limitations in terms of sp.ecificity.Citation208
Clinical Application of Adhesion Molecules in Breast Cancer
A number of tight and adherens junction proteins have been suggested or investigated as potential biomarkers in breast cancer. As discussed above, claudin expression in breast cancer has been closely examined, and the aggressive claudin-low subtype is now recognized, defined by low gene expression of claudins 3, 4 and 7.Citation157 Thus, claudin characterization will likely become a routine part of breast cancer diagnostic and prognostic workup.
P-cadherin has been identified as a cancer stem cell marker for basal-type breast cancer,Citation133,Citation209 and has been shown to be an independent marker for disease-free, but not overall, survival.Citation210 While the prominent role of E-cadherin downregulation in EMT would make it a tempting proposition as a prognostic indicator (and in fact E-cadherin loss has diagnostic value in lobular carcinomas), used alone its correlation with prognosis has been variable.Citation211-Citation216 One study found a reduction in one of E-cadherin, β-catenin, α-catenin and plakoglobin in tumor sp.ecimens to correlate significantly with breast cancer metastasis,Citation217 and a recent paper has indicated a combination of E-cadherin and carcinoembryonic antigen as a useful predictor of relapse.Citation216 As E-cadherin sequestration of β-catenin in adherens junctions prevents the latter partaking in pro-neoplastic canonical Wnt signaling, it would be logical that a measure of β-catenin distribution might be of prognostic benefit. One study found that a novel scoring system of membrane minus cytoplasmic β-catenin correlated with worse outcome in breast cancer.Citation218
Despite the obvious theoretical promise of cell junctions and their proteins as anti-metastatic therapeutic targets, junction-directed therapies are still an exciting and under-explored area. Perhaps the greatest potential lies with targeting Claudins 3 and 4, which have been recognized as the receptors for the permeability-enhancing lytic toxin Clostridium perfringens enterotoxin (CPE).Citation219 CPE may be a useful therapy in breast cancers overexpressing these proteins, as it has been shown to induce lysis of claudin 3- and 4- overexpressing breast cancer cell lines.Citation220
Another exciting potential target is JAM-A, given the positive association between its overexpression and poor prognosis in breast cancer patientsCitation91,Citation92,Citation94 and following a recent publication demonstrating anti-proliferative efficacy of a function-blocking JAM-A antibody in xenograft murine models of breast cancer.Citation221 Unpublished work from our group has also shown promising in vitro and pre-clinical in vivo efficacy of a novel small molecule inhibitor of JAM-A, which we sp.eculate could be particularly valuable in aggressive breast cancers concomitantly overexpressing HER2 and JAM-A.
ADH-1, an anti-N cadherin protein, has shown efficacy against pancreatic and prostate cancer in preclinical studies,Citation222,Citation223 in addition to promising effects on disease stabilization in early clinical trials.Citation224 However, it has yet to be evaluated in breast cancer.
While the targeting of junctional proteins in breast cancer is still in its infancy, the expanding roles of these proteins in driving malignant signaling processes suggest many exciting targets for future research.
Conclusion
It is clear that breast ductal adhesion complexes and their constituent proteins play vital roles in breast physiology and pathology, not just by influencing mechanical adhesion and stability but also by influencing key cell signaling and gene transcription events. While modulation of the physical properties of breast ductal barriers is essential for cyclical changes in lactation and engorgement, we have also discussed the role of junctional proteins in changes such as ductal development in embryogenesis and puberty. We have further examined how alterations in junctional integrity could potentially contribute to pathologies including breast inflammation and breast cancer invasion/metastasis. We have particularly focused upon the role of junctional proteins in dysregulated signal transduction and gene transcription events that are associated with neoplastic phenomena such as proliferation, dedifferentiation, invasion and metastasis. Finally, we have explored some of the many exciting prospects for junctional proteins as both prognostic biomarkers and as therapeutic targets. It is the latter function of junctional proteins that is currently the focus of much research, and that may yield meaningful contributions to patient care in the future.
Acknowledgments
The authors wish to thank Dr. Kieran Brennan for critical reading of the manuscript and for helpful discussions. AMH is grateful to Science Foundation Ireland, the Health Research Board of Ireland, the Beaumont Hospital Cancer Research and Development Trust, Breast Cancer Ireland and Cancer Research Ireland for past or current support of relevant studies from her laboratory.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Ferlay J, Shin H, Bray F, Forman D, Mathers C, Parkin D. GLOBOCAN 2008 v1.2, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 10 [Internet]. 2010 [cited; Available from: http://globocan.iarc.fr
- Brennan K, Offiah G, McSherry EA, Hopkins AM. Tight junctions: a barrier to the initiation and progression of breast cancer?. J Biomed Biotechnol 2010; 2010:460607; http://dx.doi.org/10.1155/2010/460607; PMID: 19920867
- Ramsay DT, Kent JC, Hartmann RA, Hartmann PE. Anatomy of the lactating human breast redefined with ultrasound imaging. J Anat 2005; 206:525 - 34; http://dx.doi.org/10.1111/j.1469-7580.2005.00417.x; PMID: 15960763
- Guinebretière JM, Menet E, Tardivon A, Cherel P, Vanel D. Normal and pathological breast, the histological basis. Eur J Radiol 2005; 54:6 - 14; http://dx.doi.org/10.1016/j.ejrad.2004.11.020; PMID: 15797289
- Schneeberger EE, Lynch RD. The tight junction: a multifunctional complex. Am J Physiol Cell Physiol 2004; 286:C1213 - 28; http://dx.doi.org/10.1152/ajpcell.00558.2003; PMID: 15151915
- Green KJ, Getsios S, Troyanovsky S, Godsel LM. Intercellular junction assembly, dynamics, and homeostasis. Cold Spring Harb Perspect Biol 2010; 2:a000125; http://dx.doi.org/10.1101/cshperspect.a000125; PMID: 20182611
- Meng W, Takeichi M. Adherens junction: molecular architecture and regulation. Cold Spring Harb Perspect Biol 2009; 1:a002899; http://dx.doi.org/10.1101/cshperspect.a002899; PMID: 20457565
- Brooke MA, Nitoiu D, Kelsell DP. Cell-cell connectivity: desmosomes and disease. J Pathol 2012; 226:158 - 71; http://dx.doi.org/10.1002/path.3027; PMID: 21989576
- Basham KJ, Kieffer C, Shelton DN, Leonard CJ, Bhonde VR, Vankayalapati H, Milash B, Bearss DJ, Looper RE, Welm BE. Chemical genetic screen reveals a role for desmosomal adhesion in mammary branching morphogenesis. J Biol Chem 2013; 288:2261 - 70; http://dx.doi.org/10.1074/jbc.M112.411033; PMID: 23212921
- Pitelka DR, Hamamoto ST, Duafala JG, Nemanic MK. Cell contacts in the mouse mammary gland. I. Normal gland in postnatal development and the secretory cycle. J Cell Biol 1973; 56:797 - 818; http://dx.doi.org/10.1083/jcb.56.3.797; PMID: 4569313
- Warburton MJ, Mitchell D, Ormerod EJ, Rudland P. Distribution of myoepithelial cells and basement membrane proteins in the resting, pregnant, lactating, and involuting rat mammary gland. J Histochem Cytochem 1982; 30:667 - 76; http://dx.doi.org/10.1177/30.7.6179984; PMID: 6179984
