
Abstract
The Fischer 344 (F344) rat was used by the National Toxicology Program (NTP) for over 5 decades for toxicity and carcinogenicity studies. However, in 2006, the NTP decided to switch to a different rat stock due largely to high background control incidences of Leydig cell tumors (LCTs) and mononuclear cell leukemia (MNCL), also known as large granular lymphocytic (LGL) leukemia. In the current review, we aim (1) to provide a summary of NTP bioassays with treatment-associated effects involving MNCL and LCTs in addition to male F344-specific tunica vaginalis mesothelioma (TVM); (2) to describe important pathobiological differences between these F344 rat tumor responses and similar target tissue-tumor response in humans; and (3) to present the NTP reasons for switching away from the F344 rat. We show that due to the highly variable background incidence of F344 MNCL, more reliance on historical control data than is usual for most tumor responses is warranted to evaluate potential effect of any chemical treatment in this rat strain. The high spontaneous incidence of LCTs in the testes of male F344 rats has made this tumor endpoint of little practical use in identifying potential testicular carcinogenic responses. TVM responses in F344 rats have a biological plausible relationship to LCTs unlike TVM in humans. Given their high spontaneous background incidence and species-specific biology, we contend that MNCL and LCT, along with TVM responses, in F344 rat carcinogenicity studies are inappropriate tumor types for human health risk assessment and lack relevance in predicting human carcinogenicity.
Introduction
The Fischer 344 (F344) rat was originally produced by Dr Maynie Rose Curtis at Columbia University in September 1920 from the 344th brother–sister mating of rats from the Fischer commercial breeder colony (Rao & Boorman Citation1990). This inbred rat became a favorite strain for use in tumor transplantation studies in the 1950s (Dunning & Curtis Citation1957). Due to its small size, what was considered at the time to be favorable fertility, and consistent response to a number of chemical carcinogens, it was selected as the rat of choice for National Cancer Institute (NCI) cancer bioassays in 1970 (Weisburger Citation1983; Cameron et al. Citation1985; Goodman et al. Citation1985). Use of the F344 rat by the NCI and the National Toxicology Program (NTP) in carcinogenicity studies over 5 decades has led to the creation of the largest rat cancer bioassay database in the world. In 2006, the NTP made a decision to switch from the F344, first to the Wistar rat and subsequently to the Sprague–Dawley rat for their toxicity and carcinogenicity studies (King-Herbert & Thayer Citation2006; King-Herbert et al. Citation2010). Because NTP toxicity and carcinogenicity testing practices have tended to create a testing paradigm followed by other investigators, it is unlikely that the F344 rat will see much use in carcinogenesis bioassays in the future.
There are multiple objectives to our review. First, to provide background on the NTP and discuss its reasons for switching away from use of the F344 rat. Second, to provide a retrospective summary and evaluation of mononuclear cell leukemia (MNCL)1, Leydig cell tumors (LCTs) and tunica vaginalis mesothelioma (TVM) in the several hundred F344 cancer bioassays conducted by the NTP. Third, to critically evaluate the relevance of these specific F344 rat tumors to their counterparts in humans.
Based on their high spontaneous background incidence and species-/strain-specific biology, our conclusion is that these tumor responses in F344 rat carcinogenicity studies differ from and/or are due to different mechanisms from those in humans. Thus, increased frequencies of these tumors in F344 rats do not predict human carcinogenicity.
This is not intended to be a comprehensive review of all existing literature on MNCL2, LCTs and TVMs, and it is not our intention to challenge the final NTP conclusions from the corresponding cancer bioassays. It is important to note that NTP conclusions are made in regards to the strength of evidence that a chemical exposure is responsible for increased incidence of neoplasms in rodents. The conclusions are not intended to evaluate human relevance. The intent of this review is to examine the large database and to specifically comment on the significance of these responses with respect to human health risk.
Methods
Source material for general information on the NTP carcinogenesis bioassays plus data and commentary on MNCL, LCTs and TVMs is derived from the publically available NTP database and published NTP toxicity/carcinogenicity technical reports (http://ntp.niehs.nih.gov).
Background incidence distributions for and were modeled using the mean incidences and range of incidences of spontaneous neoplasms in F344 rats as reported by Haseman et al. (Citation1998) for MNCL, LCT and combined adenoma and carcinoma liver tumors. Liver tumors were chosen for comparative purposes to illustrate incidence and variability of a common tumor type. The modeled distributions were generated in an Excel spreadsheet using random numbers that were based on the mean incidence and a rough standard deviation for each tumor type. The rough standard deviation was generated by taking the reported range (Incidencemax − Incidencemin) divided by four, which is an accepted statistical practice (Triola Citation2010). The random number generation was repeated 27 times to produce a modeled distribution for the feeding studies and 18 times to produce a modeled distribution for the inhalation studies. These values were selected based on the number of studies used to compile the descriptive statistics taken from Haseman et al. (Citation1998) (27 feeding studies and 18 inhalation studies).
Figure 1. Comparison in male rats of distribution of background incidence for high incidence MNCL and LCTs versus Liver tumor incidence in (A) feeding studies and (B) Chamber studies. MNCLs have both a high incidence and wide distribution in comparison to the other tumor types which makes it difficult to determine if differences between treatment and control are treatment related or due to random chance. The difference in background incidence levels for LCTs in feeding studies (A) and chamber studies (B) maybe explains why five out of the seven NTP studies identified as having increased LCT incidence were inhalation studies. Possible biological reasons for the differences in background incidence are discussed in the text.
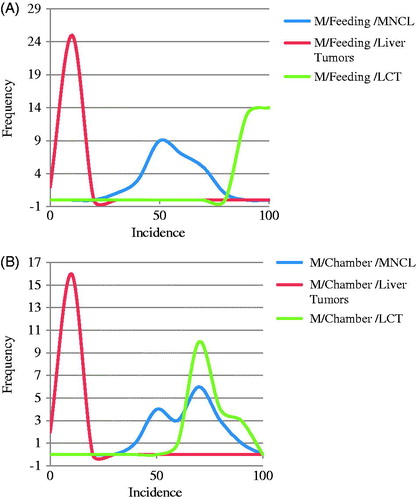
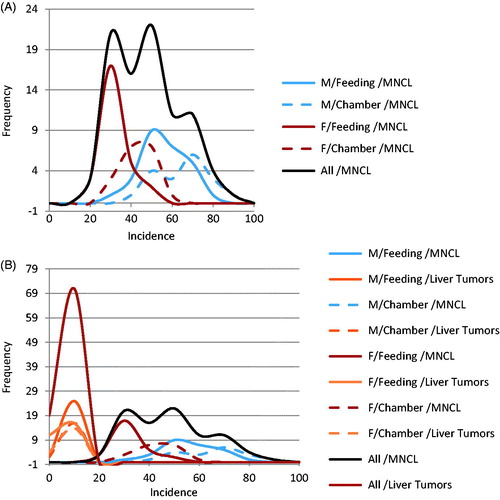
Figure 2. Highly variable and unpredictable nature of the MNCL background tumor incidence. Background incidence in Haseman at al., 1998 was separated by sex and study type. As can be seen above there are relatively large differences in background incidence for MNCL between sexes as well as between exposure methods (A). Contrast this with the effect sex and exposure type has on liver tumor incidence (B). Other factors that affect background incidence of MNCL are discussed in the text.
A brief history of the NCI/NTP carcinogenesis bioassay
As mentioned previously, the F344 rat was derived in the 1920s to fill a need for a reproducible cancer model (Lindsey Citation1979). The F344 was favored for early tumor transplantation studies (Dunning & Curtis Citation1957) due to its size and low spontaneous tumor rate (with the exception of the high rate of LCTs). Dr. Dunning provided F344 breeding stock to Walter Heston at the NCI who, in turn, provided breeding stock to the NIH Division of Research Services in 1951 (Cameron et al. Citation1985). After that the NIH colony was used as the source of F344 rats for cancer research in the 1960s. What followed were some comparisons of the F344 rat response with the Sprague–Dawley, the ACI rat and the Osborne Mendel rat with the ultimate conclusion that the F344 rat provided a more consistent response to a spectrum of chemical carcinogen classes. This led to the selection of the inbred F344 rat as the rat choice for the NCI bioassay program, largely based on the sensitivity of the F344 rat to chemically induced liver tumors (Cameron et al. Citation1985).
The NCI cancer bioassay program transferred to the NTP in 1970. The initial emphasis of the bioassay hazard assessment program was on carcinogenicity but a substantial amount of refinement occurred from the origin of the early NCI bioassays to the contemporary study design. The majority of the studies from the early 1970s have been relatively well standardized, thereby allowing meaningful retrospective study of tumor responses. The default group size has been 50 males and 50 females per dose along with increased group sizes to accommodate interim evaluations and special studies. The earliest studies involved a high dose that was the estimated maximum tolerated dose (MTD) and a lower dose that was one-half the MTD plus control groups. This approach evolved to include an additional lower dose but the conceptual reliance on an MTD has remained a part of the testing paradigm. More recent modifications of the NTP rodent bioassay testing approach include more emphasis on non-cancer endpoints, incorporation of mechanistic endpoints into studies, use of molecular biology to better understand the relevance of the observed responses and a default study design incorporating in utero exposure to assess the effects of chemical exposure through the entire life cycle.
The basic NTP testing scenario is to have the toxicity and carcinogenicity studies conducted at contract research laboratories using F344 rats from the NTP colony and with study data submitted to NTP for quality assurance and pathology peer review. A draft technical report is next prepared by NTP scientists and made publicly available for comment. The draft report is peer reviewed by an external panel of scientific experts who either endorse the conclusions of the NTP or recommend modification of those conclusions.
In their cancer bioassay technical reports that provide conclusions regarding carcinogenic responses, the NTP uses five categories of evidence of carcinogenic activity to summarize the strength of evidence observed in each species and sex. There are two categories for positive results (clear evidence and some evidence); one category for uncertain findings (equivocal evidence); one category for no observable effects (no evidence); and one category for experiments that cannot be evaluated due to major flaws (inadequate study). For the judgment of clear evidence, there is a dose-related increase of malignant neoplasms, a dose-related increase in a combination of benign and malignant neoplasms or a dose-related increase in benign neoplasms where there is evidence of progression to malignancy. In a some evidence determination, the data show a treatment-related increased incidence of neoplasia where the strength of the response is less than that for clear evidence. There may not necessarily be a clear dose-response and the neoplasms may be benign, malignant, or a combination of benign and malignant. For a call of equivocal evidence, the data are interpreted as showing a marginal increase in neoplasia that may be chemically related. No evidence of carcinogenicity is used when there is no treatment-related increase in neoplasia. These categories refer to the strength of the experimental evidence and not to potency or mechanism.
During the formal external peer review of individual draft technical reports, there are frequently discussions and debate about the levels of evidence for carcinogenicity proposed by the NTP scientists. Highlights of these discussions are included in the final technical report for each rodent carcinogenicity bioassay.
NTP switch from the F344 rat
The NTP decision to no longer use the F344 rat in its toxicity and carcinogenicity bioassays was based on several factors (King-Herbert & Thayer Citation2006; King-Herbert et al. Citation2010). Admittedly, there was some concern about switching due to loss of a very robust historical database. Given 5 decades of using the F344 rat despite the high spontaneous incidence of LCTs, it is apparent that no single factor was enough to cause a switch to a different rat strain or stock. Other problems in addition to the high spontaneous LCT incidence that prompted the decision to switch included a high and variable background occurrence of MNCL in both sexes, a high incidence of early mortality due to MNCL, time-dependent decreased fecundity and relatively poor reproductive performance, sporadic seizures and idiopathic chylothorax. With the NTP wanting to standardize a rat of choice across all studies, including reproductive toxicity assessments, and the plan for in utero exposures in future bioassays, high fecundity was paramount. The high incidence of LCTs precluded adequate assessment of testicular effects. The variable background incidence of MNCL progressively increased since the 1970s and became a major cause of early mortality in carcinogenesis studies. In addition, the MNCL response showed sporadic exacerbation by treatment or unexplained decrease following splenic toxicity. The sporadic seizures and idiopathic chylothorax were perhaps less significant issues but were additional factors leading to the decision to switch. The initial switch to the Wistar rat (Crl:WI[Han]) was short-lived due to its small litter size. The current NTP rat of choice for toxicity and carcinogenicity studies is the Sprague–Dawley (Hsd:Sprague–Dawley SD). Their choice of mouse has remained the B6C3F1.
Mononuclear cell leukemia
Early history of MNCL
The initial report of what was most likely MNCL occurred during transplantation studies of mammary adenocarcinomas in F344 rats by Dunning and Curtis (Citation1957). The leukemia cells were initially observed in blood vessels of transplanted adenocarcinomas. Upon subsequent subcutaneous passages, the leukemia cells outgrew the adenocarcinoma cells leading to progressively decreased latency between passages. After the 115th transplant generation, the leukemia growth led to death between 14 and 25 days post-transplantation with infiltration of the subcutis, lungs, liver, spleen and lymph nodes by leukemia cells and hemorrhage. Description of the leukemia cells in stained smears is consistent with morphological features of MNCL, although the characteristic eosinophilic cytoplasmic granules described in later accounts were apparently not prominent. The leukemia was readily transferred by injection of tissue fragments from the inoculation site, by injection of fragments of affected liver and by subcutaneous or intraperitoneal injection of whole blood. The clinical features of the transplanted leukemia described by Dunning are essentially the same as detailed in later reports following transplantation.
