Abstract
In breast cancer cells the Steroid Receptor RNA Activator (SRA) acts as scaffold of a complex containing HP1γ, LSD1, HDAC1/2 and CoREST, which contributes to repression of key hormone-inducible genes that must be kept silent in the absence of hormone.
The eukaryotic genome organized in chromatin regulates gene expression by modulating access of regulatory factors to the sequence information stored within DNA. In response to a given stimulus, sequence-specific transcription factors have to negotiate access to that information using signaling activated mechanisms that impinge on chromatin structure. In some cases, target sites are pre-organized in “open” chromatin characterized by highly sensitive DNaseI digestion as a consequence of the action of so-called “pioneer factors,” which gain access to DNA sequences in chromatin.Citation1
Nuclear receptors (NRs) are a group of transcription factors, some of which have the ability to access their binding sites in the chromatin context. For example, the progesterone receptor (PR) binds preferentially to its cognate sites when these are organized in well-positioned nucleosomes as a consequence of being part of a complex with protein kinases and other histone modifying enzymes.Citation2 A well-characterized model system for studying these processes is the regulation of the expression of the Mouse Mammary Tumour Virus (MMTV) provirus. Promoter activity is induced by glucocorticoids, progestins, androgens and, to a lesser extent by mineralocorticoids acting via several hormone responsive elements (HREs).Citation3-Citation5 When integrated in mouse or human chromosomes, the MMTV promoter is organized into positioned nucleosomes, with a nucleosome covering the HREs and the binding site for NF1.Citation6,Citation7 Progestin administration to T47D breast cancer cells carrying a single copy of an MMTV reporter gene activates the Src/Ras/Erk and the CDK2 cascades via cell membrane attached PR, leading to progestin-induced cell proliferation and activation of the MMTV promoter.Citation8 Within minutes of hormone addition two consecutive cycles of chromatin remodeling catalyzed by at least ten enzymes lead to the eviction of linker histone H1 and core histone H2A/H2B dimers, creating a suitable platform for transcription.Citation9,Citation10 We found that a similar mechanism applies to a large number of hormone-regulated genes,Citation1 however the chromatin organization of these target genes prior to hormone addition was largely unknown.
SRA RNA Acts a Scaffold of Repressive Complex
Long non-coding RNAs (ncRNAs) are a heterogeneous group of RNAs varying in length from 200 nt to over 100 kb, which are progressively recognized as relevant regulators of genomic processes in part by modulating chromatin state through the interactions with chromatin modifiers. Recent studies have revealed that Kcnq1ot1, Airn, Xist and HOTAIR ncRNAs regulate transcription of multiple target genes through epigenetic modifications.Citation11 To date two ncRNAS Growth arrest-specific 5 (Gas5) and Steroid Receptor RNA Activator (SRA) have been reported to be involved in hormone-dependent gene regulation. Gas5 silences the glucocorticoid receptor (GR) through formation of an RNA motif, mimicking the hormone responsive elements found in the promoter regions of glucocorticoid-responsive genes.Citation12 SRA was originally identified in a human B-lymphocyte library using the activation function 1 domain (AF-1) of PR as bait in yeast two-hybrid assay.Citation13 Although the yeast two-hybrid screening system is based upon protein-protein interaction Lanz et al. reported the presence of a stop codon upstream of the SRA sequence. Thus, SRA is an RNA molecule, which can selectively enhanced the AF-1 activity of class I nuclear receptors (i.e., steroid receptors: androgen receptor “AR,” ER-α, PR and GR) supporting its role as an RNA steroid receptor coactivator. In addition, SRA has been reported to mediate the insulation function of CTCF together with the dead box protein p68.Citation14 Direct binding to SRA has been observed for the following proteins: pseudouridylases Pus1p and Pus3p,Citation15 RNA helicases p68/p72Citation16 and nuclear repressors such as SHARPCitation17 and SLIRP.Citation18 These findings support a general function for SRA in addition to its role as steroid receptor coactivator.