- Davies EL, Gee JM, Cochrane RA, Jiang WG, Sharma AK, Nicholson RI, Mansel RE. The immunohistochemical expression of desmoplakin and its role in vivo in the progression and metastasis of breast cancer. Eur J Cancer 1999; 35:902 - 7; http://dx.doi.org/10.1016/S0959-8049(99)00031-3; PMID: 10533469
- Maynadier M, Chambon M, Basile I, Gleizes M, Nirde P, Gary-Bobo M, Garcia M. Estrogens promote cell-cell adhesion of normal and malignant mammary cells through increased desmosome formation. Mol Cell Endocrinol 2012; 364:126 - 33; http://dx.doi.org/10.1016/j.mce.2012.08.016; PMID: 22963885
- Lu M, Mira-y-Lopez R, Nakajo S, Nakaya K, Jing Y. Expression of estrogen receptor alpha, retinoic acid receptor alpha and cellular retinoic acid binding protein II genes is coordinately regulated in human breast cancer cells. Oncogene 2005; 24:4362 - 9; http://dx.doi.org/10.1038/sj.onc.1208661; PMID: 15870697
- Zhu WY, Jones CS, Amin S, Matsukuma K, Haque M, Vuligonda V, Chandraratna RA, De Luca LM. Retinoic acid increases tyrosine phosphorylation of focal adhesion kinase and paxillin in MCF-7 human breast cancer cells. Cancer Res 1999; 59:85 - 90; PMID: 9892191
- Lacroix M, Leclercq G. Relevance of breast cancer cell lines as models for breast tumours: an update. Breast Cancer Res Treat 2004; 83:249 - 89; http://dx.doi.org/10.1023/B:BREA.0000014042.54925.cc; PMID: 14758095
- MacLeod RA, Dirks WG, Matsuo Y, Kaufmann M, Milch H, Drexler HG. Widespread intraspecies cross-contamination of human tumor cell lines arising at source. Int J Cancer 1999; 83:555 - 63; http://dx.doi.org/10.1002/(SICI)1097-0215(19991112)83:4<555::AID-IJC19>3.0.CO;2-2; PMID: 10508494
- Borowsky AD. Choosing a mouse model: experimental biology in context--the utility and limitations of mouse models of breast cancer. Cold Spring Harb Perspect Biol 2011; 3:a009670; http://dx.doi.org/10.1101/cshperspect.a009670; PMID: 21646376
- Bargmann CI, Hung MC, Weinberg RA. The neu oncogene encodes an epidermal growth factor receptor-related protein. Nature 1986; 319:226 - 30; http://dx.doi.org/10.1038/319226a0; PMID: 3945311
- Kari FW, Weaver R, Neville MC. Active transport of nitrofurantoin across the mammary epithelium in vivo. J Pharmacol Exp Ther 1997; 280:664 - 8; PMID: 9023277
- Monks J, Neville MC. Albumin transcytosis across the epithelium of the lactating mouse mammary gland. J Physiol 2004; 560:267 - 80; http://dx.doi.org/10.1113/jphysiol.2004.068403; PMID: 15297572
- Nguyen DA, Parlow AF, Neville MC. Hormonal regulation of tight junction closure in the mouse mammary epithelium during the transition from pregnancy to lactation. J Endocrinol 2001; 170:347 - 56; http://dx.doi.org/10.1677/joe.0.1700347; PMID: 11479131
- Ussing HH. Transport through biological membranes. Annu Rev Physiol 1953; 15:1 - 20; http://dx.doi.org/10.1146/annurev.ph.15.030153.000245; PMID: 13125285
- Lee SY, Palmer ML, Maniak PJ, Jang SH, Ryu PD, O’Grady SM. P2Y receptor regulation of sodium transport in human mammary epithelial cells. Am J Physiol Cell Physiol 2007; 293:C1472 - 80; http://dx.doi.org/10.1152/ajpcell.00068.2007; PMID: 17715387
- Quesnell RR, Han X, Schultz BD. Glucocorticoids stimulate ENaC upregulation in bovine mammary epithelium. Am J Physiol Cell Physiol 2007; 292:C1739 - 45; http://dx.doi.org/10.1152/ajpcell.00369.2006; PMID: 17251323
- Bisbee CA, Machen TE, Bern HA. Mouse mammary epithelial cells on floating collagen gels: transepithelial ion transport and effects of prolactin. Proc Natl Acad Sci U S A 1979; 76:536 - 40; http://dx.doi.org/10.1073/pnas.76.1.536; PMID: 284373
- Marshall AM, Pai VP, Sartor MA, Horseman ND. In vitro multipotent differentiation and barrier function of a human mammary epithelium. Cell Tissue Res 2009; 335:383 - 95; http://dx.doi.org/10.1007/s00441-008-0719-0; PMID: 19005683
- Incassati A, Chandramouli A, Eelkema R, Cowin P. Key signaling nodes in mammary gland development and cancer: β-catenin. Breast Cancer Res 2010; 12:213; http://dx.doi.org/10.1186/bcr2723; PMID: 21067528
- Chu EY, Hens J, Andl T, Kairo A, Yamaguchi TP, Brisken C, Glick A, Wysolmerski JJ, Millar SE. Canonical WNT signaling promotes mammary placode development and is essential for initiation of mammary gland morphogenesis. Development 2004; 131:4819 - 29; http://dx.doi.org/10.1242/dev.01347; PMID: 15342465
- Hens J, Dann P, Hiremath M, Pan TC, Chodosh L, Wysolmerski J. Analysis of gene expression in PTHrP-/- mammary buds supports a role for BMP signaling and MMP2 in the initiation of ductal morphogenesis. Dev Dyn 2009; 238:2713 - 24; http://dx.doi.org/10.1002/dvdy.22097; PMID: 19795511
- Zeng YA, Nusse R. Wnt proteins are self-renewal factors for mammary stem cells and promote their long-term expansion in culture. Cell Stem Cell 2010; 6:568 - 77; http://dx.doi.org/10.1016/j.stem.2010.03.020; PMID: 20569694
- Weber-Hall SJ, Phippard DJ, Niemeyer CC, Dale TC. Developmental and hormonal regulation of Wnt gene expression in the mouse mammary gland. Differentiation 1994; 57:205 - 14; http://dx.doi.org/10.1046/j.1432-0436.1994.5730205.x; PMID: 7988795
- Lane TF, Leder P. Wnt-10b directs hypermorphic development and transformation in mammary glands of male and female mice. Oncogene 1997; 15:2133 - 44; http://dx.doi.org/10.1038/sj.onc.1201593; PMID: 9393971
- Brisken C, Heineman A, Chavarria T, Elenbaas B, Tan J, Dey SK, McMahon JA, McMahon AP, Weinberg RA. Essential function of Wnt-4 in mammary gland development downstream of progesterone signaling. Genes Dev 2000; 14:650 - 4; PMID: 10733525
- Linzell JL, Peaker M. Changes in colostrum composition and in the permeability of the mammary epithelium at about the time of parturition in the goat. J Physiol 1974; 243:129 - 51; PMID: 4449059
- Linzell JL, Peaker M. The permeability of mammary ducts. J Physiol 1971; 216:701 - 16; PMID: 5105749
- Berga SE. Electrical potentials and cell-to-cell dye movement in mouse mammary gland during lactation. Am J Physiol 1984; 247:C20 - 5; PMID: 6742181
- Peaker M. Mechanism of milk secretion: milk composition in relation to potential difference across the mammary epithelium. J Physiol 1977; 270:489 - 505; PMID: 903903
- Nguyen DA, Neville MC. Tight junction regulation in the mammary gland. J Mammary Gland Biol Neoplasia 1998; 3:233 - 46; http://dx.doi.org/10.1023/A:1018707309361; PMID: 10819511
- Neville M. Determinants of milk volume and composition. A. Lactogenesis in women: A cascade of events revealed by milk composition. In: Jensen R, ed. Handbook of Milk Composition. San Diego: Academic Press:87-98.
- Morgan G, Wooding FB. A freeze-fracture study of tight junction structure in sheep mammary gland epithelium during pregnancy and lactation. J Dairy Res 1982; 49:1 - 11; http://dx.doi.org/10.1017/S002202990002207X; PMID: 7076943
- Itoh M, Bissell MJ. The organization of tight junctions in epithelia: implications for mammary gland biology and breast tumorigenesis. J Mammary Gland Biol Neoplasia 2003; 8:449 - 62; http://dx.doi.org/10.1023/B:JOMG.0000017431.45314.07; PMID: 14985640
- Stelwagen K, van Espen DC, Verkerk GA, McFadden HA, Farr VC. Elevated plasma cortisol reduces permeability of mammary tight junctions in the lactating bovine mammary epithelium. J Endocrinol 1998; 159:173 - 8; http://dx.doi.org/10.1677/joe.0.1590173; PMID: 9795355
- Stelwagen K, McFadden HA, Demmer J. Prolactin, alone or in combination with glucocorticoids, enhances tight junction formation and expression of the tight junction protein occludin in mammary cells. Mol Cell Endocrinol 1999; 156:55 - 61; http://dx.doi.org/10.1016/S0303-7207(99)00145-8; PMID: 10612423
- Pai VP, Horseman ND. Biphasic regulation of mammary epithelial resistance by serotonin through activation of multiple pathways. J Biol Chem 2008; 283:30901 - 10; http://dx.doi.org/10.1074/jbc.M802476200; PMID: 18782769