Natural history of spontaneous and transplanted MNCL
The first relatively complete descriptions of spontaneous MNCL were reported by Moloney and colleagues in inbred female Wistar–Furth rats (Moloney et al. Citation1969) and subsequently in female F344 rats (Moloney et al. Citation1970). They suggested the term MNCL based on its morphological features that differed from other known rat leukemias. They describe leukemic cells as having reddish cytoplasmic granules and disease features including splenic enlargement, leukemic infiltration of liver parenchyma and anemia with a 2- to 6-week clinical course resulting in death. Numerical and/or morphological abnormalities were seen in some metaphase spreads of the leukemic cells. In a subsequent report, Moloney and King found that splenectomy at an early age greatly reduced the incidence of MNCL in inbred Wistar–Furth and in F344 rats (sex not specified) suggesting an origin or early development occurred in the spleen (Moloney & King Citation1973). Due to the relative increase in use of F344 rats in the 1970s, MNCL was soon recognized as an important cause of spontaneous death in older F344 rats (Davey & Moloney Citation1970; Sass et al. Citation1975; Sacksteder Citation1976; Coleman et al. Citation1977; Goodman et al. Citation1979). Spontaneous MNCL is rarely seen in Sprague–Dawley rats with incidences below 1% (Frith Citation1988).
A variety of variables, factors and events influence the incidence of MNCL in F344 rats (). Since many of these variables occur in specific study designs, it is important to use the most appropriate historical control data in interpretation of study results. For this reason, NTP has maintained several historical control databases to account for some of the variables affecting MNCL incidences. The high and variable background incidence of MNCL has been clearly noted as critical in assessing the relevance of potential treatment-related increases to human health risk (Lington et al. Citation1997; Caldwell Citation1999; Thomas et al. Citation2007).
Table 1. Variables, factors and events influencing the incidence of MNCL in F344 rats.
Several definitive studies of spontaneous and transplantable MNCL beginning in the early 1980s and continuing over the following 20 years serve to extend our knowledge of the clinical and pathological features of this disease. Spontaneous MNCL arises in the splenic marginal zone with subsequent infiltration of the red pulp, splenic lymphoid depletion and erythrophagocytosis by MNCL cells (Ward & Reynolds Citation1983; Stromberg & Vogtsberger 1983; Losco & Ward Citation1984). There is secondary involvement of the liver with distention of sinusoids by leukemic cells leading to hepatocellular degeneration, some evidence of nodular regeneration (Shiga & Narama Citation2015), and necrosis in severe cases. Many other organs are affected with bone marrow involvement occurring late in the disease. Paraneoplastic events associated with spontaneous MNCL include immune mediated hemolytic anemia, thromobocytopenia, myelofibrosis and osteosclerosis in some cases (Stromberg & Vogtsberger 1983; Stromberg et al. Citation1983c). Disease onset typically starts after 18 months of age leading to death by 22 months. The spontaneous latency from identification of leukemia to death from anemia and organ failure is 10–12 weeks (Stromberg et al. Citation1983c, Citation1990) although a 5-week latency was reported during the early identification of this disease (Moloney et al. Citation1970). MNCL is the main cause of death in F344 rats 20 months and older (Sass et al. Citation1975; Sacksteder Citation1976; Coleman et al. Citation1977; Goodman et al. Citation1979; Stromberg & Vogtsberger 1983; Kodell et al. Citation1995). Clinical pathology features include severe anemia, altered coagulation parameters, white blood cell counts as high as 400,000/mm3 and elevated serum enzymes reflecting liver and other organ damage (Stromberg et al. Citation1983b, Citation1983c).
The MNCL transplant model
While initial transplantation was carried out by injection of leukemic blood or pieces of affected tissue (Dunning & Curtis Citation1957), transplantation studies of MNCL starting in the 1980s involved obtaining leukemic cells from spleens of spontaneous cases with isolation of the MNCL cells on Ficoll-isopaque gradients and adjusting cell suspensions with phosphate buffered saline (Stromberg et al. Citation1985). Isolated cells were typically injected ip or iv. In contrast to the natural disease occurring in rats older than 20 months, all ages of F344 rats regardless of sex are susceptible to the transplanted MNCL cells. Progression of the transplant disease is dependent upon the number of injected cells with a dose of 2 × 107 leukemia cells resulting in a mean longevity of 67 days (Stromberg et al. Citation1985).
Many features of the transplantation MNCL model mimic the natural spontaneous disease (Stromberg et al. Citation1985) supporting the idea of using the transplant model as a short-term assay for identification of anti-leukemia chemicals (Dieter et al. Citation1989, Citation1990). Clinical feature including emaciation, icterus and anemia are similar to spontaneous MNCL. Macroscopic hallmark features of splenomegaly, hepatomegaly, lymphadenopathy and petechial hemorrhages in lymph nodes, lungs and brain are present in both natural and transplant diseases. The morphological features and clinical pathology in the MNCL transplant disease, including paraneoplastic immune mediated hemolytic anemia, thrombocytopenia, myelofibrosis and osteosclerosis, are similar to the natural disease. As there is progressive involvement of the liver and other tissues, serum enzyme markers reflecting tissue damage are elevated.
The most notable difference in the transplantation model is a shorter clinical course with a dose-dependent and serial-dependent decreased latency. In the extreme, after 23 serial passages, rats inoculated ip with 107 leukemia cells died between 12 and 16 days after transplantation (Stromberg et al. Citation1990). Transplanted MNCL has decreased pentose-shunt enzymes and malate dehydrogenase possibly related to the rapid growth rate of transplanted tumor cells (Dieter et al. Citation1985). Other features unique to the transplantation model are an increased clinical malignancy and cytotoxicity with increased serial passage, more frequent bone marrow and lymph node infiltration, and the fact that all ages of F344 rats are susceptible to transplantation (Reynolds et al. Citation1984; Dieter et al. Citation1985; Stromberg et al. Citation1985). This latter observation suggests that the natural disease may be an age-associated genetic event.
Staging MNCL
Starting in the mid-1980s the NTP began to define different stages in the progression of MNCL in their technical reports to better define whether treatment potentially accelerated the progression of the leukemia. Three stages of MNCL have been defined (Dunnick et al. Citation1989; Stefanski et al. Citation1990; Frith et al. Citation1993; Stefanski et al. Citation1995).
Stage 1 is primarily restricted to the spleen with minimal splenic enlargement but increased leukemia cells in the splenic red pulp. Some depletion of small lymphocytes in germinal centers, periarteriolar lymphoid sheaths and marginal zones has been documented in early stages of the disease (Losco & Ward Citation1984). The liver may contain a small number of leukemia cells in the sinusoids. There is no evidence of involvement of any other organs.
Stage 2 is primarily restricted to the spleen and liver. There is splenic enlargement with increased leukemia cells expanding the red pulp but lymphoid follicles and periarteriolar lymphoid sheaths are still evident. Liver involvement includes increased leukemia cells beginning to expand the sinusoids. The vasculature in other organs may also contain leukemia cells but without obvious aggregates of leukemia cells.
Stage 3 involves advanced disease in multiple organs including lung, lymph nodes, kidney, adrenal gland and brain. Splenomegaly is pronounced with effacement of normal splenic architecture and the liver is enlarged and mottled. Centrilobular hepatocellular degeneration and necrosis are often present.
Cytological, immunophenotypic and functional features of F344 MNCL cells
Cytological features of MNCL cell have been characterized from natural cases and from different transplant cell lines (Ward & Reynolds Citation1983; Stromberg et al. Citation1985). The MNCL cell ranges from 12 to 20 microns in diameter and contains a single round to reniform eccentric nucleus with dense heterochromatin and a single nucleolus. There is abundant gray-blue cytoplasm that contains red cytoplasmic granules that are slightly less than 1 micron in size and have been shown to be lysosomes by electron microscopy (Stromberg et al. Citation1983a). MNCL cells are variably immunopositive for OX-8, BC-84 and W3/13 and have variable NK-cell activity, features characteristic of normal rat large granular lymphocytes (LGLs) (Ward & Reynolds Citation1983). More complete immunophenotypic features of MNCL cells can be found in several references (Stromberg & Vogtsberger 1983; Reynolds et al. Citation1984; Reynolds Citation1985; Ward et al. Citation1990).
In their examination of the NK functionality of MNCL cells, Reynolds and colleagues showed that both NK and antibody-dependent cell-mediated cytotoxicity was similar to that of normal LGL cells but that the MNCL cells lacked a functional T-cell receptor beta chain (Reynolds et al. Citation1985). In addition to identifying phenotypic features similar to LGL cells, Stromberg and colleagues identified adherence to glass, phagocytosis and NaF-sensitive esterase granule staining, all features of MNCL cells that are characteristic of monocytes (Stromberg et al. Citation1983a). The functional ability to phagocytose erythrocytes and expression of csf-1 receptor also suggest a relationship to a monocyte/macrophage lineage (Kusewitt et al. Citation1982; Stefanski et al. Citation1990). However, Reynolds and colleagues distinguish MNCL cells from monocytes by their lack of peroxidase, lysozyme, non-specific esterase and non-adherence in cell culture (Ward & Reynolds Citation1983; Reynolds et al. Citation1984). Thus, while an exact cell of origin for F344 rat MNCL is unknown, immunological and functional features show a relationship to lymphocytes and monocytes with strong evidence of LGL lineage. MNCL cell lines have differing degrees of NK activity (cytotoxicity) and different cell surface antigens as indication that leukemic cells are heterogeneous and not clonal (Ward & Reynolds Citation1983).
F344 MNCL and human LGL leukemia
There are two major human lymphoproliferative diseases with LGL features that have morphological resemblance to F344 rat MNCL. One is a CD3+ leukemia of LGL without NK activity and the other a surface CD3- leukemia with NK activity (Steinway et al. Citation2014).
The CD3+ LGL leukemia is cytotoxic, and is characterized by an increase in the number of mature CD8+ CD57+ T-cell receptor αβ+ T lymphocytes (Steinway et al. Citation2014). Clinically, it presents as a chronic disease, associated with autoimmune conditions (e.g. rheumatoid arthritis), recurrent infections, anemia and neutropenia (Johansson et al. Citation2015). This is an indolent condition, with median survival of more than 10 years (Steinway et al. Citation2014), and therefore, based upon pathogenesis, is clearly unrelated to the F344 MNCL (Chan et al. Citation2008a). An aggressive form of this condition has been described in the literature (Gentile et al. Citation1994; Alekshun et al. Citation2007), with clinical resemblance to the MNCL form observed in F344 rats. However, this aggressive entity is considered so rare, that it was not included in the 2008 World Health Organization Classification of Hematologic Malignancies.
The cells which are representative of the lymphoproliferative disease with lineage from CD3− LGL lymphoid cells are typically CD16+ CD56 + (Sokol & Loughran Citation2006). According to the 2008 World Health Organization Classification, two types of conditions exist: chronic NK lymphoproliferative disease (LPD), defined as a provisional entity and aggressive NK cell leukemia (ANKCL). Analogous to the CD3+ LGL leukemia, chronic NK-LPD is an indolent disorder with similar clinical phenotype, although autoimmune disorders are less prevalent in this condition (Poullot et al. Citation2014). ANKCL is extremely rare, with only 98 cases reported worldwide (R. Irons, personal communication), mostly in Asia or Central/South America (Suzuki et al. Citation2004; Semenzato et al. Citation2012). It usually presents acutely, and is associated with B symptoms (particularly fever, but also night sweats and weight loss), jaundice, lymphadenopathy, hepatosplenomegaly, circulating leukemic cells and cytopenia (Zhang et al. Citation2014). It has a multi-organ profile, and is characterized by extensive bone marrow involvement and often presents with the hemophagocytic syndrome (Hasserjian & Harris Citation2007). ANKCL bears a very poor prognosis, and is acutely fatal with median survival time from diagnosis of approximately 58 days (Song et al. Citation2002). It has an Epstein–Barr virus (EBV) etiology, and unique clonal expression of selected EBV genes found in the tumor cells in almost all of the cases (Ohshima et al. Citation1998). The pathogenesis of ANKCL in humans likely requires three key events: (1) antigenic stimulation; (2) EBV infection; and (3) dominant cellular oncogenic transformation (Ryder et al. Citation2007). It is important to note that all three of these events are required for ANKCL development in humans. Its clinical and pathological features are very similar to F344 rat MNCL (Cheung et al. Citation2003; Ruskova et al. Citation2004; Suzuki et al. Citation2004; Ryder et al. Citation2007; Chan et al. Citation2008b). However, the F344 rat MNCL does not have a viral etiology (Kawa-Ha et al. Citation1989; Hart et al. Citation1992), has a much higher incidence rate compared to ANKCL, has a variable nature, and is strain specific. Therefore, the commonly occurring F344 rat MNCL cannot be considered a relevant predictor of human disease.