As for its mechanism of action, SRA is known to form a complex with the coactivator SRC-1 raising the possibility that is could act by modulating the activity of a distinct class of nuclear receptor coactivator complexes.Citation13 However, most of the experiments demonstrating the co-activator function of SRA were performed with transiently transfected reporter genes that lack a proper chromatin organization. Recently, a set of microarrays studies depleting SRA in MCF7 cells and testing the response of endogenous genes to estrogen showed that SRA levels affect only mildly the activity of genes that require direct binding of ER to DNA.Citation19 In fact, some genes such as TFF1 and ACOX2 showed increase in their basal activity, indicating that SRA may also have a repressive role. It is important to note that, even though SRA was initially thought to be non-coding, several RNA isoforms have now been found to encode an endogenous protein called SRAP.Citation20,Citation21
Recently while trying to understand how the silent state of hormone-induced genes is maintained we identified SRA as part of a repressive complex containing HP1γ, LSD1, HDAC1/2, CoREST (LSD1.com) and the demethylase KDM5B.Citation22 Moreover we found that the unliganded PR binds genomic sites and targets this SRA-containing repressive complex to 20% of hormone-inducible genes, maintaining these genes silenced prior to hormone treatment. The complex is anchored via binding of HP1γ to H3K9me3. SRA interacts with PR, HP1γ and LSD1, and its depletion compromises the loading of the repressive complex to target chromatin promoting aberrant gene de-repression. Therefore in this class of target genes SRA may play a repressive role. Indeed around 470 hormone-regulated genes depend on SRA and 60% of them exhibited very low basal activity prior to hormone addition and were upregulated by SRA knockdown.Citation22 This indicates that SRA contributes to the repression of key genes by aiding the anchor of the repressive complex. On the other hand, as mentioned above, only a fraction of genes are affected by the knockdown of SRA, which implies that other pathways besides H3K9/HP1 are responsible for silencing of target genes. It is likely that the Polycomb pathways associated with the deposition of the methyl mark on lysine 27 of histone H3 could play a relevant role in this process. Recently, it was discovered that these two major epigenetic silencing modes are dynamically regulated and contribute to the early steps of embryo development.Citation23 Whether these two pathways act in a coordinated or independent manner during hormone-mediated gene repression is an issue that remains requiring further study.
What is the role of SRA in the repressive complex? It has been proposed that lncRNAs can serve as central platforms upon which relevant molecular components are assembled; in many diverse biological signaling processes, this characteristic is vital to the precise control of the specificity and dynamics of intermolecular interactions and signaling events.Citation24 In breast cancer cells we found that depletion of SRA destabilizes the repressive complex, supporting its role as scaffold, which brings together different proteins required for the repressive function.
As mentioned above some SRA isoforms can be translated into the protein SRAP. We cannot completely rule out the possibility that part of the general effect of SRA depletion could be due to the absence of SRAP. However, we found delocalization of HP1γ and LSD1 after RNase treatment and transfection with a mutated SRA version that cannot be translated into protein decreased the basal activity of target genes and rescued the phenotype of endogenous SRA depletion. In addition, the repressive complex interacts with SRA but not with SRAP and direct interaction of PR with SRA was detected in in vitro binding assays.Citation22 All these results support a repressive role for the SRA RNA.
Traditionally, proteins were thought to be the major players in various scaffolding complexes.Citation25 Recent evidence raises the possibility that lncRNAs may also play a similar role.Citation26 In addition, although only applicable to some unstructured proteins or domains ncRNAs could contribute to the structure of the proteins to which they bind. Regarding whether SRA can act locally (in cis) or globally (in trans), we found that SRA can regulate the activity of distantly located genes. Recently, evidence for a trans mode of action for HOTAIR, linc-p21 and PANDA lncRNAs in cell growth control, apoptosis and cancer progression has accumulated.Citation27-Citation29
Hormone Displaces the SRA-Containing Repressive Complex
In response to hormone, several signaling pathways are activated, among them ERK1/2 and the downstream kinase MSK1.Citation30 Activated MSK1 phosphorylates H3 at S10 and promotes displacement of the SRA-containing repressive complex from target regions. Eviction of the repressor complex allows H3 to be trimethylated at K4 and acetylated at K14, both marks are required for hNURF and hBAF loading, and chromatin remodeling necessary to allow binding of PR-complexes containing coactivators that will promote gene transcriptionCitation9,Citation31 ().