- Kalantaridou SN, Makrigiannakis A, Zoumakis E, Chrousos GP. Reproductive functions of corticotropin-releasing hormone. Research and potential clinical utility of antalarmins (CRH receptor type 1 antagonists). Am J Reprod Immunol 2004; 51:269 - 74; http://dx.doi.org/10.1111/j.1600-0897.2004.00155.x; PMID: 15212679
- Burke CW, Roulet F. Increased exposure of tissues to cortisol in late pregnancy. Br Med J 1970; 1:657 - 9; http://dx.doi.org/10.1136/bmj.1.5697.657; PMID: 5443967
- Rubenstein NM, Chan JF, Kim JY, Hansen SH, Firestone GL. Rnd3/RhoE induces tight junction formation in mammary epithelial tumor cells. Exp Cell Res 2005; 305:74 - 82; http://dx.doi.org/10.1016/j.yexcr.2004.12.010; PMID: 15777789
- Zettl KS, Sjaastad MD, Riskin PM, Parry G, Machen TE, Firestone GL. Glucocorticoid-induced formation of tight junctions in mouse mammary epithelial cells in vitro. Proc Natl Acad Sci U S A 1992; 89:9069 - 73; http://dx.doi.org/10.1073/pnas.89.19.9069; PMID: 1409603
- Rubenstein NM, Guan Y, Woo PL, Firestone GL. Glucocorticoid down-regulation of RhoA is required for the steroid-induced organization of the junctional complex and tight junction formation in rat mammary epithelial tumor cells. J Biol Chem 2003; 278:10353 - 60; http://dx.doi.org/10.1074/jbc.M213121200; PMID: 12525486
- Failor KL, Desyatnikov Y, Finger LA, Firestone GL. Glucocorticoid-induced degradation of glycogen synthase kinase-3 protein is triggered by serum- and glucocorticoid-induced protein kinase and Akt signaling and controls beta-catenin dynamics and tight junction formation in mammary epithelial tumor cells. Mol Endocrinol 2007; 21:2403 - 15; http://dx.doi.org/10.1210/me.2007-0143; PMID: 17595317
- Feng Z, Marti A, Jehn B, Altermatt HJ, Chicaiza G, Jaggi R. Glucocorticoid and progesterone inhibit involution and programmed cell death in the mouse mammary gland. J Cell Biol 1995; 131:1095 - 103; http://dx.doi.org/10.1083/jcb.131.4.1095; PMID: 7490285
- Linzell JL, Peaker M, Taylor JC. The effects of prolactin and oxytocin on milk secretion and on the permeability of the mammary epithelium in the rabbit. J Physiol 1975; 253:547 - 63; PMID: 1214226
- Flint DJ, Gardner M. Evidence that growth hormone stimulates milk synthesis by direct action on the mammary gland and that prolactin exerts effects on milk secretion by maintenance of mammary deoxyribonucleic acid content and tight junction status. Endocrinology 1994; 135:1119 - 24; http://dx.doi.org/10.1210/en.135.3.1119; PMID: 8070355
- Sheffield L, Kotolski L. Prolactin inhibits programmed cell death during mammary gland involution. FASEB J 1992; 6:A1184
- Lin Y, Xia L, Turner JD, Zhao X. Morphologic observation of neutrophil diapedesis across bovine mammary gland epithelium in vitro. Am J Vet Res 1995; 56:203 - 7; PMID: 7717587
- Matsuda M, Imaoka T, Vomachka AJ, Gudelsky GA, Hou Z, Mistry M, Bailey JP, Nieport KM, Walther DJ, Bader M, et al. Serotonin regulates mammary gland development via an autocrine-paracrine loop. Dev Cell 2004; 6:193 - 203; http://dx.doi.org/10.1016/S1534-5807(04)00022-X; PMID: 14960274
- Stull MA, Pai V, Vomachka AJ, Marshall AM, Jacob GA, Horseman ND. Mammary gland homeostasis employs serotonergic regulation of epithelial tight junctions. Proc Natl Acad Sci U S A 2007; 104:16708 - 13; http://dx.doi.org/10.1073/pnas.0708136104; PMID: 17940054
- Balsa JA, Sánchez-Franco F, Pazos F, Lara JI, Lorenzo MJ, Maldonado G, Cacicedo L. Direct action of serotonin on prolactin, growth hormone, corticotropin and luteinizing hormone release in cocultures of anterior and posterior pituitary lobes: autocrine and/or paracrine action of vasoactive intestinal peptide. Neuroendocrinology 1998; 68:326 - 33; http://dx.doi.org/10.1159/000054381; PMID: 9822800
- Linzell JL, Peaker M. Day-to-day variations in milk composition in the goat and cow as a guide to the detection of subclinical mastitis. Br Vet J 1972; 128:284 - 95; PMID: 4672325
- Juric M, Xiao F, Amasheh S, May O, Wahl K, Bantel H, Manns MP, Seidler U, Bachmann O. Increased epithelial permeability is the primary cause for bicarbonate loss in inflamed murine colon. Inflamm Bowel Dis 2013; 19:904 - 11; http://dx.doi.org/10.1097/MIB.0b013e3182813322; PMID: 23502355
- Cui W, Li LX, Sun CM, Wen Y, Zhou Y, Dong YL, Liu P. Tumor necrosis factor alpha increases epithelial barrier permeability by disrupting tight junctions in Caco-2 cells. Braz J Med Biol Res 2010; 43:330 - 7; http://dx.doi.org/10.1590/S0100-879X2010007500020; PMID: 20445948
- Zhang Q, Fisher K. Tight junction-related barrier contributes to the electrophysiological asymmetry across vocal fold epithelium. PLoS One 2012; 7:e34017; http://dx.doi.org/10.1371/journal.pone.0034017; PMID: 22442739
- Flynn AN, Itani OA, Moninger TO, Welsh MJ. Acute regulation of tight junction ion selectivity in human airway epithelia. Proc Natl Acad Sci U S A 2009; 106:3591 - 6; http://dx.doi.org/10.1073/pnas.0813393106; PMID: 19208806
- Stepánková R, Kofronová O, Tucková L, Kozáková H, Cebra JJ, Tlaskalová- Hogenová H. Experimentally induced gluten enteropathy and protective effect of epidermal growth factor in artificially fed neonatal rats. J Pediatr Gastroenterol Nutr 2003; 36:96 - 104; http://dx.doi.org/10.1097/00005176-200301000-00018; PMID: 12500003
- Cenac N, Chin AC, Garcia-Villar R, Salvador-Cartier C, Ferrier L, Vergnolle N, Buret AG, Fioramonti J, Bueno L. PAR2 activation alters colonic paracellular permeability in mice via IFN-gamma-dependent and -independent pathways. J Physiol 2004; 558:913 - 25; http://dx.doi.org/10.1113/jphysiol.2004.061721; PMID: 15194744
- Lemmers C, Michel D, Lane-Guermonprez L, Delgrossi MH, Médina E, Arsanto JP, Le Bivic A. CRB3 binds directly to Par6 and regulates the morphogenesis of the tight junctions in mammalian epithelial cells. Mol Biol Cell 2004; 15:1324 - 33; http://dx.doi.org/10.1091/mbc.E03-04-0235; PMID: 14718572
- Fogg VC, Liu CJ, Margolis B. Multiple regions of Crumbs3 are required for tight junction formation in MCF10A cells. J Cell Sci 2005; 118:2859 - 69; http://dx.doi.org/10.1242/jcs.02412; PMID: 15976445
- Michel D, Arsanto JP, Massey-Harroche D, Béclin C, Wijnholds J, Le Bivic A. PATJ connects and stabilizes apical and lateral components of tight junctions in human intestinal cells. J Cell Sci 2005; 118:4049 - 57; http://dx.doi.org/10.1242/jcs.02528; PMID: 16129888
- Hurd TW, Gao L, Roh MH, Macara IG, Margolis B. Direct interaction of two polarity complexes implicated in epithelial tight junction assembly. Nat Cell Biol 2003; 5:137 - 42; http://dx.doi.org/10.1038/ncb923; PMID: 12545177
- Roh MH, Liu CJ, Laurinec S, Margolis B. The carboxyl terminus of zona occludens-3 binds and recruits a mammalian homologue of discs lost to tight junctions. J Biol Chem 2002; 277:27501 - 9; http://dx.doi.org/10.1074/jbc.M201177200; PMID: 12021270
- Ebnet K, Suzuki A, Horikoshi Y, Hirose T, Meyer Zu Brickwedde MK, Ohno S, Vestweber D. The cell polarity protein ASIP/PAR-3 directly associates with junctional adhesion molecule (JAM). EMBO J 2001; 20:3738 - 48; http://dx.doi.org/10.1093/emboj/20.14.3738; PMID: 11447115
- Feng W, Wu H, Chan LN, Zhang M. Par-3-mediated junctional localization of the lipid phosphatase PTEN is required for cell polarity establishment. J Biol Chem 2008; 283:23440 - 9; http://dx.doi.org/10.1074/jbc.M802482200; PMID: 18550519
- Chen X, Macara IG. Par-3 controls tight junction assembly through the Rac exchange factor Tiam1. Nat Cell Biol 2005; 7:262 - 9; http://dx.doi.org/10.1038/ncb1226; PMID: 15723052
- Martin-Belmonte F, Gassama A, Datta A, Yu W, Rescher U, Gerke V, Mostov K. PTEN-mediated apical segregation of phosphoinositides controls epithelial morphogenesis through Cdc42. Cell 2007; 128:383 - 97; http://dx.doi.org/10.1016/j.cell.2006.11.051; PMID: 17254974