NTP studies with potential MNCL responses
From 1978 to 2006 when the NTP stopped using the F344 rat in cancer bioassays, there have been 40 F344 rat studies with a potential chemical-related increased incidence of MNCL and 26 chemicals with a chemical-related decrease in MNCL in one or both sexes. The majority of these 66 studies had chemical-related increased incidence of neoplasia at tissue sites other than the hematopoietic system. Here we will focus our review on potentially positive MNCL responses. The majority of the chemicals with decreased incidences of MNCL had an associated splenic toxicity in pre-chronic studies at similar doses to the 2-year carcinogenicity studies (Elwell et al. Citation1996). Among the 40 chemicals with a potential positive MNCL response, two [bisphenol A (NTP TR215) and phenol (NTP TR203)] were ultimately concluded by the NTP to not be carcinogenic based on more appropriate statistical analysis for bisphenol A and lack of a convincing dose response for phenol. Of the remaining 38 studies, nine were documented in NTP technical reports as having MNCL as the only tumor response in one or both sexes. Consequently, levels of evidence of carcinogenicity for 29 chemical studies are based on the tumor response in the hepatic, pulmonary, renal, endocrine and/or other non-hematopoietic system tissues in addition to a MNCL response.
The rat studies with decreased incidences of MNCL are listed in . Fifteen of these 26 chemicals had one or more tumor responses in rats and/or mice primarily involving liver, kidney, lung and/or endocrine tissues. The magnitude of the MNCL decrease was dramatic, with incidence values frequently reaching 0%. The relationship of the decreased incidence to splenic toxicity underscores the importance of the splenic microenvironment in the development of MNCL. It is noted that splenectomy at 2 months of age had previously been shown to dramatically reduce the incidence of MNCL later in life (Moloney & King Citation1973).
Table 2. Decreased frequency of MNCL in NTP studies.
Identification of the 38 chemicals with potential MNCL responses along with statistical analyses is provided as Supplemental Table S1. We have listed these studies based on the levels of evidence of carcinogenicity for the MNCL response as per the data in the NTP technical reports (). This has been challenging for a variety of bioassay outcomes. When there is an overwhelming tumor response in one or more non-hematopoietic tissues, MNCL may not have received particular mention but was simply included in the overall level of evidence determination for that bioassay. In other situations where a bioassay is classified as having clear evidence of carcinogenicity in a non-hematopoietic tissue, a suspicious MNCL response may be listed as “may have been related” to treatment – basically an equivocal call. When the MNCL response in treated groups falls within the historical control range, there has often been documented discussion and debate during the draft technical report peer review regarding how to classify the leukemia response. Since MNCL is a common cause of early mortality in F344 rat studies, a life-table or poly 3 statistical analysis has tended to dominate the ultimate classification of the MNCL response with respect to its level of evidence of carcinogenicity, especially when a treatment-related increased incidence is associated with decreased survival in a treated group. For those studies where the NTP concluded that MNCL was a carcinogenic response, we note that many of the responses occurred in only one sex. In those studies where there is dose-related and robust (p < 0.01) increased incidence of MNCL in both sexes, we believe a judgment of clear evidence of carcinogenicity is appropriate.
Table 3. Male and female F344 rat MNCL responses in NTP studies.
Factors consistent with a weight-of-evidence approach in decisions regarding chemical safety and risk assessment include use of historical control data, a clear dose-response, having the effect in both sexes, lesion latency, the stage of the disease and the appropriate degree of statistical stringency. For example, in pairwise comparisons, the use of a statistical cut-off of p < 0.05 for rare tumors and p < 0.01 for commonly occurring tumors has been a reasonable suggestion (Haseman Citation1983). A major factor that has been of persistent concern in evaluating a MNCL response in NTP F344 rat bioassays is the high and variable background incidence in study controls. Exactly how the factors listed in influence the incidence of MNCL is unknown. Due to the influence of gender and route of exposure on the incidence of MNCL, NTP has employed multiple historical control databases in evaluating MNCL responses. We considered these factors in our commentary on each of the 38 studies with an indication of a treatment-related increased incidence of MNCL. Thomas et al. provide a general review of changes over time in NTP historical control rates of MNCL in tabular form, in their 2007 paper. They document a progressive increase in MNCL in males from 7.9% in 1971 to 52.5% in 1995–1998 and in females from 2.1% in 1971 to 24.2% in 1995–1998.
Early studies evaluated before use of levels of evidence of carcinogenicity
There are six early studies conducted and evaluated by NCI or NTP prior to the introduction of levels of evidence of carcinogenicity by NTP (). These studies are simply listed as positive for leukemia. Based on a review of archived slides from five of the studies, some of the leukemia in these studies is consistent with MNCL. Slides were not available for lasiocarpine (NTP TR039), a known carcinogen. The butyl benzyl phthalate (NTP TR213) study was subsequently repeated (see NTP TR458) and no treatment-related increased incidence of MNCL was present in the repeat study. For four of these six early studies there are no historical control data for MNCL. In these early studies, the data evaluation is less complete compared to subsequent studies, and MNCL was combined with lymphoma for purposes of evaluation. We are unable to draw firm conclusions regarding the MNCL incidence in these cases.
Table 4. NCI/NTP F344 rat studies with a positive leukemia response started and evaluated prior to introduction of NTP levels of evidence of carcinogenicity.
Studies with clear evidence of carcinogenicity for MNCL
Study detail and commentary on nine studies in which the MNCL response in one or both sexes was considered to be clear evidence of carcinogenicity by the NTP are summarized in . For furan (NTP TR402) and tetrafluoroethylene (NTP TR450) the data are consistent with a judgment of clear evidence of carcinogenicity. In both cases there is a robust dose-response in both sexes. For the remaining seven studies, the call of clear evidence of carcinogenicity for MNCL is debatable. For tetrachloroethylene (NTP TR311), the call of clear evidence of carcinogenicity was questioned during the formal peer review due to the high rates of MNCL in controls. MNCL incidences in treated groups were within historical control ranges for glycidol (NTP TR374) and C.I. Direct Blue 15 (NTP TR397) and these two studies along with 2,2-bis(bromomethyl)-1,3-propanediol (NTP TR452) have robust multisite tumor responses representing significant pathophysiology as well as competing causes of early mortality. In considering the appropriate degree of statistical significance for common tumor responses, the marginally significant life table statistical flags for mirex (NTP TR313), o-nitroanisole (NTP TR416) and gallium arsenide (NTP TR492) do not strongly support the conclusion that there is clear evidence of carcinogenicity for MNCL. For o-nitroanisole (NTP TR416), MNCL was the only identified tumor response in the main study, and it was suggested (but not accepted by consensus vote of the peer review panel) that the clear evidence of carcinogenicity for MNCL in both sexes be reduced to some evidence for males and equivocal evidence for females. For three studies with clear evidence of MNCL carcinogenicity in one sex, there was some evidence of carcinogenicity for MNCL in females (tetrachloroethylene NTP TR311) and equivocal evidence in males (C.I. Direct Blue 15 NTP TR397 and tetrafluoroethylene NTP TR450).
Table 5. NTP studies where at least one sex had MNCL classified as clear evidence of carcinogenicity.
In summary, with the exception of two of the nine studies with a judgment of clear evidence of carcinogenicity for MNCL, the data supporting clear evidence of carcinogenicity for MNCL from the remaining seven studies is less compelling. Clear evidence of carcinogenicity was only present in one sex for the seven studies, three studies had competing causes for reduced survival, and statistical significance is of low stringency for three studies.
Studies with some evidence of carcinogenicity for MNCL
For all the seven studies where at least one sex had a determination of some evidence of carcinogenicity for MNCL (), there was some discussion during the formal peer review about the MNCL response. The discussions during the peer review of dibromoacetic acid (NTP TR537) resulted in downgrading some evidence to equivocal evidence of carcinogenicity for MNCL in males. For riddelliine (NTP TR508) the MNCL response was initially included under clear evidence of carcinogenicity in both sexes along with liver tumors but the MNCL response is documented in the peer review comments as being more consistent with some evidence of carcinogenicity. The dimethyl morpholinophosphoramidate (NTP TR298) study represents the first time that the only evidence of a carcinogenic response in both sexes was MNCL. Consequently, this narrowly focused the comments during the formal peer review, and although the final decision was some evidence of carcinogenicity for both sexes, there was a contrary opinion expressed regarding the female MNCL responses, indicating that the review group had some uncertainty about the NTP interpretation. For females in the dibromoacetic acid (NTP TR537) study, the only tumor response was some evidence of carcinogenicity for MNCL. In this study, there was equivocal evidence of MNCL carcinogenicity in males (dibromoacetic acid NTP TR537).
Table 6. NTP studies where at least one sex had MNCL classified as some evidence of carcinogenicity.
In summary, there was discussion during the formal peer review regarding the MNCL response for all the seven studies with some evidence of MNCL carcinogenicity. Despite some comments by peer reviewers indicating that historical control data are important, the final NTP level of evidence calls for MNCL carcinogenicity were largely driven by life table statistical analyses versus concurrent controls.
Studies with equivocal evidence of carcinogenicity for MNCL
There are 16 studies where there was equivocal evidence of carcinogenicity for MNCL in one or both sexes (). In the case of acetaminophen (NTP TR 394), there was discussion during the formal peer review regarding the MNCL but the fact that there was a negative study in F344 rats at a higher dose of acetaminophen (Hiraga & Fujii, Citation1985) was not considered. In 10 of the 16 bioassays where one or both sexes had a final determination of equivocal evidence of carcinogenicity for MNCL, there was discussion centered on the MNCL response during the NTP formal peer review. For four bioassays [chlorinated water (NTP TR392), chloraminated water (NTP TR392), pyridine (NTP TR470) and 4-methylimidazole (NTP TR535)], MNCL was the only tumor response identified in one or both sexes. In those cases, the discussion during the formal peer review was particularly focused on MNCL. Different factors influenced the final judgment regarding equivocal evidence of carcinogenicity for MNCL. For 12 of the 16 bioassays, the MNCL incidences fell within the relevant historical control range or were only one percentage point above the upper range value. An unusually high incidence of MNCL in control and treated groups was present in the benzophenone (NTP TR533) study and an unusually low concurrent control value was present in the tris(2-chloroethyl) phosphate (NTP TR391) study. The initial NTP opinions were downgraded from some to equivocal evidence based on a highly variable background incidence for the diallylphthalate (NTP TR284) study and based on the lack of a convincing response in the opposite sex for pyridine (NTP TR470) in addition to the fact that a concurrent dose water study in the same lab had a 38% control incidence versus the 24% incidence for pyridine. These factors demonstrate that the NTP has sometimes considered the variable MNCL response in evaluating the level of evidence of carcinogenicity and that a potential MNCL response in only one sex is less convincing than a response present in both sexes. For the 3,3′-dimethylbenzidine dihydrochloride (NTP TR390) study terminated at 14 months, equivocal evidence of carcinogenicity for MNCL is questionable due to the relatively short duration of the study.
Table 7. Occurrence of MNCL in NTP studies with at least one sex classified as equivocal evidence of carcinogenicity.
By definition, equivocal evidence of carcinogenicity represents an uncertain judgment. This uncertainty was reflected in the discussion and debate during the formal peer review for the majority of the 16 studies with equivocal evidence of carcinogenicity for MNCL. In these studies, there were definitive treatment-related tumor responses in non-hematopoietic tissue sites. However, for 12 of the 16 studies, the MNCL response was within or only one tumor outside the relevant historical control range. When the judgment regarding MNCL carcinogenicity is uncertain, responses that fall within the historical control should get more consideration with less dependence on the implied precision of a particular statistical test.
Conclusions
Review of the natural history and characteristics of the F344 rat MNCL reveals two important features: its species- and strain-specificity and its unusually high and variable background incidence. The high and variable background incidence is present in both sexes and is in dramatic contrast to other tumor types such as hepatocellular adenomas and carcinomas ( and ). There is sufficient evidence from the literature documenting that MNCL is a distinct form of LGL leukemia that is highly prevalent in both sexes of F344 rats and extremely rare in other stocks and strains of laboratory rats. Furthermore, the only potential human LGL counterpart is an extremely rare but aggressive leukemia that, unlike the F344 MNCL, has a viral etiology. Thus, the evidence indicates that MNCL, a spontaneous tumor that occurs at high incidence in aging F344 rats, is distinct from human LGL and, therefore, MNCL data should not be used in assessing potential human health hazards.
Examination of 66 NTP F344 rat studies reveals 26 studies with significant and dramatic treatment-related decreases in the incidence of MNCL. The majority of these studies had tumor responses in non-hematopoietic tissue sites and often some indication of splenic toxicity in prechronic studies. Of the 40 studies with potential chemical treatment-related increases in MNCL, two were ultimately considered to not be carcinogenic and 38 had differing levels of evidence of carcinogenicity based on NTP determination with confirmation by peer review. For many of these 38 studies, there was discussion and debate during formal peer review indicating that for most studies with clear or some evidence of carcinogenicity for MNCL, the treatment-associated increases fell within an appropriate historical control range. While in conventional toxicology practice the concurrent control is the most appropriate comparator for assessing typical tumor responses, the high background incidence levels, variability and unpredictable nature of this tumor type makes it difficult to differentiate between a true positive and a false positive (). Due to these multiple biological factors that impact the background incidence of MNCL, we contend that analyses of F344 rat MNCL warrant more reliance on historical control data than is usual for most tumor responses. The traditional statistical approach may have an increased Type I error rate due to the biology of this unique leukemia.