Figure 1. An SRA-containing repressive complex is involved in basal gene repression. In the absence of hormone, the repressive complex HP1γ-LSD1.com is bound to genomic target sites through the operation of three factors: i) the unliganded receptor, ii) its associated ncRNA SRA and iii) the H3K9me3. In the presence of hormone, activated MSK1 phosphorylates H3 at serine 10 and promotes HP1γ-LSD1.com eviction from target regions and recruitment to hormone-repressed genes. Whether the same complex participates in both derepression and active repression is still under investigation. Displacement of the repressive complex allows H3 to be trimethylated at K4 and acetylated at K14, both marks required for NURF and BAF loading and hormonal-dependent gene activation.Citation9,Citation31
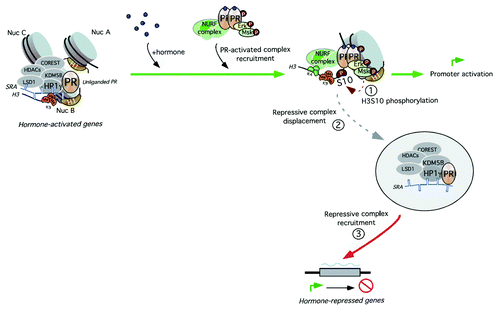
What happens to the repressor complex once displaced from its genomic target sites? In a good example of recycling of co-regulators, we have observed that this complex is recruited to hormone-repressed genes and mediates SRA-dependent active repression, most likely due to its function in maintaining the stability of the complex (unpublished data). Recently, it has been reported that protein components of the RNA-induced silencing complex (RISC) bind SRA and act as hormone receptor coregulators that are targeted to the genomic target sites of the receptors, thus linking hormone regulation of gene expression and miRNA processing.Citation32
Previous works reported that SRA is present in a ribonucleoprotein complex containing the Steroid Receptor Coactivator 1 (SRC1) and this complex is recruited by the steroid receptor to target genes in the presence of hormone.Citation13 Thus, SRA was considered to be mainly associated with the receptor in the presence of hormone and to act as an RNA co-activator, although this view was exclusively based on transient transfection experiments and therefore on targets not properly organized in chromatin. On the other hand, SRA was also reported to have a repressive function through the interaction with the SRA stem-loop interacting RNA binding protein (SLIRP) and the SMRT/HDAC1 associated repressor protein (SHARP) proteins.Citation17,Citation18 Finally, it has been proposed that SRAP could be a transcriptional regulator, able to function as a repressor though direct binding to its own RNA SRACitation33
In our studies we found several discrepancies with originally proposed role for SRA: 1) the SRA-containing repressive complex is already formed in the absence of hormone and is associated with the unliganded PR; 2) no association with SRAP has been observed; 3) SRA participates in basal repression and does not interact either with SRC1 nor SLIRP; 4) SRA is also necessary for hormone-dependent repression but this effect is independent of SLIRP and SHARP; and 5) In in vitro binding assays we found direct interaction of PR with SRA. However, we cannot exclude that SRA could be involved in hormone-dependent gene activation as part of a different complex.
The question remains of whether SRA is implicated in other normal or pathologic cellular processes? It has been reported that SRA is involved in skeletal muscle differentiation by regulating the function of MyoD.Citation34 In T47D breast cancer cells we found that SRA depletion compromises hormone-dependent proliferation and also affects apoptosis.Citation22 Elevated SRA expression profiles have been found during tumor progression,Citation35-Citation37 and the ratio of SRA/SRAP expression differs in breast cancer tumor cells, with better outcome for patients with tumors exhibiting SRAP overexpression.Citation38 These observations make SRA RNA a promising tumor-control target.Citation35-Citation37 However, transgenic mice overexpressing SRA in the mammary gland do not show an increased incidence of tumors suggesting the need for additional factors. Future experiments should focus on identifying these factors and their site of action in the genome of breast cancer cells.