- Nagasaka K, Nakagawa S, Yano T, Takizawa S, Matsumoto Y, Tsuruga T, Nakagawa K, Minaguchi T, Oda K, Hiraike-Wada O, et al. Human homolog of Drosophila tumor suppressor Scribble negatively regulates cell-cycle progression from G1 to S phase by localizing at the basolateral membrane in epithelial cells. Cancer Sci 2006; 97:1217 - 25; http://dx.doi.org/10.1111/j.1349-7006.2006.00315.x; PMID: 16965391
- Albertson R, Chabu C, Sheehan A, Doe CQ. Scribble protein domain mapping reveals a multistep localization mechanism and domains necessary for establishing cortical polarity. J Cell Sci 2004; 117:6061 - 70; http://dx.doi.org/10.1242/jcs.01525; PMID: 15536119
- Ivanov AI, Young C, Den Beste K, Capaldo CT, Humbert PO, Brennwald P, Parkos CA, Nusrat A. Tumor suppressor scribble regulates assembly of tight junctions in the intestinal epithelium. Am J Pathol 2010; 176:134 - 45; http://dx.doi.org/10.2353/ajpath.2010.090220; PMID: 19959811
- Navarro C, Nola S, Audebert S, Santoni MJ, Arsanto JP, Ginestier C, Marchetto S, Jacquemier J, Isnardon D, Le Bivic A, et al. Junctional recruitment of mammalian Scribble relies on E-cadherin engagement. Oncogene 2005; 24:4330 - 9; http://dx.doi.org/10.1038/sj.onc.1208632; PMID: 15806148
- Bilder D, Perrimon N. Localization of apical epithelial determinants by the basolateral PDZ protein Scribble. Nature 2000; 403:676 - 80; http://dx.doi.org/10.1038/35001108; PMID: 10688207
- Qin Y, Capaldo C, Gumbiner BM, Macara IG. The mammalian Scribble polarity protein regulates epithelial cell adhesion and migration through E-cadherin. J Cell Biol 2005; 171:1061 - 71; http://dx.doi.org/10.1083/jcb.200506094; PMID: 16344308
- Martin-Belmonte F, Perez-Moreno M. Epithelial cell polarity, stem cells and cancer. Nat Rev Cancer 2012; 12:23 - 38; PMID: 22169974
- Aigner K, Dampier B, Descovich L, Mikula M, Sultan A, Schreiber M, Mikulits W, Brabletz T, Strand D, Obrist P, et al. The transcription factor ZEB1 (deltaEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene 2007; 26:6979 - 88; http://dx.doi.org/10.1038/sj.onc.1210508; PMID: 17486063
- Nolan ME, Aranda V, Lee S, Lakshmi B, Basu S, Allred DC, Muthuswamy SK. The polarity protein Par6 induces cell proliferation and is overexpressed in breast cancer. Cancer Res 2008; 68:8201 - 9; http://dx.doi.org/10.1158/0008-5472.CAN-07-6567; PMID: 18922891
- Aranda V, Haire T, Nolan ME, Calarco JP, Rosenberg AZ, Fawcett JP, Pawson T, Muthuswamy SK. Par6-aPKC uncouples ErbB2 induced disruption of polarized epithelial organization from proliferation control. Nat Cell Biol 2006; 8:1235 - 45; http://dx.doi.org/10.1038/ncb1485; PMID: 17060907
- Zhan L, Rosenberg A, Bergami KC, Yu M, Xuan Z, Jaffe AB, Allred C, Muthuswamy SK. Deregulation of scribble promotes mammary tumorigenesis and reveals a role for cell polarity in carcinoma. Cell 2008; 135:865 - 78; http://dx.doi.org/10.1016/j.cell.2008.09.045; PMID: 19041750
- Osanai M, Murata M, Nishikiori N, Chiba H, Kojima T, Sawada N. Occludin-mediated premature senescence is a fail-safe mechanism against tumorigenesis in breast carcinoma cells. Cancer Sci 2007; 98:1027 - 34; http://dx.doi.org/10.1111/j.1349-7006.2007.00494.x; PMID: 17459053
- Hoover KB, Liao SY, Bryant PJ. Loss of the tight junction MAGUK ZO-1 in breast cancer: relationship to glandular differentiation and loss of heterozygosity. Am J Pathol 1998; 153:1767 - 73; http://dx.doi.org/10.1016/S0002-9440(10)65691-X; PMID: 9846967
- Mandell KJ, Babbin BA, Nusrat A, Parkos CA. Junctional adhesion molecule 1 regulates epithelial cell morphology through effects on beta1 integrins and Rap1 activity. J Biol Chem 2005; 280:11665 - 74; http://dx.doi.org/10.1074/jbc.M412650200; PMID: 15677455
- Naik MU, Naik TU, Suckow AT, Duncan MK, Naik UP. Attenuation of junctional adhesion molecule-A is a contributing factor for breast cancer cell invasion. Cancer Res 2008; 68:2194 - 203; http://dx.doi.org/10.1158/0008-5472.CAN-07-3057; PMID: 18381425
- McSherry EA, McGee SF, Jirstrom K, Doyle EM, Brennan DJ, Landberg G, Dervan PA, Hopkins AM, Gallagher WM. JAM-A expression positively correlates with poor prognosis in breast cancer patients. Int J Cancer 2009; 125:1343 - 51; http://dx.doi.org/10.1002/ijc.24498; PMID: 19533747
- Brennan K, McSherry EA, Hudson L, Kay EW, Hill AD, Young LS, et al. Junctional adhesion molecule-A is co-expressed with HER2 in breast tumors and acts as a novel regulator of HER2 protein degradation and signaling. Oncogene 2012; PMID: 22751120
- Götte M, Mohr C, Koo CY, Stock C, Vaske AK, Viola M, Ibrahim SA, Peddibhotla S, Teng YH, Low JY, et al. miR-145-dependent targeting of junctional adhesion molecule A and modulation of fascin expression are associated with reduced breast cancer cell motility and invasiveness. Oncogene 2010; 29:6569 - 80; http://dx.doi.org/10.1038/onc.2010.386; PMID: 20818426
- Murakami M, Giampietro C, Giannotta M, Corada M, Torselli I, Orsenigo F, Cocito A, d’Ario G, Mazzarol G, Confalonieri S, et al. Abrogation of junctional adhesion molecule-A expression induces cell apoptosis and reduces breast cancer progression. PLoS One 2011; 6:e21242; http://dx.doi.org/10.1371/journal.pone.0021242; PMID: 21695058
- Christofori G. New signals from the invasive front. Nature 2006; 441:444 - 50; http://dx.doi.org/10.1038/nature04872; PMID: 16724056
- Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell 2009; 139:871 - 90; http://dx.doi.org/10.1016/j.cell.2009.11.007; PMID: 19945376
- Nieman MT, Prudoff RS, Johnson KR, Wheelock MJ. N-cadherin promotes motility in human breast cancer cells regardless of their E-cadherin expression. J Cell Biol 1999; 147:631 - 44; http://dx.doi.org/10.1083/jcb.147.3.631; PMID: 10545506
- Hashizume R, Koizumi H, Ihara A, Ohta T, Uchikoshi T. Expression of beta-catenin in normal breast tissue and breast carcinoma: a comparative study with epithelial cadherin and alpha-catenin. Histopathology 1996; 29:139 - 46; http://dx.doi.org/10.1046/j.1365-2559.1996.d01-499.x; PMID: 8872147
- Rasbridge SA, Gillett CE, Sampson SA, Walsh FS, Millis RR. Epithelial (E-) and placental (P-) cadherin cell adhesion molecule expression in breast carcinoma. J Pathol 1993; 169:245 - 50; http://dx.doi.org/10.1002/path.1711690211; PMID: 8383197
- Moll R, Mitze M, Frixen UH, Birchmeier W. Differential loss of E-cadherin expression in infiltrating ductal and lobular breast carcinomas. Am J Pathol 1993; 143:1731 - 42; PMID: 8256859
- Vos CB, Cleton-Jansen AM, Berx G, de Leeuw WJ, ter Haar NT, van Roy F, Cornelisse CJ, Peterse JL, van de Vijver MJ. E-cadherin inactivation in lobular carcinoma in situ of the breast: an early event in tumorigenesis. Br J Cancer 1997; 76:1131 - 3; http://dx.doi.org/10.1038/bjc.1997.523; PMID: 9365159
- De Leeuw WJ, Berx G, Vos CB, Peterse JL, Van de Vijver MJ, Litvinov S, Van Roy F, Cornelisse CJ, Cleton-Jansen AM. Simultaneous loss of E-cadherin and catenins in invasive lobular breast cancer and lobular carcinoma in situ. J Pathol 1997; 183:404 - 11; http://dx.doi.org/10.1002/(SICI)1096-9896(199712)183:4<404::AID-PATH1148>3.0.CO;2-9; PMID: 9496256
- Berx G, Becker KF, Höfler H, van Roy F. Mutations of the human E-cadherin (CDH1) gene. Hum Mutat 1998; 12:226 - 37; http://dx.doi.org/10.1002/(SICI)1098-1004(1998)12:4<226::AID-HUMU2>3.0.CO;2-D; PMID: 9744472
- Adachi Y, Takeuchi T, Nagayama T, Ohtsuki Y, Furihata M. Zeb1-mediated T-cadherin repression increases the invasive potential of gallbladder cancer. FEBS Lett 2009; 583:430 - 6; http://dx.doi.org/10.1016/j.febslet.2008.12.042; PMID: 19116147