Leydig cell tumors
Features of Leydig cell (LC) proliferative lesions
The two most common proliferative lesions of the testes in the F344 rats are the Leydig cell (LC) hyperplasia and LC adenoma. These tumors are age-related and appear spontaneously in this strain, and their incidence approaches 100% in 18–24-month-old F344 rats. LCTs are infrequent and less severe in the Wistar and Sprague–Dawley rats (Creasy et al. Citation2012), with less than 2% incidence rate reported. Nevertheless, according to Creasy et al. (Citation2012), the incidence of this type of tumor in different strains of Wistar rats may reach 40%, depending upon the commercial source providing the rats. LCTs in rodents are almost always benign and malignancy is rarely reported (Boorman et al. Citation1990; Cook et al. Citation1999). Nolte et al (Citation2011), who reviewed the spontaneous occurrence of LC adenomas in F344, Sprague–Dawley and Wistar rats, found only a single case of LC carcinoma in a Wistar rat.
The distinction between LC hyperplasia and adenoma is difficult, since LC adenoma is considered as a continuum of the hyperplasia, and it is not possible to distinguish between these two based on cellular characteristics alone (Steinbach et al. Citation2015). Therefore, when discussing historical data dealing with LC adenoma, it is important to describe the methodology used to define the adenoma.
The main morphological features of LC hyperplasia are the presence of focal, multifocal and diffuse collections of LCs between the seminiferous tubules, with no or minimal compression of the surrounding tissue. Most importantly, in the case of focal hyperplasia, the diameter of the lesion is smaller than or equal to three seminiferous tubules. The recently published INHAND criteria emphasized that an additional criterion for distinguishing between “normal” and LC hyperplasia is the degree of demarcation of the focal lesion from the surrounding tissue (Creasy et al. Citation2012).
Leydig cell (LC) adenoma is a proliferative lesion exceeding the diameter of three seminiferous tubules, usually with peripheral compression of adjacent seminiferous tubules. LC adenomas may contain cystic areas and less differentiated areas composed of either basophilic cells with scanty cytoplasm or elongated spindle-shaped cells.
Chemically induced proliferative LC lesions
The high incidence of LCTs in the F344 rat reflects hormone imbalance between testicular LH receptor levels and serum testosterone (Turek & Desjardins Citation1979). Cook et al. (Citation1999) described five mechanisms by which chemicals can influence hormone balance and disrupt the hypothalamic–pituitary–testes (HPT) axis leading to LC hyperplasia and tumors. These include agonists of estrogens, GnRH and dopamine receptors, androgen receptor antagonists, and inhibitors of 5α-reductase. They concluded that the agents associated with LC proliferation disturb hormonal homeostasis by endocrine, paracrine or autocrine mechanisms, leading to LC growth and/or inhibition of LC death. Of interest, it appears, that genotoxic compounds that induce LCs also operate via disruption of hormonal homeostasis, however whether that is due to DNA-reactive agents initiating LCs with LH then promoting tumor development, or via an unidentified hormonal mechanism unrelated to their genotoxic properties cannot be concluded. What is important is differentiating between genotoxic and non-genotoxic compounds since genotoxic agents, which produce LC hyperplasia or LCTs, would be considered relevant to humans. The distinction is that site concordance is not essential for genotoxic compounds as this mechanism can produce alternative tumor types in humans. However, the hormonal disruption cascade by non-genotoxic compounds that leads to LCTs in rodents is unique to rats. Initiation of a similar cascade would not occur in other organ types, and this mechanism in rats is not relevant to humans. Therefore, the induction of LCTs in F344 is not relevant for prediction of LCTs in humans and is not a mechanism that would be anticipated to produce alternative tumor types in humans.
Factors influencing the spontaneous incidence of LCTs
There is a variety of diverse factors associated with the variable incidence of LCTs. Nolte et al. (Citation2011) used the Registry of Industrial Toxicology Animal-data (RITA) database, which included 7453 male rats from different strains, in order to analyze the variables that may influence the spontaneous formation of LCTs. Some of the factors from analysis of the RITA database and from other publications are discussed below.
Strain and breeder
Commercial breeder-dependent differences in the spontaneous incidence of LC adenoma were seen in two F344 rat studies with LC adenoma incidences of 76% for one animal source and 90% for the other (Nolte et al. Citation2011). Similar breeder-related differences were also present in SD rats with incidence rates between 2.3 and 6.3%. In the Wistar rat, the mean incidence of LC adenoma was 39.9% for one breeder, while in other commercial sources of Wistar rats the incidences ranged between 2.8 and 12.5%. Taken together, these observations indicate that animal strain is the most relevant factor in influencing LCT incidences with commercial sources having a secondary but significant influence on LCT incidences for a given rat stock or strain.
Dependence on body weight
There is strong evidence that increased body weight and associated increases in prolactin serum levels reduce LCT incidence in the rat (Sharpe & McNeilly Citation1979; Nolte et al. Citation2011). However, Nolte et al. (Citation2011) pointed out that there were no obvious differences in the mean terminal body weight between animals with or without LC adenomas in the RITA database.
Age dependence
The incidence of LCTs is age dependent (Nolte et al. Citation2011). Occurrence is rare in the first year of life but LCTs are frequently observed in the male F344 rat in the 2nd year. In any case, LCTs are generally not fatal and, therefore it was concluded that their presence does not reduce the survival.
Dependence on administration route
An analysis evaluating the correlation between the route of administration and LCT incidence for specific commercial breeders revealed a higher incidence in studies with dietary admixture compared to gavage administration (Nolte et al. Citation2011). Rather than a direct effect of diet, it was suggested that the effect was dependent on year of study start given that dietary studies were conducted, on average, 6–13 years earlier than the gavage studies. Of importance here was the negative correlation between year of study start and LCT incidence for both breeders used in the analyses. Similarly, there was slightly but consistently improved survival in NTP feed studies relative to NTP inhalation studies, regardless of using either the NIH-07 or NTP-2000 diets (Haseman et al. Citation2003). Ultimately, the increased survival may influence the increased incidence of LCT reported in the feeding studies.
Individual caging versus group caging
Analysis of the RITA database did not provide a definite conclusion regarding the potential influence of the caging parameter on tumor incidence. However, Haseman et al. (Citation1997, Citation2003), who analyzed data from 22 separate 2-year feed and inhalation studies, found that differences in animal care and housing protocols may be a major factor contributing to the observed differences in LCT incidence rates. Their analysis found a relatively high incidence of pituitary gland tumors and a decreased incidence of LCTs in NTP dermal and inhalation studies (which use individual housing) compared to NTP gavage and feeding studies (which use group housing). A link between the number of animals per cage and stress levels has been proposed, with singly housed animals having a higher degree of stress (Nyska et al. Citation2002). The single-housed animals in the NTP dermal and inhalation studies would have higher stress levels leading to increased serum corticosteroids, which can impair testosterone synthesis by LCs (Nyska et al. Citation1998; Ben-Eliyahu et al. Citation1999; Nyska et al. Citation2002).
Effect of sexual activity
Weisburger et al. (Citation2002) investigated whether breeding was a possible protective factor against LCT development. Comparing virgins versus sexually active rats, the authors concluded that functionality of the testicular apparatus does not seem to influence the occurrence of this neoplasm in F344 rats, apparently contrary to some observations made 70 years earlier (Dunning & Curtis Citation1946).
Human Leydig cell tumors
In general, human testicular cancers are rare, representing 1% of all human cancers, with ∼52,000 new cases of testicular cancers estimated worldwide in 2008 (Purdue et al. Citation2005; Le Cornet et al. Citation2014). Although LCTs are the most common non-germ cell testicular tumors (Cheville Citation1999), they comprise only 1–3% of all primary testicular tumors, the majority of human testicular tumors having a germ cell origin (Kim et al. Citation1985; Vukina et al. Citation2015). In children, testicular tumors are rare; however, the relative prevalence of LCTs is much higher than in adults, reaching 8% of primary testicular tumors (Vukina et al. Citation2015).
LCTs can be diagnosed at any age, but they have two age peaks: between 5 and 10 years, which is in contrast to the rarity of the tumor in rats younger than 1 year, and in adulthood, between 20 and 60 years (Fernandez et al. Citation2004).
Almost all the cases present as a unilateral and unifocal mass, representing a discrete solid tumor that displaces the bordering parenchyma (Kim et al. Citation1985). In most of the cases, the tumors are very small, measuring less than 1 cm in diameter (Maizlin et al. Citation2004). When the size is greater than 5 cm in diameter, malignancy should be strongly suspected (Kim et al. Citation1985). Sometimes, increased endocrine function of the tumor cells can result in clinical signs and symptoms, such as precocious puberty or gynecomastia. However, clinical symptoms are not observed very commonly (Ozyavuz et al. Citation1993; Gana et al. Citation1995), and are estimated to be present in only 30% of LCT patients (Woodward et al. Citation2002).
A strong risk factor for testicular cancers in humans is undescended testis, testicular dysgenesis syndrome and infertility (Giwercman et al. Citation1987; Hoei-Hansen et al. Citation2003; Skakkebaek et al. Citation2003). Indeed, malignancy occurs in 3.5–14.5% of undescended testis (Maqdasy et al. Citation2015), although seminoma is the most prevalent type of cancer in these cases (Trabert et al. Citation2013). Nevertheless, a clear association has not been found between LCTs and undescended testis, and the etiology of LCTs in humans is still largely unknown (Tsitouridis et al. Citation2014).
Macroscopically, LCTs are well-defined solid tumors, which occasionally demonstrate lobulated masses. They have a variable coloration, ranging from yellow to brown or gray-white, depending on the lipid content (Kim et al. Citation1985; Tsitouridis et al. Citation2014). Histopathologically, these tumors are characterized by sheets or septated lobules of minimally pleomorphic low N:C ratio cells. They have an eosinophilic cytoplasm, cytologically bland nuclei and prominent nucleoli (Kim et al. Citation1985; Vukina et al. Citation2015). Zona fasciculata-type vacuolated cells are occasionally seen inside the tumor, and can sometimes even be the predominant cells (Kim et al. Citation1985). Other features that have been described less commonly include spindle cell cytology, adipose metaplasia and osseous metaplasia (Ulbright et al. Citation2002). While Reinke crystals are considered a characteristic finding in LCTs, they are present in only the minority of the cases (Kim et al. Citation1985). Sometimes a thin fibrous capsule can be seen separating the tumor from the compressed normal testicular tissue (Unluer et al. Citation1990). Almost all of the cells of LCTs in humans will be positive for inhibin, Mart-1 and calretinin stains (Vukina et al. Citation2015).
Similar to rats, human LCTs are generally benign (Farkas et al. Citation2000). Therefore, some studies suggest conservative or testis-sparing surgery instead of orchidectomy, especially in young patients with a single testicle (Masoudi et al. Citation1999; Giannarini et al. Citation2010). While in children LCTs are always benign, in adults approximately 10% have a malignant phenotype with metastatic potential, especially in older men (median age of 62.1 years) (Gonzalez et al. Citation2007; Hendry et al. Citation2015). The distinction of malignant potential is based on specific histopathological characteristics that include: large tumor size (>5 cm), cytological atypia, increased mitotic activity, increased MIB-1 expression, necrosis, vascular invasion, infiltrative margins, extension beyond the testicular parenchyma and DNA aneuploidy (Albers et al. Citation2011). The most common sites of metastasis are the regional lymph nodes, followed by the liver, lungs and bone (Kim et al. Citation1985; Tsitouridis et al. Citation2014).
In general, LCTs in humans share many similarities to their counterparts in rats, having a relatively benign nature and similar morphologic and clinical characteristics. Nevertheless, in humans, the tumor is very rare, in contrast to the much higher prevalence in F344 rats. Furthermore, in humans, LCT has two age peaks, while in F344 rats there is a gradually increasing incidence with age.
NTP studies with an LCT response
Features of seven NTP studies in F344 rats that have an increased LCT response to treatment are summarized in and statistical analyses of these LCT responses are provided in Supplemental Table S2. Of these seven studies, one had clear evidence of carcinogenicity for LCTs (Isoprene, NTP TR486) and the remaining six were judged to have equivocal evidence of carcinogenicity for LCTs. For five of the seven studies, the route of administration was inhalation, with this relatively high frequency of association possibly linked to the stress of single cage housing as discussed previously (i.e. the single caging environment ultimately increases the power of the inhalation studies to detect treatment related effects). Interestingly, in all five of these inhalation studies a kidney tumor response was reported, but a biological connection between the two responses is not apparent. In the case of kava kava extract (NTP TR571), a corn oil gavage study, the equivocal evidence of carcinogenicity represents the only tumor response in the study.
Table 8. NTP studies that had a Leydig cell tumor response.
The background incidence of LCTs in the F344 rat is exceptionally high, making it practically impossible to assess testicular carcinogenic potential using this strain of rat (). The NTP historical control rate of LCTs has been constant over time ranging from 54% to as high as 98%. The fact that five of the seven NTP studies with at least an equivocal treatment-related LCT response are from inhalation studies is noteworthy because the spontaneous incidence of LCTs in inhalation study controls is lower than in other routes of test article administration, thereby allowing identification of potential treatment-related increases based on statistical evidence (). This supports that strains with high background incidences of tumor types do not have enough statistical power to detect effects, using common bioassay protocols. To obtain robust data one would need to either increase the animal numbers used or switch the strain of animal used.