Acknowledgments
The experimental work was supported by grants from the Departament d'Innovació Universitat i Empresa (DIUiE), Ministerio de Educación y Ciencia (MEC) BFU2010–01531 and BFU2011–28587. G.P.V. is a recipient of the I3 Program.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Zaret KS, Carroll JS. Pioneer transcription factors: establishing competence for gene expression. Genes Dev 2011; 25:2227 - 41; http://dx.doi.org/10.1101/gad.176826.111; PMID: 22056668
- Ballaré C, Castellano G, Gaveglia L, Althammer S, González-Vallinas J, Eyras E, et al. Nucleosome-driven transcription factor binding and gene regulation. Mol Cell 2013; 49:67 - 79; PMID: 23177737
- Hynes N, van Ooyen AJ, Kennedy N, Herrlich P, Ponta H, Groner B. Subfragments of the large terminal repeat cause glucocorticoid-responsive expression of mouse mammary tumor virus and of an adjacent gene. Proc Natl Acad Sci U S A 1983; 80:3637 - 41; http://dx.doi.org/10.1073/pnas.80.12.3637; PMID: 6304728
- Payvar F, DeFranco D, Firestone GL, Edgar B, Wrange O, Okret S, et al. Sequence-specific binding of glucocorticoid receptor to MTV DNA at sites within and upstream of the transcribed region. Cell 1983; 35:381 - 92; http://dx.doi.org/10.1016/0092-8674(83)90171-X; PMID: 6317184
- Scheidereit C, Geisse S, Westphal HM, Beato M. The glucocorticoid receptor binds to defined nucleotide sequences near the promoter of mouse mammary tumour virus. Nature 1983; 304:749 - 52; http://dx.doi.org/10.1038/304749a0; PMID: 6310405
- Richard-Foy H, Hager GL. Sequence-specific positioning of nucleosomes over the steroid-inducible MMTV promoter. EMBO J 1987; 6:2321 - 8; PMID: 2822386
- Truss M, Bartsch J, Schelbert A, Haché RJ, Beato M. Hormone induces binding of receptors and transcription factors to a rearranged nucleosome on the MMTV promoter in vivo. EMBO J 1995; 14:1737 - 51; PMID: 7737125
- Migliaccio A, Piccolo D, Castoria G, Di Domenico M, Bilancio A, Lombardi M, et al. Activation of the Src/p21ras/Erk pathway by progesterone receptor via cross-talk with estrogen receptor. EMBO J 1998; 17:2008 - 18; http://dx.doi.org/10.1093/emboj/17.7.2008; PMID: 9524123
- Vicent GP, Zaurin R, Nacht AS, Li A, Font-Mateu J, Le Dily F, et al. Two chromatin remodeling activities cooperate during activation of hormone responsive promoters. PLoS Genet 2009; 5:e1000567; http://dx.doi.org/10.1371/journal.pgen.1000567; PMID: 19609353
- Wright RH, Castellano G, Bonet J, Le Dily F, Font-Mateu J, Ballaré C, et al. CDK2-dependent activation of PARP-1 is required for hormonal gene regulation in breast cancer cells. Genes Dev 2012; 26:1972 - 83; http://dx.doi.org/10.1101/gad.193193.112; PMID: 22948662
- Saxena A, Carninci P. Long non-coding RNA modifies chromatin: epigenetic silencing by long non-coding RNAs. Bioessays 2011; 33:830 - 9; http://dx.doi.org/10.1002/bies.201100084; PMID: 21915889
- Kino T, Hurt DE, Ichijo T, Nader N, Chrousos GP. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci Signal 2010; 3:ra8; http://dx.doi.org/10.1126/scisignal.2000568; PMID: 20124551