- Schmalhofer O, Brabletz S, Brabletz T. E-cadherin, beta-catenin, and ZEB1 in malignant progression of cancer. Cancer Metastasis Rev 2009; 28:151 - 66; http://dx.doi.org/10.1007/s10555-008-9179-y; PMID: 19153669
- Strathdee G. Epigenetic versus genetic alterations in the inactivation of E-cadherin. Semin Cancer Biol 2002; 12:373 - 9; http://dx.doi.org/10.1016/S1044-579X(02)00057-3; PMID: 12191636
- Kang Y, Massagué J. Epithelial-mesenchymal transitions: twist in development and metastasis. Cell 2004; 118:277 - 9; http://dx.doi.org/10.1016/j.cell.2004.07.011; PMID: 15294153
- De Craene B, van Roy F, Berx G. Unraveling signalling cascades for the Snail family of transcription factors. Cell Signal 2005; 17:535 - 47; http://dx.doi.org/10.1016/j.cellsig.2004.10.011; PMID: 15683729
- Peinado H, Ballestar E, Esteller M, Cano A. Snail mediates E-cadherin repression by the recruitment of the Sin3A/histone deacetylase 1 (HDAC1)/HDAC2 complex. Mol Cell Biol 2004; 24:306 - 19; http://dx.doi.org/10.1128/MCB.24.1.306-319.2004; PMID: 14673164
- Peinado H, Del Carmen Iglesias-de la Cruz M, Olmeda D, Csiszar K, Fong KS, Vega S, Nieto MA, Cano A, Portillo F. A molecular role for lysyl oxidase-like 2 enzyme in snail regulation and tumor progression. EMBO J 2005; 24:3446 - 58; http://dx.doi.org/10.1038/sj.emboj.7600781; PMID: 16096638
- Howe LR, Watanabe O, Leonard J, Brown AM. Twist is up-regulated in response to Wnt1 and inhibits mouse mammary cell differentiation. Cancer Res 2003; 63:1906 - 13; PMID: 12702582
- Saxena M, Stephens MA, Pathak H, Rangarajan A. Transcription factors that mediate epithelial-mesenchymal transition lead to multidrug resistance by upregulating ABC transporters. Cell Death Dis 2011; 2:e179; http://dx.doi.org/10.1038/cddis.2011.61; PMID: 21734725
- Cheng GZ, Chan J, Wang Q, Zhang W, Sun CD, Wang LH. Twist transcriptionally up-regulates AKT2 in breast cancer cells leading to increased migration, invasion, and resistance to paclitaxel. Cancer Res 2007; 67:1979 - 87; http://dx.doi.org/10.1158/0008-5472.CAN-06-1479; PMID: 17332325
- Suyama K, Shapiro I, Guttman M, Hazan RB. A signaling pathway leading to metastasis is controlled by N-cadherin and the FGF receptor. Cancer Cell 2002; 2:301 - 14; http://dx.doi.org/10.1016/S1535-6108(02)00150-2; PMID: 12398894
- Lin HJ, Zuo T, Chao JR, Peng Z, Asamoto LK, Yamashita SS, Huang TH. Seed in soil, with an epigenetic view. Biochim Biophys Acta 2009; 1790:920 - 4; http://dx.doi.org/10.1016/j.bbagen.2008.12.004; PMID: 19162126
- Fedor-Chaiken M, Hein PW, Stewart JC, Brackenbury R, Kinch MS. E-cadherin binding modulates EGF receptor activation. Cell Commun Adhes 2003; 10:105 - 18; PMID: 14681060
- Lin SY, Xia W, Wang JC, Kwong KY, Spohn B, Wen Y, Pestell RG, Hung MC. Beta-catenin, a novel prognostic marker for breast cancer: its roles in cyclin D1 expression and cancer progression. Proc Natl Acad Sci U S A 2000; 97:4262 - 6; http://dx.doi.org/10.1073/pnas.060025397; PMID: 10759547
- Ozaki S, Ikeda S, Ishizaki Y, Kurihara T, Tokumoto N, Iseki M, Arihiro K, Kataoka T, Okajima M, Asahara T. Alterations and correlations of the components in the Wnt signaling pathway and its target genes in breast cancer. Oncol Rep 2005; 14:1437 - 43; PMID: 16273236
- Prasad CP, Mirza S, Sharma G, Prashad R, DattaGupta S, Rath G, Ralhan R. Epigenetic alterations of CDH1 and APC genes: relationship with activation of Wnt/beta-catenin pathway in invasive ductal carcinoma of breast. Life Sci 2008; 83:318 - 25; http://dx.doi.org/10.1016/j.lfs.2008.06.019; PMID: 18662704
- Karayiannakis AJ, Nakopoulou L, Gakiopoulou H, Keramopoulos A, Davaris PS, Pignatelli M. Expression patterns of beta-catenin in in situ and invasive breast cancer. Eur J Surg Oncol 2001; 27:31 - 6; http://dx.doi.org/10.1053/ejso.1999.1017; PMID: 11237489
- Nagahata T, Shimada T, Harada A, Nagai H, Onda M, Yokoyama S, Shiba T, Jin E, Kawanami O, Emi M. Amplification, up-regulation and over-expression of DVL-1, the human counterpart of the Drosophila disheveled gene, in primary breast cancers. Cancer Sci 2003; 94:515 - 8; http://dx.doi.org/10.1111/j.1349-7006.2003.tb01475.x; PMID: 12824876
- Liu CC, Prior J, Piwnica-Worms D, Bu G. LRP6 overexpression defines a class of breast cancer subtype and is a target for therapy. Proc Natl Acad Sci U S A 2010; 107:5136 - 41; http://dx.doi.org/10.1073/pnas.0911220107; PMID: 20194742
- Björklund P, Svedlund J, Olsson AK, Akerström G, Westin G. The internally truncated LRP5 receptor presents a therapeutic target in breast cancer. PLoS One 2009; 4:e4243; http://dx.doi.org/10.1371/journal.pone.0004243; PMID: 19158955
- Jönsson M, Dejmek J, Bendahl PO, Andersson T. Loss of Wnt-5a protein is associated with early relapse in invasive ductal breast carcinomas. Cancer Res 2002; 62:409 - 16; PMID: 11809689
- Biswas DK, Shi Q, Baily S, Strickland I, Ghosh S, Pardee AB, Iglehart JD. NF-kappa B activation in human breast cancer specimens and its role in cell proliferation and apoptosis. Proc Natl Acad Sci U S A 2004; 101:10137 - 42; http://dx.doi.org/10.1073/pnas.0403621101; PMID: 15220474
- Ryo A, Nakamura M, Wulf G, Liou YC, Lu KP. Pin1 regulates turnover and subcellular localization of beta-catenin by inhibiting its interaction with APC. Nat Cell Biol 2001; 3:793 - 801; http://dx.doi.org/10.1038/ncb0901-793; PMID: 11533658
- Wulf GM, Ryo A, Wulf GG, Lee SW, Niu T, Petkova V, Lu KP. Pin1 is overexpressed in breast cancer and cooperates with Ras signaling in increasing the transcriptional activity of c-Jun towards cyclin D1. EMBO J 2001; 20:3459 - 72; http://dx.doi.org/10.1093/emboj/20.13.3459; PMID: 11432833
- Zhao H, Cui Y, Dupont J, Sun H, Hennighausen L, Yakar S. Overexpression of the tumor suppressor gene phosphatase and tensin homologue partially inhibits wnt-1-induced mammary tumorigenesis. Cancer Res 2005; 65:6864 - 73; http://dx.doi.org/10.1158/0008-5472.CAN-05-0181; PMID: 16061670
- Depowski PL, Rosenthal SI, Ross JS. Loss of expression of the PTEN gene protein product is associated with poor outcome in breast cancer. Mod Pathol 2001; 14:672 - 6; http://dx.doi.org/10.1038/modpathol.3880371; PMID: 11454999
- Knudsen KA, Lin CY, Johnson KR, Wheelock MJ, Keshgegian AA, Soler AP. Lack of correlation between serum levels of E- and P-cadherin fragments and the presence of breast cancer. Hum Pathol 2000; 31:961 - 5; http://dx.doi.org/10.1053/hupa.2000.9074; PMID: 10987257
- Turashvili G, McKinney SE, Goktepe O, Leung SC, Huntsman DG, Gelmon KA, Los G, Rejto PA, Aparicio SA. P-cadherin expression as a prognostic biomarker in a 3992 case tissue microarray series of breast cancer. Mod Pathol 2011; 24:64 - 81; http://dx.doi.org/10.1038/modpathol.2010.189; PMID: 20852590
- Sousa B, Paredes J, Milanezi F, Lopes N, Martins D, Dufloth R, Vieira D, Albergaria A, Veronese L, Carneiro V, et al. P-cadherin, vimentin and CK14 for identification of basal-like phenotype in breast carcinomas: an immunohistochemical study. Histol Histopathol 2010; 25:963 - 74; PMID: 20552547
- Albergaria A, Ribeiro AS, Vieira AF, Sousa B, Nobre AR, Seruca R, Schmitt F, Paredes J. P-cadherin role in normal breast development and cancer. Int J Dev Biol 2011; 55:811 - 22; http://dx.doi.org/10.1387/ijdb.113382aa; PMID: 22161837
- Lou Y, Preobrazhenska O, auf dem Keller U, Sutcliffe M, Barclay L, McDonald PC, Roskelley C, Overall CM, Dedhar S. Epithelial-mesenchymal transition (EMT) is not sufficient for spontaneous murine breast cancer metastasis. Dev Dyn 2008; 237:2755 - 68; http://dx.doi.org/10.1002/dvdy.21658; PMID: 18773493
- Ben Hamida A, Labidi IS, Mrad K, Charafe-Jauffret E, Ben Arab S, Esterni B, Xerri L, Viens P, Bertucci F, Birnbaum D, et al. Markers of subtypes in inflammatory breast cancer studied by immunohistochemistry: prominent expression of P-cadherin. BMC Cancer 2008; 8:28; http://dx.doi.org/10.1186/1471-2407-8-28; PMID: 18230143