It is of interest to note that in one study male F344 rats as well as male Wistar rats were exposed to pyridine (NTP TR470), and there was equivocal evidence of carcinogenicity for LCTs in the Wistar but not the F344 rat. Since the control group incidence of LCT was 85% in the F344 rat study, the lack of evidence for LCT carcinogenicity in the F344 rat study is possibly due to the reduced power of the F344 rat to detect true positives due to the high background incidence.
Taken together, it is obvious that analysis of LCT data taken from NTP studies should be judged very carefully. First, the background incidence of LCT in historical controls can reach 100% in some cases; second, there is a proposed link between cage-related stress conditions in inhalation studies and lower incidence of LCT that confounds determinations of chemical influence; and, finally, the mechanism for LCT induction in rats differs from the mechanism for induction of LCTs in humans. Thus an increased frequency of LCTs in F344 rats is not predictive of LCT induction in humans or useful for human health risk assessment. The mechanism of LCT induction in rats by non-genotoxic compounds is a testis-specific process that does not occur in humans, and, therefore, is not anticipated to be relevant for other potential tumor types in humans.
Conclusions
The high spontaneous incidence of LCTs in F344 rats has made this bioassay model of little practical use in identifying potential testicular carcinogenic responses. Inhalation studies may be a questionable exception due to a stress response in single cage housing, but even then a putative LCT response in NTP studies has ranked only as equivocal evidence of testicular carcinogenicity. In contrast to the age-related high incidence in F344 rats, human LCTs are rare and have two age peaks of occurrence. Thus, the F344 LCT is not relevant to human testicular cancer, except possibly for the rare cases of men with genetic susceptibility to develop LCTs, such as individuals harboring somatic mutations in the LHCG-R or GNAS genes or germline mutations in FH (Libé et al. Citation2012). We believe that use of the F344 rat strain has long been recognized as not being useful for identification of testicular carcinogens and applaud the NTP for finally switching to a different rat for toxicity and carcinogenicity studies.
Tunica vaginalis mesothelioma
Features and pathogenesis of tunica vaginalis mesotheliomas in F344 rats
An extensive published review of TVM induction in rats is available (Maronpot et al. Citation2009). Spontaneous TVM in the rat is an age-associated tumor with background incidences ranging from 0.2% to 5%, and with most descriptions in the literature based on observations in F344 rats. Histomorphological features of tunica vaginalis and peritoneal mesotheliomas are similar in all species, including humans (Ilgren & Wagner, Citation1991; Ilgren, Citation1993). In rats, these tumors typically start as single-layered collections of hyperplastic mesothelium overlying a thin fibrovascular stroma initially localized in the tunica vaginalis. With continued growth they can form small papillary projections of hyperplastic mesothelium as well as multilayered sheets and nests of cuboidal to polygonal cells with round nuclei and single prominent nucleoli. The mesothelioma growths can form glandular and tubular structures, can have a sarcomatous phenotype, and may form cystic structures lined by flattened mesothelium. Rat TVMs may be classified as epithelial, sarcomatous or mixed, consistent with mesothelioma classification in humans. Implant metastasis, induced by transcoelomic spread from the primary tumor in the tunica vaginalis, can be seen throughout the peritoneal cavity. Features of malignancy include pleomorphism, cytological atypia and local invasiveness but even without these features TVMs are generally regarded as malignant even if confined to the scrotal sac. Immunohistochemical features of mesotheliomas are useful in differentiating them from adenocarcinomas (Maronpot et al. Citation2009). Pathological features of treatment-associated TVM in the F344 rat are similar to spontaneous cases but generally have a reduced latency and greater extension into the peritoneal cavity. They are generally less invasive and less pleomorphic than mesotheliomas associated with exposure to asbestos or nanotubes (Maronpot et al. Citation2009).
Most of what is known about TVMs is from the uniquely sensitive F344 rat. Both spontaneous and xenobiotic-induced TVMs are closely associated with and most likely secondary to LCTs, a common spontaneous tumor in F344 rats. Hormone imbalance is a likely key event associated with both spontaneous and treatment-induced TVMs (Turek & Desjardins Citation1979; Tanigawa et al. Citation1987; Shipp et al. Citation2006). The high incidence of LCTs in the F344 rat also reflects hormone imbalance between testicular LH receptors levels and serum testosterone and is potentially causally linked to development of TVMs in this rat (Turek & Desjardins Citation1979). One hypothesis is that the hormone imbalance associated with LCTs exposes the tunica vaginalis compartment to transudates containing altered androgen levels leading to release of growth factors, mesothelial mitogenesis and development of TVM (Karpe et al. Citation1982; Gerris & Schoysman Citation1984). An alternative hypothesis implicates mechanical pressure from the LCTs, leading to tunica vaginalis mesothelial cell expression of autocrine growth factor stimulating mitogenesis (Gerwin et al. Citation1987; Tanigawa et al. Citation1987; Versnel et al. Citation1988). In response to shearing forces and pressure from an adjacent testis enlarged by the presence of a LCT, mesothelial cells produce autocrine growth factors that lead to mitogenesis.
Although hormone imbalance and mechanical pressure are likely key events in the genesis of TVM, xenobiotics or their metabolites can potentially reach the tunica vaginalis mesothelium and directly or indirectly lead to mesothelial mitogenesis and development of TVM. Oxidative stress secondary to reactive oxygen species is considered an important mediator of asbestos-associated mesotheliomas and could play a role in the development of TVM (Attanoos & Gibbs Citation1997; Schurkes et al. Citation2004). Cell cycle alterations in testicular mesothelium have been seen following subchronic exposure to acrylamide (Lafferty et al. Citation2004), a known xenobiotic associated with induction of TVM in F344 rats.
In addition to 17 NTP studies with a treatment-related increased incidence in TVM, there are reports in the literature that additional xenobiotics have caused TVM in F344 rats from different commercial colonies (Maronpot et al. Citation2009). This observation reflects the sensitivity of the F344 rat from different commercial sources to development of TVM. The strain specificity of the F344 rat to develop TVM following xenobiotic exposure by other than a peritoneal injection route is unique to the F344 rat and is not seen in other strains or stocks of rats, even following sustained increased LH levels (Prentice et al. Citation1992). Furthermore, in hazard identification studies conducted by NTP and others with parallel studies in mice, peritoneal mesothelioma responses have never been seen in mice or in female rats.
TVM in humans
Just like in rats, the tunica vaginalis in humans is of mesothelial origin, similar to the pleura, peritoneum and pericardium. It is derived from the processus vaginalis, an outpouching of the peritoneum, which descends to cover the testis in the scrotum. Unlike in the rat, the cranial end of the processus vaginalis closes, forming a closed cavity, which is not connected to the peritoneal cavity (Woodward et al. Citation2003; Hassan & Alexander Citation2005; Garriga et al. Citation2009; Maronpot et al. Citation2009). Several mesothelial lesions can arise from the paratesticular area, and include reactive mesothelial hyperplasia, mesothelial cysts, adenomatoid tumor, well-differentiated papillary mesothelioma and malignant mesothelioma (Erdogan et al. Citation2014).
Most of the mesotheliomas in humans develop in the pleura, comprising 68–85% of all malignant mesothelioma cases. Of the remainder, 9.1–24.1% of malignant mesotheliomas cases develop in the peritoneum (Chen & Hsu Citation2009). TVM is the rarest mesothelioma, representing only 0.3–5% of all mesotheliomas (Chekol & Sun Citation2012). Since its first description in 1957 (Barbera & Rubino Citation1957), approximately 250 cases have been reported (Chen & Hsu Citation2009; Bisceglia et al. Citation2010; Chekol & Sun Citation2012; Mensi et al. Citation2012; Bandyopadhyay et al. Citation2015), and in Italy, the standardized incidence rate for TVM is 0.2 cases per million (Bisceglia et al. Citation2010; Marinaccio et al. Citation2010). Clinically, the tumor usually presents as recurrent or enlarging hydrocele, and not as a scrotal mass. Other less common presentations include inguinal hernia, epididymitis, spermatocele, testicular torsion and testicular traumatic injury (Plas et al. Citation1998; Hamm et al. Citation1999; Kato et al. Citation2012; Yen et al. Citation2012; Segura-Gonzalez et al. Citation2015).
While exposure to asbestos has been linked to TVM development, it is reported in only 30–40% of the cases (Guney et al. Citation2007; Ikegami et al. Citation2008). Trauma, herniorrhaphy, ionizing radiation and long-standing hydrocele are additional reported risk factors (Peterson et al. Citation1984; Goel et al. Citation2008). Like pleural mesotheliomas (Peterson et al. Citation1984), TVMs have been suggested to develop following chronic inflammatory condition, and recurrent epididymitis has been proposed as a risk factor for the development of TVM (Yen et al. Citation2012). It should be noted that while many studies have shown the presence of SV40 in pleural mesotheliomas in humans, its presence in TVM is not well documented, and was negative in several reports (Xiao et al. Citation2000; Erdogan et al. Citation2014).
A wide age range has been reported for this tumor, ranging between 7 and 91 years, with the mean age at diagnosis being 53.5 years (Boyum & Wasserman Citation2008; Chekol & Sun Citation2012). Only 10% of the cases occur in patients younger than 25 years old (Plas et al. Citation1998).
The molecular pathogenesis for mesothelioma in general is still not entirely clear. A homozygous deletion of the 9q21 locus is one of the most common genetic alterations found in mesothelioma (Husain et al. Citation2013), and recently germline mutation in BAP1 (BRCA 1 associated protein) was suggested to predispose to malignant mesothelioma (Testa et al. Citation2011). Additional genomic changes include losses in 1p36, 22q12 and 14q32 and gains in 5p and 7p (Kivipensas et al. Citation1996; Takeda et al. Citation2012).
Three clinicopathologic types of malignant mesotheliomas of the male genital tract have been described: diffuse tubulopapillary mesothelioma, well-differentiated papillary mesothelioma and multicystic mesothelioma (Rajan et al. Citation2013). Macroscopically, the tumor appears as a firm, solid, cystic, yellow or white pearly mass. Thickening of the tunica vaginalis is commonly observed, and nodules of different sizes can cover it. The tumor can invade adjacent structures, such as the testicular parenchyma, epididymis and spermatic cord (Segura-Gonzalez et al. Citation2015).
Microscopically, three subtypes of malignant mesotheliomas are recognized: epithelial, mesenchymal (or sarcomatous) and mixed (Hassan & Alexander Citation2005). Most of the tumors are of the epithelial type, with papillary, tubulopapillary and solid patterns. The mixed type is the second in frequency, and the pure sarcomatous type is very rare (Jones et al. Citation1995; Plas et al. Citation1998). The neoplastic cells are typically cuboid, with scant eosinophilic cytoplasm (Plas et al. Citation1998; Chekol & Sun Citation2012). Mesothelial hyperplasia of the tunica may sometimes be extensive, and mimic malignant mesothelioma, especially of the epithelial type. Such hyperplasia may result from stubborn or repetitive serosal injury inflammation in hydroceles and inguinal hernia sacs (Churg Citation2003; Lee et al. Citation2014). Biomarkers have been utilized to differentiate between these two conditions, including p53, Ki-67 and GLUT-1, but no antibody has had an absolute discriminatory value (Attanoos et al. Citation2003; Kato et al. Citation2007; Taheri et al. Citation2008; Lagana et al. Citation2012; Lee et al. Citation2013).
Immunohistochemically, TVMs show similar reaction as pleural mesotheliomas, with positive staining to calretinin antibody, Wilms tumor (WT1), epithelial membrane antigen, thrombomodulin and cytokeratin 7. The tumor is negative to cytokeratin 20 and carcinoembryonic antigen, and variable expression of cytokeratin 5/6 is evident (Amin et al. Citation1995; Winstanley et al. Citation2006; Pacheco et al. Citation2009).
Metastasis is reported in less than 15% of cases, and 35.3% of the patients that undergo staging lymphadenectomy do not have any tumors (Plas et al. Citation1998; Abe et al. Citation2002; Chekol & Sun Citation2012). The first line of treatment is radical orchidectomy, but unfortunately, local recurrence is common due to the aggressive nature of the tumor, occurring in 53% of the cases, usually within 2 years of diagnosis (Boyum & Wasserman Citation2008). Median overall survival ranges from 14 to 23 months in most of the series (Segura-Gonzalez et al. Citation2015). The most common sites for metastasis are the retroperitoneal lymph nodes, followed by inguinal and iliac lymph nodes. When distant metastasis occurs, it commonly involves the lung, liver and rarely bones (Bandyopadhyay et al. Citation2015). History positive for asbestos exposure is significantly associated with shorter interval before tumor recurrence (Plas et al. Citation1998).
Since the tunica vaginalis in the adult human does not directly connect to the peritoneal cavity, human TVMs originate only from the scrotal sac where they are locally more invasive and metastatic compared to TVM in the F344 rat (Guney et al. Citation2007). Furthermore, the rarity of LCTs in humans indicates an alternative mechanism for TVM induction in humans in comparison to the likely primary mode of action in the F344 rat (secondary to LCTs for TVM induction). Consequently, the specificity of the TVM response in the male F344 rat is not likely to be relevant to other species, including humans.