- Lanz RB, McKenna NJ, Onate SA, Albrecht U, Wong J, Tsai SY, et al. A steroid receptor coactivator, SRA, functions as an RNA and is present in an SRC-1 complex. Cell 1999; 97:17 - 27; http://dx.doi.org/10.1016/S0092-8674(00)80711-4; PMID: 10199399
- Yao H, Brick K, Evrard Y, Xiao T, Camerini-Otero RD, Felsenfeld G. Mediation of CTCF transcriptional insulation by DEAD-box RNA-binding protein p68 and steroid receptor RNA activator SRA. Genes Dev 2010; 24:2543 - 55; http://dx.doi.org/10.1101/gad.1967810; PMID: 20966046
- Zhao X, Patton JR, Ghosh SK, Fischel-Ghodsian N, Shen L, Spanjaard RA. Pus3p- and Pus1p-dependent pseudouridylation of steroid receptor RNA activator controls a functional switch that regulates nuclear receptor signaling. Mol Endocrinol 2007; 21:686 - 99; http://dx.doi.org/10.1210/me.2006-0414; PMID: 17170069
- Watanabe M, Yanagisawa J, Kitagawa H, Takeyama K, Ogawa S, Arao Y, et al. A subfamily of RNA-binding DEAD-box proteins acts as an estrogen receptor alpha coactivator through the N-terminal activation domain (AF-1) with an RNA coactivator, SRA. EMBO J 2001; 20:1341 - 52; http://dx.doi.org/10.1093/emboj/20.6.1341; PMID: 11250900
- Shi Y, Downes M, Xie W, Kao HY, Ordentlich P, Tsai CC, et al. Sharp, an inducible cofactor that integrates nuclear receptor repression and activation. Genes Dev 2001; 15:1140 - 51; http://dx.doi.org/10.1101/gad.871201; PMID: 11331609
- Hatchell EC, Colley SM, Beveridge DJ, Epis MR, Stuart LM, Giles KM, et al. SLIRP, a small SRA binding protein, is a nuclear receptor corepressor. Mol Cell 2006; 22:657 - 68; http://dx.doi.org/10.1016/j.molcel.2006.05.024; PMID: 16762838
- Foulds CE, Tsimelzon A, Long W, Le A, Tsai SY, Tsai MJ, et al. Research resource: expression profiling reveals unexpected targets and functions of the human steroid receptor RNA activator (SRA) gene. Mol Endocrinol 2010; 24:1090 - 105; http://dx.doi.org/10.1210/me.2009-0427; PMID: 20219889
- Chooniedass-Kothari S, Emberley E, Hamedani MK, Troup S, Wang X, Czosnek A, et al. The steroid receptor RNA activator is the first functional RNA encoding a protein. FEBS Lett 2004; 566:43 - 7; http://dx.doi.org/10.1016/j.febslet.2004.03.104; PMID: 15147866
- Emberley E, Huang GJ, Hamedani MK, Czosnek A, Ali D, Grolla A, et al. Identification of new human coding steroid receptor RNA activator isoforms. Biochem Biophys Res Commun 2003; 301:509 - 15; http://dx.doi.org/10.1016/S0006-291X(02)03070-X; PMID: 12565891
- Vicent GP, Nacht AS, Zaurin R, Font-Mateu J, Soronellas D, Le Dily F, et al. Unliganded progesterone receptor-mediated targeting of an RNA-containing repressive complex silences a subset of hormone-inducible genes. Genes Dev 2013; 27:1179 - 97; http://dx.doi.org/10.1101/gad.215293.113; PMID: 23699411
- Nestorov P, Tardat M, Peters AH. H3K9/HP1 and Polycomb: two key epigenetic silencing pathways for gene regulation and embryo development. Curr Top Dev Biol 2013; 104:243 - 91; http://dx.doi.org/10.1016/B978-0-12-416027-9.00008-5; PMID: 23587244
- Spitale RC, Tsai MC, Chang HY. RNA templating the epigenome: long noncoding RNAs as molecular scaffolds. Epigenetics 2011; 6:539 - 43; http://dx.doi.org/10.4161/epi.6.5.15221; PMID: 21393997