- Davis MA, Reynolds AB. Blocked acinar development, E-cadherin reduction, and intraepithelial neoplasia upon ablation of p120-catenin in the mouse salivary gland. Dev Cell 2006; 10:21 - 31; http://dx.doi.org/10.1016/j.devcel.2005.12.004; PMID: 16399075
- Stairs DB, Bayne LJ, Rhoades B, Vega ME, Waldron TJ, Kalabis J, Klein-Szanto A, Lee JS, Katz JP, Diehl JA, et al. Deletion of p120-catenin results in a tumor microenvironment with inflammation and cancer that establishes it as a tumor suppressor gene. Cancer Cell 2011; 19:470 - 83; http://dx.doi.org/10.1016/j.ccr.2011.02.007; PMID: 21481789
- van Amerongen R, Berns A. Knockout mouse models to study Wnt signal transduction. Trends Genet 2006; 22:678 - 89; http://dx.doi.org/10.1016/j.tig.2006.10.001; PMID: 17045694
- Wolf K, Friedl P. Molecular mechanisms of cancer cell invasion and plasticity. Br J Dermatol 2006; 154:Suppl 1 11 - 5; http://dx.doi.org/10.1111/j.1365-2133.2006.07231.x; PMID: 16712711
- Wolf K, Mazo I, Leung H, Engelke K, von Andrian UH, Deryugina EI, Strongin AY, Bröcker EB, Friedl P. Compensation mechanism in tumor cell migration: mesenchymal-amoeboid transition after blocking of pericellular proteolysis. J Cell Biol 2003; 160:267 - 77; http://dx.doi.org/10.1083/jcb.200209006; PMID: 12527751
- Dumin JA, Dickeson SK, Stricker TP, Bhattacharyya-Pakrasi M, Roby JD, Santoro SA, Parks WC. Pro-collagenase-1 (matrix metalloproteinase-1) binds the alpha(2)beta(1) integrin upon release from keratinocytes migrating on type I collagen. J Biol Chem 2001; 276:29368 - 74; http://dx.doi.org/10.1074/jbc.M104179200; PMID: 11359786
- Webb DJ, Parsons JT, Horwitz AF. Adhesion assembly, disassembly and turnover in migrating cells -- over and over and over again. Nat Cell Biol 2002; 4:E97 - 100; http://dx.doi.org/10.1038/ncb0402-e97; PMID: 11944043
- Friedl P, Noble PB, Walton PA, Laird DW, Chauvin PJ, Tabah RJ, Black M, Zänker KS. Migration of coordinated cell clusters in mesenchymal and epithelial cancer explants in vitro. Cancer Res 1995; 55:4557 - 60; PMID: 7553628
- Davies M, Robinson M, Smith E, Huntley S, Prime S, Paterson I. Induction of an epithelial to mesenchymal transition in human immortal and malignant keratinocytes by TGF-beta1 involves MAPK, Smad and AP-1 signalling pathways. J Cell Biochem 2005; 95:918 - 31; http://dx.doi.org/10.1002/jcb.20458; PMID: 15861394
- Mandell KJ, Parkos CA. The JAM family of proteins. Adv Drug Deliv Rev 2005; 57:857 - 67; http://dx.doi.org/10.1016/j.addr.2005.01.005; PMID: 15820556
- McSherry EA, Brennan K, Hudson L, Hill AD, Hopkins AM. Breast cancer cell migration is regulated through junctional adhesion molecule-A-mediated activation of Rap1 GTPase. Breast Cancer Res 2011; 13:R31; http://dx.doi.org/10.1186/bcr2853; PMID: 21429211
- Furuse M, Tsukita S. Claudins in occluding junctions of humans and flies. Trends Cell Biol 2006; 16:181 - 8; http://dx.doi.org/10.1016/j.tcb.2006.02.006; PMID: 16537104
- Anderson JM, Van Itallie CM, Fanning AS. Setting up a selective barrier at the apical junction complex. Curr Opin Cell Biol 2004; 16:140 - 5; http://dx.doi.org/10.1016/j.ceb.2004.01.005; PMID: 15196556
- Webb PG, Spillman MA, Baumgartner HK. Claudins play a role in normal and tumor cell motility. BMC Cell Biol 2013; 14:19; http://dx.doi.org/10.1186/1471-2121-14-19; PMID: 23521713
- Lanigan F, McKiernan E, Brennan DJ, Hegarty S, Millikan RC, McBryan J, Jirstrom K, Landberg G, Martin F, Duffy MJ, et al. Increased claudin-4 expression is associated with poor prognosis and high tumour grade in breast cancer. Int J Cancer 2009; 124:2088 - 97; http://dx.doi.org/10.1002/ijc.24159; PMID: 19142967
- Escudero-Esparza A, Jiang WG, Martin TA. Claudin-5 is involved in breast cancer cell motility through the N-WASP and ROCK signalling pathways. J Exp Clin Cancer Res 2012; 31:43; http://dx.doi.org/10.1186/1756-9966-31-43; PMID: 22559840
- Wu Q, Liu Y, Ren Y, Xu X, Yu L, Li Y, Quan C. Tight junction protein, claudin-6, downregulates the malignant phenotype of breast carcinoma. Eur J Cancer Prev 2010; 19:186 - 94; http://dx.doi.org/10.1097/CEJ.0b013e328337210e; PMID: 20215972
- Martin TA, Harrison GM, Watkins G, Jiang WG. Claudin-16 reduces the aggressive behavior of human breast cancer cells. J Cell Biochem 2008; 105:41 - 52; http://dx.doi.org/10.1002/jcb.21797; PMID: 18442037
- Osanai M, Murata M, Chiba H, Kojima T, Sawada N. Epigenetic silencing of claudin-6 promotes anchorage-independent growth of breast carcinoma cells. Cancer Sci 2007; 98:1557 - 62; http://dx.doi.org/10.1111/j.1349-7006.2007.00569.x; PMID: 17645772
- Tokés AM, Kulka J, Paku S, Szik A, Páska C, Novák PK, Szilák L, Kiss A, Bögi K, Schaff Z. Claudin-1, -3 and -4 proteins and mRNA expression in benign and malignant breast lesions: a research study. Breast Cancer Res 2005; 7:R296 - 305; http://dx.doi.org/10.1186/bcr983; PMID: 15743508
- Blanchard AA, Skliris GP, Watson PH, Murphy LC, Penner C, Tomes L, Young TL, Leygue E, Myal Y. Claudins 1, 3, and 4 protein expression in ER negative breast cancer correlates with markers of the basal phenotype. Virchows Arch 2009; 454:647 - 56; http://dx.doi.org/10.1007/s00428-009-0770-6; PMID: 19387682
- Herschkowitz JI, Simin K, Weigman VJ, Mikaelian I, Usary J, Hu Z, Rasmussen KE, Jones LP, Assefnia S, Chandrasekharan S, et al. Identification of conserved gene expression features between murine mammary carcinoma models and human breast tumors. Genome Biol 2007; 8:R76; http://dx.doi.org/10.1186/gb-2007-8-5-r76; PMID: 17493263
- Ladwein M, Pape UF, Schmidt DS, Schnölzer M, Fiedler S, Langbein L, Franke WW, Moldenhauer G, Zöller M. The cell-cell adhesion molecule EpCAM interacts directly with the tight junction protein claudin-7. Exp Cell Res 2005; 309:345 - 57; http://dx.doi.org/10.1016/j.yexcr.2005.06.013; PMID: 16054130
- Lei Z, Maeda T, Tamura A, Nakamura T, Yamazaki Y, Shiratori H, Yashiro K, Tsukita S, Hamada H. EpCAM contributes to formation of functional tight junction in the intestinal epithelium by recruiting claudin proteins. Dev Biol 2012; 371:136 - 45; http://dx.doi.org/10.1016/j.ydbio.2012.07.005; PMID: 22819673
- Wu CJ, Mannan P, Lu M, Udey MC. Epithelial cell adhesion molecule (EpCAM) regulates claudin dynamics and tight junctions. J Biol Chem 2013; 288:12253 - 68; http://dx.doi.org/10.1074/jbc.M113.457499; PMID: 23486470
- Wheelock MJ, Soler AP, Knudsen KA. Cadherin junctions in mammary tumors. J Mammary Gland Biol Neoplasia 2001; 6:275 - 85; http://dx.doi.org/10.1023/A:1011319507155; PMID: 11547897
- Sarrió D, Palacios J, Hergueta-Redondo M, Gómez-López G, Cano A, Moreno-Bueno G. Functional characterization of E- and P-cadherin in invasive breast cancer cells. BMC Cancer 2009; 9:74; http://dx.doi.org/10.1186/1471-2407-9-74; PMID: 19257890
- Heyder C, Gloria-Maercker E, Entschladen F, Hatzmann W, Niggemann B, Zänker KS, Dittmar T. Realtime visualization of tumor cell/endothelial cell interactions during transmigration across the endothelial barrier. J Cancer Res Clin Oncol 2002; 128:533 - 8; http://dx.doi.org/10.1007/s00432-002-0377-7; PMID: 12384796
- Brandt B, Heyder C, Gloria-Maercker E, Hatzmann W, Rötger A, Kemming D, Zänker KS, Entschladen F, Dittmar T. 3D-extravasation model -- selection of highly motile and metastatic cancer cells. Semin Cancer Biol 2005; 15:387 - 95; http://dx.doi.org/10.1016/j.semcancer.2005.06.006; PMID: 16054390
- Uchide K, Sakon M, Ariyoshi H, Nakamori S, Tokunaga M, Monden M. Cancer cells cause vascular endothelial cell (vEC) retraction via 12(S)HETE secretion; the possible role of cancer cell derived microparticle. Ann Surg Oncol 2007; 14:862 - 8; http://dx.doi.org/10.1245/s10434-006-9225-3; PMID: 17103063