NTP studies with increased incidences of TVMs
Fourteen of 17 NTP studies with a treatment-related increased incidence of TVM in F344 rats had clear evidence of carcinogenicity or were considered positive for four studies completed before NTP began categorizing studies using levels of evidence of carcinogenicity (). Statistical analyses are provided in Supplemental Table S3. Thirteen studies were multi-site carcinogens with two or more additional tumor responses in tissues other than the peritoneal mesothelium while in three studies (methyleugenol (NTP TR491), o-nitrotoluene (NTP TR504) and o-toluidine hydrochloride (NTP TR153), TVM was the only tumor response. Pentachlorophenol (NTP TR 483) had some evidence of carcinogenicity for TVM with a questionable nasal tumor response. The positive TVM responses were typically clear and robust with the exception of two studies [1-bromopropane (NTP TR564) and nitrofurazone (NTP TR337)] with equivocal evidence of carcinogenicity where the TVM response was only one tumor above the upper incidence of the control range. TVMs were all considered malignant, started in the tunica vaginalis mesothelium, and usually spread to multiple sites within the peritoneal cavity. Among the 17 studies with a TVM response, there were three studies that incorporated stop exposures at 3 (o-nitrotoluene) or 12 (methyleugenol and pentachlorophenol) months, usually at a higher dose, and had increased incidences of TVM. For pentachlorophenol, the TVM response was present in the stop exposure group but not in the main study groups.
Table 9. NTP studies that had a tunica vaginalis mesothelioma (TVM) response.
Conclusions
TVMs are rarely seen in male control F344 rats making detection of increases relatively easy to assess. Even in studies with up to a 90% incidence of TVM in male F344 rats, there was never a mesothelioma response in females, including the cytembena study (NTP TR207) where the route of administration was intraperitoneal and accompanied by peritoneal inflammation in both sexes. Furthermore, human TVMs are found only in the scrotal sac, and bear a more invasive and metastatic potential when compared to TVM in the F344 rat. Moreover, F344 rat TVM is considered a secondary event to the common LCT, and the rarity of LCTs in humans suggests that a different mechanism underlies development of this tumor in humans. This supports the contention that the TVM response is a rat strain and gender-specific response characteristic of the F344 rat without significant relevance to human health.
Perspective on the legacy of the F344/N rat
Cancer bioassays conducted in F344 rats have been used by the NTP since 1978 for hazard identification. In this context, some have regarded a positive tumor response as evidence of potential carcinogenic hazard at any site in humans. In our present evaluation, we have assessed the relevance of MNCL, LCTs and TVM to human cancer risk based on tissue site-concordance. The topic of tissue site-concordance between tumor responses in rodents and expected tumor responses in humans has long been contested in the literature and been the topic of scientific debate. While we cannot easily resolve this debate here, we point out our rationale for favoring site-concordance in our assessment of the human relevance of F344 rat MNCL, LCT and TVM bioassay responses.
When evaluating risk assessment, a higher level of confidence has generally been given to animal data when there is site-concordance with tumor data (EPA Citation2005). This higher level of confidence derives from experimental evidence indicating “that there are more physiological, biochemical and metabolic similarities between laboratory animals and humans than there are differences” (Rall et al. Citation1987). Tissue site concordance between animals and humans exposed to the same xenobiotics has been shown for many chemicals including aflatoxins B1, vinyl chloride, some arsenicals, asbestos, benzene and diethylstilbesterol among others (Tomatis et al. Citation1989; Goodman & Wilson Citation1991). Even though target sites for carcinogenesis are often concordant between animals and humans (Tomatis et al. Citation1989; Huff Citation1994), admittedly concordance by site is not universal. Outcomes in any given animal study are influenced by multiple factors, including experimental design, exposure levels, species, strain, sex, route of exposure, duration of exposure, metabolism, diet, pathology, DNA reactivity of the test agent and mode of action leading to a cancer response (Maronpot et al. Citation2004). Yet there are enough examples of tissue site concordance to favor its use in assessing human carcinogenic risk. Furthermore, a fundamental premise for the contemporary approach of employing the mode of action/human relevance framework to assess carcinogenic hazard to humans is based on tissue site concordance between animals and humans (Cohen et al. Citation2004; EPA Citation2005) and even the premise of toxicokinetic and toxicodynamic modeling is based on tissue site concordance (EPA Citation2005). Consequently, we have assessed the human relevance of F344 rat MNCL, LCT and TVM based on tissue site-concordance since there is no biologically compelling reason to expect that any potential mechanisms inducing these specific tumors in F344 rats would lead to human cancer at an entirely different tissue site.
Unfortunately, it became clear from the NTP experience over time, as highlighted in our review, that the F344/N rat strain harbors significant health issues which critically affect the integrity of certain organs, as well as the life-span of these animals. These and other health issues in this rat strain led to the NTP decision to switch to a more appropriate strain of rats for their toxicity and carcinogenicity studies (King-Herbert et al. Citation2010). Specifically, we highlight three cancers that are uniquely common in the F344/N rat strain. MNCL and LCT occur with a high background incidence and for MNCL the high background incidence is extremely variable. The very high spontaneous incidence of LCT prevents the use of this strain to reliably predict a potential testicular carcinogenic effect. TVM occurs rarely in rat carcinogenicity studies but it is unique to the F344 rat with a biologically plausible association with the high background incidence of LCT. In addition, we have provided documentation from contemporary literature indicating important pathobiological differences between these F344 rat tumors and similar target tissue-tumor responses in humans. We believe that rodent tumor responses are potentially relevant to human health risk unless there are good reasons to believe this is not the case.
For concluding on the relevance of study findings to humans, two main factors are considered: first, whether the observed tumor is related to the chemical exposure and second, whether the chemically induce tumor in the rodent model is relevant for human risk assessment. As discussed in depth previously in this review, we summarize below how data for these tumor types relate to these main factors.
For MNCL, there are statistical considerations for determining whether or not changes in incidence are related to chemical exposure as well as considerations of whether chemically induced alterations in this tumor type are relevant to humans. First, for determining whether a chemical does influence incidence of this tumor type it is important to compare values to the appropriate historical control values and to use more stringent statistical criteria such as those outlined in Thomas et al. (Citation2007) (p < 0.01 and p < 0.005 for a trend test) for tumor responses assessed against a high background frequency as for MNCL. The etiology of the most similar human LGL leukemia (ANKCL) to F344 rat MNCL is related to infection with Epstein–Barr virus, and is not associated with drug or chemical exposure. Though the specific mode of action for the F344 rat MNCL is not known, it is not associated with a viral etiology. There is a qualitative difference in how human ANKCL leukemia and F344 rat MNCL are initiated; therefore, despite some commonalities between the pathologies of these tumors, MNCL is not a model for human LGL leukemia. In addition, there is no evidence for a genotoxic mechanism of action for MNCL induction; rather it is due to a yet unknown secondary mechanism. These data indicate there is not a concern for prediction of a site-concordant or non-concordant human relevant tumor type.
For the LCTs, the high background incidence makes it difficult to determine if the tumor incidence observed in the F344 rat is related to chemical exposure. The mode of action for the induction of the LCTs is unique to F344 rats. There is no equivalent mode of action in humans; therefore, non-genotoxic chemicals operating through this mode of action in F344 rats would not influence human carcinogenicity incidence.
Development of TVM in F344 rats is strain- and gender-specific and closely tied to the presence of LCTs. Since induction of LCTs in F344 rats is not relevant to human cancer risk, the development of TVMs in this rat strain is also not relevant to human cancer risk.
Given the material presented in this retrospective review, we contend that MNCL, LCT, and TVM responses in F344 rat carcinogenicity studies lack relevance in predicting human carcinogenicity.
Supplemental_Content.zip
Download Zip (299.2 KB)Acknowledgements
The authors gratefully acknowledge the comments and suggestions of the four reviewers selected by the Editor and unknown to the authors.
Declaration of interest
Dr Robert R. Maronpot: I retired from the National Institute of Environmental Health Sciences and National Toxicology Program in 2007 and have been a private consultant since that time. It was my idea to prepare this manuscript with my colleagues and ExxonMobil agreed to compensate us for our time spent researching and writing the paper. I have no responsibility for or any financial interest in any of the chemicals mentioned in this manuscript and no financial interests in any way to ExxonMobil.
Dr Abraham Nyska: I am an adjunct faculty member of the Sackler School of Medicine, Tel Aviv University and Timrat, Israel and agreed to participate in preparation of this manuscript at the invitation of Dr Robert R. Maronpot. I have no financial interest in Exxon Mobil and am not consulting elsewhere with Exxon Mobil on other projects. My participation in writing this review was not influenced by Exxon Mobil in any way.
Dr Yuval Ramot: I am a faculty member of Hadassah-Hebrew University Medical Center, Jerusalem, Israel and agreed to participate in preparation of this manuscript at the invitation of Dr Robert R. Maronpot. I have no financial interest in Exxon Mobil and am not consulting elsewhere with Exxon Mobil on other projects. My participation in writing this review was not influenced by Exxon Mobil in any way.
Dr Jennifer E. Foreman: I am employed by ExxonMobil Biomedical Sciences, Inc., a separately incorporated, wholly owned affiliate of Exxon Mobil Corporation. I agreed to participate in preparation of this manuscript at the invitation of Dr Robert R. Maronpot.
The authors alone are responsible for the preparation and content of this article.
Notes
1. Note: MNCL is now considered a large granular lymphocytic (LGL) leukemia but has traditionally been called MNCL in NTP bioassays. We will continue to refer to it as MNCL to avoid confusion with listings in the NTP database.
2. A review of NTP MNCL studies with respect to human health risk was previously published (Thomas et al. Citation2007). This study includes the studies covered in that 2007 publication plus six additional F344 rat studies with a MNCL response completed prior to the NTP decision to no longer use the F344 rat.
References
- Abe K, Kato N, Miki K, Nimura S, Suzuki M, Kiyota H, Onodera S, Oishi Y. 2002. Malignant mesothelioma of testicular tunica vaginalis. Int J Urol. 9:602–603.
- Albers P, Albrecht W, Algaba F, Bokemeyer C, Cohn-Cedermark G, Fizazi K, Horwich A, Laguna MP. 2011. EAU guidelines on testicular cancer: 2011 update. Eur Urol. 60:304–319.
- Alekshun TJ, Tao J, Sokol L. 2007. Aggressive T-cell large granular lymphocyte leukemia: a case report and review of the literature. Am J Hematol. 82:481–485.
- Amin KM, Litzky LA, Smythe WR, Mooney AM, Morris JM, Mews DJ, Pass HI, Kari C, Rodeck U, Rauscher FJ, III, et al. 1995. Wilms' tumor 1 susceptibility (WT1) gene products are selectively expressed in malignant mesothelioma. Am J Pathol. 146:344–356.
- Attanoos RL, Gibbs AR. 1997. Pathology of malignant mesothelioma. Histopathology. 30:403–418.
- Attanoos RL, Griffin A, Gibbs AR. 2003. The use of immunohistochemistry in distinguishing reactive from neoplastic mesothelium. A novel use for desmin and comparative evaluation with epithelial membrane antigen, p53, platelet-derived growth factor-receptor, P-glycoprotein and Bcl-2. Histopathology. 43:231–238.
- Bandyopadhyay A, Bhattacharya S, Konar K. 2015. Preoperative cytological diagnosis of malignant mesothelioma of tunica vaginalis. Diagn Cytopathol. 43:850–854.
- Barbera V, Rubino M. 1957. Papillary mesothelioma of the tunica vaginalis. Cancer. 10:183–189.
- Ben-Eliyahu S, Page GG, Yirmiya R, Shakhar G. 1999. Evidence that stress and surgical interventions promote tumor development by suppressing natural killer cell activity. Int J Cancer. 80:880–888.
- Bisceglia M, Dor DB, Carosi I, Vairo M, Pasquinelli G. 2010. Paratesticular mesothelioma. Report of a case with comprehensive review of literature. Adv Anat Pathol. 17:53–70.
- Boorman G, Chapin R, Mitsumori K. 1990. Testis and epididymis. In: Boorman G, Eustis S, Elwell M, Montgomery C, Jr, MacKenzie W, editors. Pathology of the Fischer rat. Reference and Atlas. New York: Academic Press. p. 405–418.
- Boyum J, Wasserman NF. 2008. Malignant mesothelioma of the tunica vaginalis testis: a case illustrating Doppler color flow imaging and its potential for preoperative diagnosis. J Ultrasound Med. 27:1249–1255.
- Caldwell DJ. 1999. Review of mononuclear cell leukemia in F-344 rat bioassays and its significance to human cancer risk: a case study using alkyl phthalates. Regul Toxicol Pharmacol. 30:45–53.
- Cameron TP, Hickman RL, Kornreich MR, Tarone RE. 1985. History, survival, and growth patterns of B6C3F1 mice and F344 rats in the National Cancer Institute Carcinogenesis Testing Program. Fundam Appl Toxicol. 5:526–538.
- Chan J, Jaffe E, Ralfkiaer E, Ko JH. 2008b. Aggressive NK-cell leukemia. In: Swerdlow S, Campo E, Harris N, Jaffe E, Pileri S, Stein H, Thiele J, Vardiman J, editors. World Health Organization classification of tumours of hematopoietic and lymphoid tissues. Lyon: IARC Press. p. 276–277.
- Chan W, Foucar K, Morice W, Catovsky D. 2008a. T-cell large granular lymphocytic leukemia. In: Swerdlow S, Campo E, Harris N, Jaffe E, Pileri S, Stein H, Thiele J, Vardiman J, editors. World Health Organization classification of tumours of hematopoietic and lymphoid tissues. Lyon: IARC Press. p. 272–273.