- Good MC, Zalatan JG, Lim WA. Scaffold proteins: hubs for controlling the flow of cellular information. Science 2011; 332:680 - 6; http://dx.doi.org/10.1126/science.1198701; PMID: 21551057
- Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell 2011; 43:904 - 14; http://dx.doi.org/10.1016/j.molcel.2011.08.018; PMID: 21925379
- Gupta RA, Shah N, Wang KC, Kim J, Horlings HM, Wong DJ, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010; 464:1071 - 6; http://dx.doi.org/10.1038/nature08975; PMID: 20393566
- Huarte M, Guttman M, Feldser D, Garber M, Koziol MJ, Kenzelmann-Broz D, et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 2010; 142:409 - 19; http://dx.doi.org/10.1016/j.cell.2010.06.040; PMID: 20673990
- Hung T, Wang Y, Lin MF, Koegel AK, Kotake Y, Grant GD, et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nat Genet 2011; 43:621 - 9; http://dx.doi.org/10.1038/ng.848; PMID: 21642992
- Vicent GP, Ballaré C, Nacht AS, Clausell J, Subtil-Rodríguez A, Quiles I, et al. Induction of progesterone target genes requires activation of Erk and Msk kinases and phosphorylation of histone H3. Mol Cell 2006; 24:367 - 81; http://dx.doi.org/10.1016/j.molcel.2006.10.011; PMID: 17081988
- Vicent GP, Nacht AS, Font-Mateu J, Castellano G, Gaveglia L, Ballaré C, et al. Four enzymes cooperate to displace histone H1 during the first minute of hormonal gene activation. Genes Dev 2011; 25:845 - 62; http://dx.doi.org/10.1101/gad.621811; PMID: 21447625
- Redfern AD, Colley SM, Beveridge DJ, Ikeda N, Epis MR, Li X, et al. RNA-induced silencing complex (RISC) Proteins PACT, TRBP, and Dicer are SRA binding nuclear receptor coregulators. Proc Natl Acad Sci U S A 2013; 110:6536 - 41; http://dx.doi.org/10.1073/pnas.1301620110; PMID: 23550157
- Colley SM, Leedman PJ. Steroid Receptor RNA Activator - A nuclear receptor coregulator with multiple partners: Insights and challenges. Biochimie 2011; 93:1966 - 72; http://dx.doi.org/10.1016/j.biochi.2011.07.004; PMID: 21807064
- Caretti G, Schiltz RL, Dilworth FJ, Di Padova M, Zhao P, Ogryzko V, et al. The RNA helicases p68/p72 and the noncoding RNA SRA are coregulators of MyoD and skeletal muscle differentiation. Dev Cell 2006; 11:547 - 60; http://dx.doi.org/10.1016/j.devcel.2006.08.003; PMID: 17011493
- Lanz RB, Chua SS, Barron N, Söder BM, DeMayo F, O’Malley BW. Steroid receptor RNA activator stimulates proliferation as well as apoptosis in vivo. Mol Cell Biol 2003; 23:7163 - 76; http://dx.doi.org/10.1128/MCB.23.20.7163-7176.2003; PMID: 14517287
- Leygue E, Dotzlaw H, Watson PH, Murphy LC. Expression of estrogen receptor beta1, beta2, and beta5 messenger RNAs in human breast tissue. Cancer Res 1999; 59:1175 - 9; PMID: 10096542
- Murphy LC, Simon SL, Parkes A, Leygue E, Dotzlaw H, Snell L, et al. Altered expression of estrogen receptor coregulators during human breast tumorigenesis. Cancer Res 2000; 60:6266 - 71; PMID: 11103781
- Chooniedass-Kothari S, Hamedani MK, Troup S, Hubé F, Leygue E. The steroid receptor RNA activator protein is expressed in breast tumor tissues. Int J Cancer 2006; 118:1054 - 9; http://dx.doi.org/10.1002/ijc.21425; PMID: 16152589