- Kebers F, Lewalle JM, Desreux J, Munaut C, Devy L, Foidart JM, Noël A. Induction of endothelial cell apoptosis by solid tumor cells. Exp Cell Res 1998; 240:197 - 205; http://dx.doi.org/10.1006/excr.1998.3935; PMID: 9596992
- Qi J, Chen N, Wang J, Siu CH. Transendothelial migration of melanoma cells involves N-cadherin-mediated adhesion and activation of the beta-catenin signaling pathway. Mol Biol Cell 2005; 16:4386 - 97; http://dx.doi.org/10.1091/mbc.E05-03-0186; PMID: 15987741
- Strell C, Lang K, Niggemann B, Zaenker KS, Entschladen F. Surface molecules regulating rolling and adhesion to endothelium of neutrophil granulocytes and MDA-MB-468 breast carcinoma cells and their interaction. Cell Mol Life Sci 2007; 64:3306 - 16; http://dx.doi.org/10.1007/s00018-007-7402-6; PMID: 17994288
- Tabariès S, Dupuy F, Dong Z, Monast A, Annis MG, Spicer J, Ferri LE, Omeroglu A, Basik M, Amir E, et al. Claudin-2 promotes breast cancer liver metastasis by facilitating tumor cell interactions with hepatocytes. Mol Cell Biol 2012; 32:2979 - 91; http://dx.doi.org/10.1128/MCB.00299-12; PMID: 22645303
- Tabariès S, Dong Z, Annis MG, Omeroglu A, Pepin F, Ouellet V, Russo C, Hassanain M, Metrakos P, Diaz Z, et al. Claudin-2 is selectively enriched in and promotes the formation of breast cancer liver metastases through engagement of integrin complexes. Oncogene 2011; 30:1318 - 28; http://dx.doi.org/10.1038/onc.2010.518; PMID: 21076473
- Erin N, Wang N, Xin P, Bui V, Weisz J, Barkan GA, Zhao W, Shearer D, Clawson GA. Altered gene expression in breast cancer liver metastases. Int J Cancer 2009; 124:1503 - 16; http://dx.doi.org/10.1002/ijc.24131; PMID: 19117052
- Tsukada Y, Fouad A, Pickren J, Lane W. Central nervous system metastasis from breast carcinoma. Autopsy study. Cancer 1983; 52:2359 - 54; http://dx.doi.org/10.1002/1097-0142(19831215)52:12<2349::AID-CNCR2820521231>3.0.CO;2-B; PMID: 6640508
- Cho SY, Choi HY. Causes of death and metastatic patterns in patients with mammary cancer. Ten-year autopsy study. Am J Clin Pathol 1980; 73:232 - 4; PMID: 6243853
- Pestalozzi BC, Zahrieh D, Price KN, Holmberg SB, Lindtner J, Collins J, Crivellari D, Fey MF, Murray E, Pagani O, et al, International Breast Cancer Study Group (IBCSG). Identifying breast cancer patients at risk for Central Nervous System (CNS) metastases in trials of the International Breast Cancer Study Group (IBCSG). Ann Oncol 2006; 17:935 - 44; http://dx.doi.org/10.1093/annonc/mdl064; PMID: 16603601
- Hicks DG, Short SM, Prescott NL, Tarr SM, Coleman KA, Yoder BJ, Crowe JP, Choueiri TK, Dawson AE, Budd GT, et al. Breast cancers with brain metastases are more likely to be estrogen receptor negative, express the basal cytokeratin CK5/6, and overexpress HER2 or EGFR. Am J Surg Pathol 2006; 30:1097 - 104; http://dx.doi.org/10.1097/01.pas.0000213306.05811.b9; PMID: 16931954
- Arshad F, Wang L, Sy C, Avraham S, Avraham HK. Blood-brain barrier integrity and breast cancer metastasis to the brain. Patholog Res Int 2010; 2011:920509; PMID: 21253507
- Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. Structure and function of the blood-brain barrier. Neurobiol Dis 2010; 37:13 - 25; http://dx.doi.org/10.1016/j.nbd.2009.07.030; PMID: 19664713
- Wolburg H, Wolburg-Buchholz K, Kraus J, Rascher-Eggstein G, Liebner S, Hamm S, Duffner F, Grote EH, Risau W, Engelhardt B. Localization of claudin-3 in tight junctions of the blood-brain barrier is selectively lost during experimental autoimmune encephalomyelitis and human glioblastoma multiforme. Acta Neuropathol 2003; 105:586 - 92; PMID: 12734665
- Nitta T, Hata M, Gotoh S, Seo Y, Sasaki H, Hashimoto N, Furuse M, Tsukita S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J Cell Biol 2003; 161:653 - 60; http://dx.doi.org/10.1083/jcb.200302070; PMID: 12743111
- Lee BC, Lee TH, Avraham S, Avraham HK. Involvement of the chemokine receptor CXCR4 and its ligand stromal cell-derived factor 1alpha in breast cancer cell migration through human brain microvascular endothelial cells. Mol Cancer Res 2004; 2:327 - 38; PMID: 15235108
- Li YM, Pan Y, Wei Y, Cheng X, Zhou BP, Tan M, Zhou X, Xia W, Hortobagyi GN, Yu D, et al. Upregulation of CXCR4 is essential for HER2-mediated tumor metastasis. Cancer Cell 2004; 6:459 - 69; http://dx.doi.org/10.1016/j.ccr.2004.09.027; PMID: 15542430
- Mendes O, Kim HT, Stoica G. Expression of MMP2, MMP9 and MMP3 in breast cancer brain metastasis in a rat model. Clin Exp Metastasis 2005; 22:237 - 46; http://dx.doi.org/10.1007/s10585-005-8115-6; PMID: 16158251
- Tester AM, Waltham M, Oh SJ, Bae SN, Bills MM, Walker EC, Kern FG, Stetler-Stevenson WG, Lippman ME, Thompson EW. Pro-matrix metalloproteinase-2 transfection increases orthotopic primary growth and experimental metastasis of MDA-MB-231 human breast cancer cells in nude mice. Cancer Res 2004; 64:652 - 8; http://dx.doi.org/10.1158/0008-5472.CAN-0384-2; PMID: 14744781
- Stark AM, Anuszkiewicz B, Mentlein R, Yoneda T, Mehdorn HM, Held-Feindt J. Differential expression of matrix metalloproteinases in brain- and bone-seeking clones of metastatic MDA-MB-231 breast cancer cells. J Neurooncol 2007; 81:39 - 48; http://dx.doi.org/10.1007/s11060-006-9207-0; PMID: 16850107
- Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science 1972; 175:720 - 31; http://dx.doi.org/10.1126/science.175.4023.720; PMID: 4333397
- Ipsen JH, Karlström G, Mouritsen OG, Wennerström H, Zuckermann MJ. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim Biophys Acta 1987; 905:162 - 72; http://dx.doi.org/10.1016/0005-2736(87)90020-4; PMID: 3676307
- Karnovsky MJ, Kleinfeld AM, Hoover RL, Klausner RD. The concept of lipid domains in membranes. J Cell Biol 1982; 94:1 - 6; http://dx.doi.org/10.1083/jcb.94.1.1; PMID: 6889603
- Lee AG, Birdsall NJ, Metcalfe JC, Toon PA, Warren GB. Clusters in lipid bilayers and the interpretation of thermal effects in biological membranes. Biochemistry 1974; 13:3699 - 705; http://dx.doi.org/10.1021/bi00715a013; PMID: 4368511
- van Meer G, Poorthuis BJ, Wirtz KW, Op den Kamp JA, van Deenen LL. Transbilayer distribution and mobility of phosphatidylcholine in intact erythrocyte membranes. A study with phosphatidylcholine exchange protein. Eur J Biochem 1980; 103:283 - 8; http://dx.doi.org/10.1111/j.1432-1033.1980.tb04313.x; PMID: 7363893
- Stier A, Sackmann E. Spin labels as enzyme substrates. Heterogeneous lipid distribution in liver microsomal membranes. Biochim Biophys Acta 1973; 311:400 - 8; http://dx.doi.org/10.1016/0005-2736(73)90320-9; PMID: 4354130
- Yu J, Fischman DA, Steck TL. Selective solubilization of proteins and phospholipids from red blood cell membranes by nonionic detergents. J Supramol Struct 1973; 1:233 - 48; http://dx.doi.org/10.1002/jss.400010308; PMID: 4804838
- Simons K, van Meer G. Lipid sorting in epithelial cells. Biochemistry 1988; 27:6197 - 202; http://dx.doi.org/10.1021/bi00417a001; PMID: 3064805
- Pike LJ. Rafts defined: a report on the Keystone Symposium on Lipid Rafts and Cell Function. J Lipid Res 2006; 47:1597 - 8; http://dx.doi.org/10.1194/jlr.E600002-JLR200; PMID: 16645198
- Nusrat A, Parkos CA, Verkade P, Foley CS, Liang TW, Innis-Whitehouse W, Eastburn KK, Madara JL. Tight junctions are membrane microdomains. J Cell Sci 2000; 113:1771 - 81; PMID: 10769208
- Li Q, Zhang Q, Zhang M, Wang C, Zhu Z, Li N, Li J. Effect of n-3 polyunsaturated fatty acids on membrane microdomain localization of tight junction proteins in experimental colitis. FEBS J 2008; 275:411 - 20; http://dx.doi.org/10.1111/j.1742-4658.2007.06210.x; PMID: 18167140