- Chekol SS, Sun CC. 2012. Malignant mesothelioma of the tunica vaginalis testis: diagnostic studies and differential diagnosis. Arch Pathol Lab Med. 136:113–117.
- Chen JL, Hsu YH. 2009. Malignant mesothelioma of the tunica vaginalis testis: a case report and literature review. Kaohsiung J Med Sci. 25:77–81.
- Cheung MM, Chan JK, Wong KF. 2003. Natural killer cell neoplasms: a distinctive group of highly aggressive lymphomas/leukemias. Semin Hematol. 40:221–232.
- Cheville JC. 1999. Classification and pathology of testicular germ cell and sex cord-stromal tumors. Urol Clin North Am. 26:595–609.
- Churg A. 2003. Paratesticular mesothelial proliferations. Semin Diagn Pathol. 20:272–278.
- Cohen SM, Klaunig J, Meek ME, Hill RN, et al. 2004. Evaluating the human relevance of chemically induced animal tumors. Toxicol Sci. 78:181–186.
- Coleman GL, Barthold W, Osbaldiston GW, Foster SJ, Jonas AM. 1977. Pathological changes during aging in barrier-reared Fischer 344 male rats. J Gerontol. 32:258–278.
- Cook JC, Klinefelter GR, Hardisty JF, Sharpe RM, Foster PM. 1999. Rodent Leydig cell tumorigenesis: a review of the physiology, pathology, mechanisms, and relevance to humans. Crit Rev Toxicol. 29:169–261.
- Creasy D, Bube A, de Rijk E, Kandori H, Kuwahara M, Masson R, Nolte T, Reams R, Regan K, Rehm S, et al. 2012. Proliferative and nonproliferative lesions of the rat and mouse male reproductive system. Toxicol Pathol. 40:40S–121S.
- Davey FR, Moloney WC. 1970. Postmortem observations on Fischer rats with leukemia and other disorders. Lab Invest. 23:327–334.
- Dieter MP, Jameson CW, French JE, Gangjee S, Stefanski SA, Chhabra RS, Chan PC. 1989. Development and validation of a cellular transplant model for leukemia in Fischer rats: a short-term assay for potential anti-leukemic chemicals. Leuk Res. 13:841–849.
- Dieter MP, Jameson CW, Maronpot RR, Langenbach R, Braun AG. 1990. The chemotherapeutic potential of glycol alkyl ethers: structure-activity studies of nine compounds in a Fischer-rat leukemia transplant model. Cancer Chemother Pharmacol. 26:173–180.
- Dieter MP, Maronpot RR, French JE. 1985. Comparison of the morphology and enzyme activity of mononuclear cells from Fischer 344 rats with either spontaneous or transplanted leukemia. Cancer Res. 45:4301–4307.
- Dunnick JK, Eustis SL, Huff JE, Haseman JK. 1989. Two-year toxicity and carcinogenicity studies of ampicillin trihydrate and penicillin VK in rodents. Fundam Appl Toxicol. 12:252–257.
- Dunning WF, Curtis MR. 1946. The respective roles of longevity and genetic specificity in the occurrence of spontaneous tumors in the hybrids between two inbred lines of rats. Cancer Res. 6:61–81.
- Dunning WF, Curtis MR. 1957. A transplantable acute leukemia in an inbred line of rats. J Natl Cancer Inst. 19:845–853.
- Elwell MR, Dunnick JK, Hailey JR, Haseman JK. 1996. Chemicals associated with decreases in the incidence of mononuclear cell leukemia in the Fischer rat. Toxicol Pathol. 24:238–245.
- EPA. 2005. Guidelines for carcinogen risk assessment. EPA/630/P=03/001F. Washington, DC: Environmental Protection Agency. p. 166.
- Erdogan S, Acikalin A, Zeren H, Gonlusen G, Zorludemir S, Izol V. 2014. Well-differentiated papillary mesothelioma of the tunica vaginalis: a case study and review of the literature. Korean J Pathol. 48:225–228.
- Farkas LM, Szekely JG, Pusztai C, Baki M. 2000. High frequency of metastatic Leydig cell testicular tumours. Oncology. 59:118–121.
- Fernandez GC, Tardaguila F, Rivas C, Trinidad C, Pesqueira D, Zungri E, San Miguel P. 2004. Case report: MRI in the diagnosis of testicular Leydig cell tumour. Br J Radiol. 77:521–524.
- Friedman MA, Dulak LH, Stedham MA. 1995. A lifetime oncogenicity study in rats with acrylamide. Fundam Appl Toxicol. 27:95–105.
- Frith CH. 1988. Morphologic classification and incidence of hematopoietic neoplasms in the Sprague-Dawley rat. Toxicol Pathol. 16:451–457.
- Frith CH, Ward JM, Chandra M. 1993. The morphology, immunohistochemistry, and incidence of hematopoietic neoplasms in mice and rats. Toxicol Pathol. 21:206–218.
- Gana BM, Windsor PM, Lang S, Macintyre J, Baxby K. 1995. Leydig cell tumour. Br J Urol. 75:676–678.
- Garriga V, Serrano A, Marin A, Medrano S, Roson N, Pruna X. 2009. US of the tunica vaginalis testis: anatomic relationships and pathologic conditions. Radiographics. 29:2017–2032.
- Gentile TC, Uner AH, Hutchison RE, Wright J, Ben-Ezra J, Russell EC, Loughran TP, Jr. 1994. CD3+, CD56+ aggressive variant of large granular lymphocyte leukemia. Blood. 84:2315–2321.
- Gerris J, Schoysman R. 1984. Tunica vaginalis fluid (TVF): hormonal composition. Assessment of testosterone, 5 alpha-dihydrotestosterone, delta 4-androstenedione, dehydroepiandrosteronesulphate, estradiol, LH, FSH and PRL. Acta Eur Fertil. 15:205–216.
- Gerwin BI, Lechner JF, Reddel RR, Roberts AB, Robbins KC, Gabrielson EW, Harris CC. 1987. Comparison of production of transforming growth factor-beta and platelet-derived growth factor by normal human mesothelial cells and mesothelioma cell lines. Cancer Res. 47:6180–6184.
- Giannarini G, Dieckmann KP, Albers P, Heidenreich A, Pizzocaro G. 2010. Organ-sparing surgery for adult testicular tumours: a systematic review of the literature. Eur Urol. 57:780–790.
- Giwercman A, Grindsted J, Hansen B, Jensen OM, Skakkebaek NE. 1987. Testicular cancer risk in boys with maldescended testis: a cohort study. J Urol. 138:1214–1216.
- Goel A, Agrawal A, Gupta R, Hari S, Dey AB. 2008. Malignant mesothelioma of the tunica vaginalis of the testis without exposure to asbestos. Cases J. 1:310.
- Gonzalez M, Merlani P, Egger JF, Pugin F, Morel P. 2007. Hemorrhagic shock caused by rupture of an intra-abdominal leydig cell tumour: case report. Case Rep Gastroenterol. 1:53–58.
- Goodman D, Boorman G, Strandberg J. 1985. Selection and use of the B6C3F1 mouse and F344 rat in long-term assays for carcinogenicity. In: Millman H, Weisburger E, editors. Handbook of carcinogen testing. New Jersey: Noyes Publications. p. 282–320.
- Goodman DG, Ward JM, Squire RA, Chu KC, Linhart MS. 1979. Neoplastic and nonneoplastic lesions in aging F344 rats. Toxicol Appl Pharmacol. 48:237–248.
- Goodman G, Wilson R. 1991. Predicting the carcinogenicity of chemicals in humans from rodent bioassay data. Environ Health Perspect. 94:195–218.
- Guney N, Basaran M, Karayigit E, Muslumanoglu A, Guney S, Kilicaslan I, Gulbarut S. 2007. Malignant mesothelioma of the tunica vaginalis testis: a case report and review of the literature. Med Oncol. 24:449–452.
- Hamm M, Rupp C, Rottger P, Rathert P. 1999. [Malignant mesothelioma of the tunica vaginalis testis]. Chirurg. 70:302–305.
- Harada T, Maronpot RR, Morris RW, Boorman GA. 1990. Effects of mononuclear cell leukemia on altered hepatocellular foci in Fischer 344 rats. Vet Pathol. 27:110–116.
- Hart DN, Baker BW, Inglis MJ, Nimmo JC, Starling GC, Deacon E, Rowe M, Beard ME. 1992. Epstein-Barr viral DNA in acute large granular lymphocyte (natural killer) leukemic cells. Blood. 79:2116–2123.
- Haseman JK. 1983. A reexamination of false-positive rates for carcinogenesis studies. Fundam Appl Toxicol. 3:334–339.
- Haseman JK, Hailey JR, Morris RW. 1998. Spontaneous neoplasm incidences in Fischer 344 rats and B6C3F1 mice in two-year carcinogenicity studies: a National Toxicology Program update. Toxicol Pathol. 26:428–441.
- Haseman JK, Huff JE, Rao GN, Arnold JE, Boorman GA, McConnell EE. 1985. Neoplasms observed in untreated and corn oil gavage control groups of F344/N rats and (C57BL/6N X C3H/HeN)F1 (B6C3F1) mice. J Natl Cancer Inst. 75:975–984.
- Haseman JK, Ney E, Nyska A, Rao GN. 2003. Effect of diet and animal care/housing protocols on body weight, survival, tumor incidences, and nephropathy severity of F344 rats in chronic studies. Toxicol Pathol. 31:674–681.
- Haseman JK, Rao GN. 1992. Effects of corn oil, time-related changes, and inter-laboratory variability on tumor occurrence in control Fischer 344 (F344/N) rats. Toxicol Pathol. 20:52–60.
- Haseman JK, Young E, Eustis SL, Hailey JR. 1997. Body weight-tumor incidence correlations in long-term rodent carcinogenicity studies. Toxicol Pathol. 25:256–263.
- Hassan R, Alexander R. 2005. Nonpleural mesotheliomas: mesothelioma of the peritoneum, tunica vaginalis, and pericardium. Hematol Oncol Clin North Am. 19:1067–1087, vi.
- Hasserjian RP, Harris NL. 2007. NK-cell lymphomas and leukemias: a spectrum of tumors with variable manifestations and immunophenotype. Am J Clin Pathol. 127:860–868.
- Hellman S, Moloney WC, Meissner WA. 1982. Paradoxical effect of radiation on tumor incidence in the rat: implications for radiation therapy. Cancer Res. 42:433–436.
- Hendry J, Fraser S, White J, Rajan P, Hendry DS. 2015. Retroperitoneal lymph node dissection (RPLND) for malignant phenotype Leydig cell tumours of the testis: a 10-year experience. Springerplus. 4:20.
- Hiraga K, Fujii T. 1985. Carcinogenicity testing of acetaminophen in F344 rats. Jpn J Cancer Res. 76:79–85.
- Hoei-Hansen CE, Holm M, Rajpert-De Meyts E, Skakkebaek NE. 2003. Histological evidence of testicular dysgenesis in contralateral biopsies from 218 patients with testicular germ cell cancer. J Pathol. 200:370–374.
- Huff JE. 1994. Chemicals causally associated with cancers in humans and laboratory animals. A perfect concordance. In: Waalkes MP, Ward JM, editors. Carcinogenesis. New York: Raven Press. p. 25–37.
- Hursting SD, Switzer BR, French JE, Kari FW. 1993. The growth hormone: insulin-like growth factor 1 axis is a mediator of diet restriction-induced inhibition of mononuclear cell leukemia in Fischer rats. Cancer Res. 53:2750–2757.
- Hursting SD, Switzer BR, French JE, Kari FW. 1994. Inhibition of rat mononuclear cell leukemia by corn oil gavage: in vivo, in situ and immune competence studies. Carcinogenesis. 15:193–199.
- Husain AN, Colby T, Ordonez N, Krausz T, Attanoos R, Beasley MB, Borczuk AC, Butnor K, Cagle PT, Chirieac LR, et al. 2013. Guidelines for pathologic diagnosis of malignant mesothelioma: 2012 update of the consensus statement from the International Mesothelioma Interest Group. Arch Pathol Lab Med. 137:647–667.
- Ikegami Y, Kawai N, Tozawa K, Hayashi Y, Kohri K. 2008. Malignant mesothelioma of the tunica vaginalis testis related to recent asbestos. Int J Urol. 15:560–561.
- Ilgren E. 1993. Mesotheliomas of animals: a comprehensive tabular compendium of the world's literature. Oxford: CRC Press.
- Ilgren EB, Wagner JC. 1991. Background incidence of mesothelioma: animal and human evidence. Regul Toxicol Pharmacol. 13:133–149.
- Imai K, Yoshimura K, Yamaguchi K, Matsui E, Isaka H, Hashimoto K, Boorman G. 1990. Effects of dietary restriction on age-associated pathologic changes in F-344 rats. J Toxicol Pathol. 3:209–221.
- Inbar S, Neeman E, Avraham R, Benish M, Rosenne E, Ben-Eliyahu S. 2011. Do stress responses promote leukemia progression? An animal study suggesting a role for epinephrine and prostaglandin-E2 through reduced NK activity. PLoS One. 6:e19246.