- Sugibayashi K, Onuki Y, Takayama K. Displacement of tight junction proteins from detergent-resistant membrane domains by treatment with sodium caprate. Eur J Pharm Sci 2009; 36:246 - 53; http://dx.doi.org/10.1016/j.ejps.2008.09.011; PMID: 19013238
- Lambert D, O’Neill CA, Padfield PJ. Methyl-beta-cyclodextrin increases permeability of Caco-2 cell monolayers by displacing specific claudins from cholesterol rich domains associated with tight junctions. Cell Physiol Biochem 2007; 20:495 - 506; http://dx.doi.org/10.1159/000107533; PMID: 17762176
- Irwin ME, Bohin N, Boerner JL. Src family kinases mediate epidermal growth factor receptor signaling from lipid rafts in breast cancer cells. Cancer Biol Ther 2011; 12:718 - 26; http://dx.doi.org/10.4161/cbt.12.8.16907; PMID: 21775822
- Irwin ME, Mueller KL, Bohin N, Ge Y, Boerner JL. Lipid raft localization of EGFR alters the response of cancer cells to the EGFR tyrosine kinase inhibitor gefitinib. J Cell Physiol 2011; 226:2316 - 28; http://dx.doi.org/10.1002/jcp.22570; PMID: 21660955
- Hitosugi T, Sato M, Sasaki K, Umezawa Y. Lipid raft specific knockdown of SRC family kinase activity inhibits cell adhesion and cell cycle progression of breast cancer cells. Cancer Res 2007; 67:8139 - 48; http://dx.doi.org/10.1158/0008-5472.CAN-06-4539; PMID: 17804726
- Roura S, Miravet S, Piedra J, García de Herreros A, Duñach M. Regulation of E-cadherin/Catenin association by tyrosine phosphorylation. J Biol Chem 1999; 274:36734 - 40; http://dx.doi.org/10.1074/jbc.274.51.36734; PMID: 10593980
- Biscardi JS, Ishizawar RC, Silva CM, Parsons SJ. Tyrosine kinase signalling in breast cancer: epidermal growth factor receptor and c-Src interactions in breast cancer. Breast Cancer Res 2000; 2:203 - 10; http://dx.doi.org/10.1186/bcr55; PMID: 11250711
- Foley J, Nickerson NK, Nam S, Allen KT, Gilmore JL, Nephew KP, Riese DJ 2nd. EGFR signaling in breast cancer: bad to the bone. Semin Cell Dev Biol 2010; 21:951 - 60; http://dx.doi.org/10.1016/j.semcdb.2010.08.009; PMID: 20813200
- Solis GP, Lüchtenborg AM, Katanaev VL. Wnt secretion and gradient formation. Int J Mol Sci 2013; 14:5130 - 45; http://dx.doi.org/10.3390/ijms14035130; PMID: 23455472
- Katanaev VL, Solis GP, Hausmann G, Buestorf S, Katanayeva N, Schrock Y, Stuermer CA, Basler K. Reggie-1/flotillin-2 promotes secretion of the long-range signalling forms of Wingless and Hedgehog in Drosophila. EMBO J 2008; 27:509 - 21; http://dx.doi.org/10.1038/sj.emboj.7601981; PMID: 18219274
- Mañes S, Mira E, Gómez-Moutón C, Lacalle RA, Keller P, Labrador JP, Martínez-A C. Membrane raft microdomains mediate front-rear polarity in migrating cells. EMBO J 1999; 18:6211 - 20; http://dx.doi.org/10.1093/emboj/18.22.6211; PMID: 10562533
- Donatello S, Babina IS, Hazelwood LD, Hill AD, Nabi IR, Hopkins AM. Lipid raft association restricts CD44-ezrin interaction and promotion of breast cancer cell migration. Am J Pathol 2012; 181:2172 - 87; http://dx.doi.org/10.1016/j.ajpath.2012.08.025; PMID: 23031255
- Ivanov AI. Pharmacological inhibition of endocytic pathways: is it specific enough to be useful?. Methods Mol Biol 2008; 440:15 - 33; http://dx.doi.org/10.1007/978-1-59745-178-9_2; PMID: 18369934
- Lim E, Vaillant F, Wu D, Forrest NC, Pal B, Hart AH, Asselin-Labat ML, Gyorki DE, Ward T, Partanen A, et al, kConFab. Aberrant luminal progenitors as the candidate target population for basal tumor development in BRCA1 mutation carriers. Nat Med 2009; 15:907 - 13; http://dx.doi.org/10.1038/nm.2000; PMID: 19648928
- Faria G, Cardoso MJ, Martins D, Bettencourt H, Amendoeira I, Schimitt F. P-cadherin as prognostic factor for loco-regional relapse in breast cancer. Acta Med Port 2012; 25:97 - 105; PMID: 22985920
- Lipponen P, Saarelainen E, Ji H, Aaltomaa S, Syrjänen K. Expression of E-cadherin (E-CD) as related to other prognostic factors and survival in breast cancer. J Pathol 1994; 174:101 - 9; http://dx.doi.org/10.1002/path.1711740206; PMID: 7965405
- Peralta Soler A, Knudsen KA, Salazar H, Han AC, Keshgegian AA. P-cadherin expression in breast carcinoma indicates poor survival. Cancer 1999; 86:1263 - 72; http://dx.doi.org/10.1002/(SICI)1097-0142(19991001)86:7<1263::AID-CNCR23>3.0.CO;2-2; PMID: 10506713
- Querzoli P, Coradini D, Pedriali M, Boracchi P, Ambrogi F, Raimondi E, La Sorda R, Lattanzio R, Rinaldi R, Lunardi M, et al. An immunohistochemically positive E-cadherin status is not always predictive for a good prognosis in human breast cancer. Br J Cancer 2010; 103:1835 - 9; http://dx.doi.org/10.1038/sj.bjc.6605991; PMID: 21063415
- Eljuga D, Bulić K, Petrovečki M, Bali V, Ozimec E, Razumović JJ. Reduced E-cadherin expression is a predictor of lower overall survival and metastatic disease in invasive ductal breast cancer. Onkologie 2012; 35:414 - 8; http://dx.doi.org/10.1159/000341071; PMID: 22846972
- Singhai R, Patil VW, Jaiswal SR, Patil SD, Tayade MB, Patil AV. E-Cadherin as a diagnostic biomarker in breast cancer. N Am J Med Sci 2011; 3:227 - 33; http://dx.doi.org/10.4297/najms.2011.3227; PMID: 22558599
- Saadatmand S, de Kruijf EM, Sajet A, Dekker-Ensink NG, van Nes JG, Putter H, Smit VT, van de Velde CJ, Liefers GJ, Kuppen PJ. Expression of cell adhesion molecules and prognosis in breast cancer. Br J Surg 2013; 100:252 - 60; http://dx.doi.org/10.1002/bjs.8980; PMID: 23175431
- Bukholm IK, Nesland JM, Kåresen R, Jacobsen U, Børresen-Dale AL. E-cadherin and alpha-, beta-, and gamma-catenin protein expression in relation to metastasis in human breast carcinoma. J Pathol 1998; 185:262 - 6; http://dx.doi.org/10.1002/(SICI)1096-9896(199807)185:3<262::AID-PATH97>3.0.CO;2-Y; PMID: 9771479
- López-Knowles E, Zardawi SJ, McNeil CM, Millar EK, Crea P, Musgrove EA, Sutherland RL, O’Toole SA. Cytoplasmic localization of beta-catenin is a marker of poor outcome in breast cancer patients. Cancer Epidemiol Biomarkers Prev 2010; 19:301 - 9; http://dx.doi.org/10.1158/1055-9965.EPI-09-0741; PMID: 20056651
- Katahira J, Sugiyama H, Inoue N, Horiguchi Y, Matsuda M, Sugimoto N. Clostridium perfringens enterotoxin utilizes two structurally related membrane proteins as functional receptors in vivo. J Biol Chem 1997; 272:26652 - 8; http://dx.doi.org/10.1074/jbc.272.42.26652; PMID: 9334247
- Kominsky SL, Vali M, Korz D, Gabig TG, Weitzman SA, Argani P, Sukumar S. Clostridium perfringens enterotoxin elicits rapid and specific cytolysis of breast carcinoma cells mediated through tight junction proteins claudin 3 and 4. Am J Pathol 2004; 164:1627 - 33; http://dx.doi.org/10.1016/S0002-9440(10)63721-2; PMID: 15111309
- Goetsch L, Haeuw JF, Beau-Larvor C, Gonzalez A, Zanna L, Malissard M, Lepecquet AM, Robert A, Bailly C, Broussas M, et al. A novel role for junctional adhesion molecule-A in tumor proliferation: modulation by an anti-JAM-A monoclonal antibody. Int J Cancer 2013; 132:1463 - 74; http://dx.doi.org/10.1002/ijc.27772; PMID: 22886345
- Shintani Y, Fukumoto Y, Chaika N, Grandgenett PM, Hollingsworth MA, Wheelock MJ, Johnson KR. ADH-1 suppresses N-cadherin-dependent pancreatic cancer progression. Int J Cancer 2008; 122:71 - 7; http://dx.doi.org/10.1002/ijc.23027; PMID: 17721921
- Li H, Price DK, Figg WD. ADH1, an N-cadherin inhibitor, evaluated in preclinical models of angiogenesis and androgen-independent prostate cancer. Anticancer Drugs 2007; 18:563 - 8; http://dx.doi.org/10.1097/CAD.0b013e328020043e; PMID: 17414625
- Perotti A, Sessa C, Mancuso A, Noberasco C, Cresta S, Locatelli A, Carcangiu ML, Passera K, Braghetti A, Scaramuzza D, et al. Clinical and pharmacological phase I evaluation of Exherin (ADH-1), a selective anti-N-cadherin peptide in patients with N-cadherin-expressing solid tumours. Ann Oncol 2009; 20:741 - 5; http://dx.doi.org/10.1093/annonc/mdn695; PMID: 19190075