- Johansson P, Bergmann A, Rahmann S, Wohlers I, Scholtysik R, Przekopowitz M, Seifert M, Tschurtschenthaler G, Webersinke G, Jager U, et al. 2015. Recurrent alterations of TNFAIP3 (A20) in T-cell large granular lymphocytic leukemia. Int J Cancer. 138:121–124.
- Johnson KA, Gorzinski SJ, Bodner KM, Campbell RA, Wolf CH, Friedman MA, Mast RW. 1986. Chronic toxicity and oncogenicity study on acrylamide incorporated in the drinking water of Fischer 344 rats. Toxicol Appl Pharmacol. 85:154–168.
- Jones MA, Young RH, Scully RE. 1995. Malignant mesothelioma of the tunica vaginalis. A clinicopathologic analysis of 11 cases with review of the literature. Am J Surg Pathol. 19:815–825.
- Karpe B, Fredricsson B, Svensson J, Ritzen EM. 1982. Testosterone concentration within the tunica vaginalis of boys and adult men. Int J Androl. 5:549–556.
- Kato R, Matsuda Y, Maehana T, Miyao N, Konishi Y, Kon S. 2012. Intrascrotal malignant mesothelioma diagnosed after surgery for hydrocele testis: a case report. Hinyokika Kiyo. 58:45–48.
- Kato Y, Tsuta K, Seki K, Maeshima AM, Watanabe S, Suzuki K, Asamura H, Tsuchiya R, Matsuno Y. 2007. Immunohistochemical detection of GLUT-1 can discriminate between reactive mesothelium and malignant mesothelioma. Mod Pathol. 20:215–220.
- Kawa-Ha K, Ishihara S, Ninomiya T, Yumura-Yagi K, Hara J, Murayama F, Tawa A, Hirai K. 1989. CD3-negative lymphoproliferative disease of granular lymphocytes containing Epstein-Barr viral DNA. J Clin Invest. 84:51–55.
- Kim I, Young RH, Scully RE. 1985. Leydig cell tumors of the testis. A clinicopathological analysis of 40 cases and review of the literature. Am J Surg Pathol. 9:177–192.
- King-Herbert A, Thayer K. 2006. NTP workshop: animal models for the NTP rodent cancer bioassay: stocks and strains – should we switch? Toxicol Pathol. 34:802–805.
- King-Herbert AP, Sills RC, Bucher JR. 2010. Commentary: update on animal models for NTP studies. Toxicol Pathol. 38:180–181.
- Kivipensas P, Bjorkqvist AM, Karhu R, Pelin K, Linnainmaa K, Tammilehto L, Mattson K, Kallioniemi QP, Knuutila S. 1996. Gains and losses of DNA sequences in malignant mesothelioma by comparative genomic hybridization. Cancer Genet Cytogenet. 89:7–13.
- Kodell RL, Blackwell BN, Bucci TJ, Greenman DL. 1995. Cause-of-death assignment at the National Center for Toxicological Research. Toxicol Pathol. 23:241–247.
- Kusewitt D, Hahn F, Pickerell J. 1982. Hematologic and serum chemical characteristics of mononuclear leukemia in Fischer 344 rats. Lab Anim Sci. 32:275–277.
- Lafferty JS, Kamendulis LM, Kaster J, Jiang J, Klaunig JE. 2004. Subchronic acrylamide treatment induces a tissue-specific increase in DNA synthesis in the rat. Toxicol Lett. 154:95–103.
- Lagana SM, Taub RN, Borczuk AC. 2012. Utility of glucose transporter 1 in the distinction of benign and malignant thoracic and abdominal mesothelial lesions. Arch Pathol Lab Med. 136:804–809.
- Le Cornet C, Lortet-Tieulent J, Forman D, Béranger R, Flechon A, Fervers B, Schüz J, Bray F. 2014. Testicular cancer incidence to rise by 25% by 2025 in Europe? Model-based predictions in 40 countries using population-based registry data. Eur J Cancer. 50:831–839.
- Lee AF, Gown AM, Churg A. 2013. IMP3 and GLUT-1 immunohistochemistry for distinguishing benign from malignant mesothelial proliferations. Am J Surg Pathol. 37:421–426.
- Lee S, Illei PB, Han JS, Epstein JI. 2014. Florid mesothelial hyperplasia of the tunica vaginalis mimicking malignant mesothelioma: a clinicopathologic study of 12 cases. Am J Surg Pathol. 38:54–59.
- Libé R, Fratticci A, Lahlou N, Jornayvaz FR, Tissier F, Louiset E, Guibourdenche J, Vieillefond A, Zerbib M, Bertherat J. 2012. A rare cause of hypertestosteronemia in a 68-year-old patient: a Leydig cell tumor due to a somatic GNAS (guanine nucleotide-binding protein, alpha-stimulating activity polypeptide 1)-activating mutation. J Androl. 33:578–584.
- Lindsey J. 1979. Historical foundations. In: Baker H, Lindsey J, Weisbroth S, editors. The laboratory rat. Vol. 1. Biology and diseases. New York: Academic Press. p. 2–33.
- Lington AW, Bird MG, Plutnick RT, Stubblefield WA, Scala RA. 1997. Chronic toxicity and carcinogenic evaluation of diisononyl phthalate in rats. Fundam Appl Toxicol. 36:79–89.
- Lipman RD, Dallal GE, Bronson RT. 1999. Effects of genotype and diet on age-related lesions in ad libitum fed and calorie-restricted F344, BN, and BNF3F1 rats. J Gerontol A Biol Sci Med Sci. 54:B478–B491.
- Losco PE, Ward JM. 1984. The early stage of large granular lymphocyte leukemia in the F344 rat. Vet Pathol. 21:286–291.
- Maizlin ZV, Belenky A, Kunichezky M, Sandbank J, Strauss S. 2004. Leydig cell tumors of the testis: gray scale and color Doppler sonographic appearance. J Ultrasound Med. 23:959–964.
- Maqdasy S, Bogenmann L, Batisse-Lignier M, Roche B, Franck F, Desbiez F, Tauveron I. 2015. Leydig cell tumor in a patient with 49,XXXXY karyotype: a review of literature. Reprod Biol Endocrinol. 13:72.
- Marinaccio A, Binazzi A, Di Marzio D, Scarselli A, Verardo M, Mirabelli D, Gennaro V, Mensi C, Merler E, De Zotti R, et al. 2010. Incidence of extrapleural malignant mesothelioma and asbestos exposure, from the Italian national register. Occup Environ Med. 67:760–765.
- Maronpot RR, Flake G, Huff J. 2004. Relevance of animal carcinogenesis findings to human cancer predictions and prevention. Toxicol Pathol. 32:40–48.
- Maronpot RR, Thoolen RJ, Hansen B. 2015. Two-year carcinogenicity study of acrylamide in Wistar Han rats with in utero exposure. Exp Toxicol Pathol. 67:189–195.
- Maronpot RR, Zeiger E, McConnell EE, Kolenda-Roberts H, Wall H, Friedman MA. 2009. Induction of tunica vaginalis mesotheliomas in rats by xenobiotics. Crit Rev Toxicol. 39:512–537.
- Masoudi JF, Van Arsdalen K, Rovner ES. 1999. Organ-sparing surgery for bilateral leydig cell tumor of the testis. Urology. 54:744.
- Mensi C, Pellegatta M, Sieno C, Consonni D, Riboldi L, Bertazzi PA. 2012. Mesothelioma of tunica vaginalis testis and asbestos exposure. BJU Int. 110:533–537.
- Moloney WC, Boschetti AE, King V. 1969. Observations on leukemia in Wistar Furth rats. Cancer Res. 29:938–946.
- Moloney WC, Boschetti AE, King VP. 1970. Spontaneous leukemia in Fischer rats. Cancer Res. 30:41–43.
- Moloney WC, King VP. 1973. Reduction of leukemia incidence following splenectomy in the rat. Cancer Res. 33:573–574.
- Morris JE, Sasser LB, Miller DL, Dagle GE, Rafferty CN, Ebi KL, Anderson LE. 1999. Clinical progression of transplanted large granular lymphocytic leukemia in Fischer 344 rats exposed to 60 Hz magnetic fields. Bioelectromagnetics. 20:48–56.
- Nolte T, Rittinghausen S, Kellner R, Karbe E, Kittel B, Rinke M, Deschl U. 2011. RITA-Registry of Industrial Toxicology Animal data: the application of historical control data for Leydig cell tumors in rats. Exp Toxicol Pathol. 63:645–656.
- Nyska A, Hester SD, Cooper RL, Goldman JM, Stoker TE, House D, Wolf DC. 2002. Single or group housing altered hormonal physiology and affected pituitary and interstitial cell kinetics. J Toxicol Sci. 27:449–457.
- Nyska A, Leininger JR, Maronpot RR, Haseman JK, Hailey JR. 1998. Effect of individual versus group caging on the incidence of pituitary and Leydig cell tumors in F344 rats: proposed mechanism. Med Hypotheses. 50:525–529.
- Ohshima K, Suzumiya J, Kanda M, Kato A, Kikuchi M. 1998. Integrated and episomal forms of Epstein-Barr virus (EBV) in EBV associated disease. Cancer Lett. 122:43–50.
- Ozyavuz R, Ozen HA, Gedikoglu G, Ozgu IH, Sahin A, Tekgul S, Remzi D. 1993. Leydig cell tumour of the testis: presentation of two cases. Int Urol Nephrol. 25:385–388.
- Pacheco AJ, Torres JL, de la Guardia FV, Arrabal Polo MA, Gomez AZ. 2009. Intraparenchymatous adenomatoid tumor dependent on the rete testis: a case report and review of literature. Indian J Urol. 25:126–128.
- Peterson JT, Jr, Greenberg SD, Buffler PA. 1984. Non-asbestos-related malignant mesothelioma. A review. Cancer. 54:951–960.
- Plas E, Riedl CR, Pfluger H. 1998. Malignant mesothelioma of the tunica vaginalis testis: review of the literature and assessment of prognostic parameters. Cancer. 83:2437–2446.
- Poullot E, Zambello R, Leblanc F, Bareau B, De March E, Roussel M, Boulland ML, Houot R, Renault A, Fest T, et al. 2014. Chronic natural killer lymphoproliferative disorders: characteristics of an international cohort of 70 patients. Ann Oncol. 25:2030–2035.
- Prentice DE, Siegel RA, Donatsch P, Qureshi S, Ettlin RA. 1992. Mesulergine induced Leydig cell tumours, a syndrome involving the pituitary-testicular axis of the rat. Arch Toxicol Suppl. 15:197–204.
- Purdue MP, Devesa SS, Sigurdson AJ, McGlynn KA. 2005. International patterns and trends in testis cancer incidence. Int J Cancer. 115:822–827.
- Rajan V, Nandhakumar R, Shanmugasundaram S, Ravi R, Natarajan S, Mohan G, Nanjundappan PM. 2013. Paratesticular malignant mesothelioma – a rare case presentation. Indian J Surg. 75:174–176.
- Rall DP, Hogan MD, Huff JE, Schwetz BA, Tennant RW. 1987. Alternatives to using human experience in assessing health risks. Annu Rev Public Health. 8:355–385.
- Rao G, Boorman G. 1990. History of the F344 rat. In: Boorman G, Eustis S, Elwell M, Montgomery C, Jr, MacKenzie W, editors. Pathology of the Fischer rat. Reference and Atlas. New York: Academic Press. p. 5–8.
- Rao GN, Haseman JK. 1993. Influence of corn oil and diet on body weight, survival, and tumor incidences in F344/N rats. Nutr Cancer. 19:21–30.
- Rao GN, Haseman JK, Grumbein S, Crawford DD, Eustis SL. 1990. Growth, body weight, survival, and tumor trends in F344/N rats during an eleven-year period. Toxicol Pathol. 18:61–70.
- Reynolds CW. 1985. Large granular lymphocyte (LGL) lymphoproliferative diseases: naturally cytotoxic tumors in man and experimental animals. Crit Rev Oncol Hematol. 2:185–208.
- Reynolds CW, Bere EW, Jr, Ward JM. 1984. Natural killer activity in the rat. III. Characterization of transplantable large granular lymphocyte (LGL) leukemias in the F344 rat. J Immunol. 132:534–540.
- Reynolds CW, Bonyhadi M, Herberman RB, Young HA, Hedrick SM. 1985. Lack of gene rearrangement and mRNA expression of the beta chain of the T cell receptor in spontaneous rat large granular lymphocyte leukemia lines. J Exp Med. 161:1249–1254.
- Ruskova A, Thula R, Chan G. 2004. Aggressive natural killer-cell leukemia: report of five cases and review of the literature. Leuk Lymphoma. 45:2427–2439.
- Ryder J, Wang X, Bao L, Gross SA, Hua F, Irons RD. 2007. Aggressive natural killer cell leukemia: report of a Chinese series and review of the literature. Int J Hematol. 85:18–25.
- Sacksteder MR. 1976. Occurrence of spontaneous tumors in the germfree F344 rat. J Natl Cancer Inst. 57:1371–1373.
- Sass B, Rabstein LS, Madison R, Nims RM, Peters RL, Kelloff GJ. 1975. Incidence of spontaneous neoplasms in F344 rats throughout the natural life-span. J Natl Cancer Inst. 54:1449–1456.
- Sasser LB, Morris JE, Miller DL, Rafferty CN, Ebi KL, Anderson LE. 1996. Exposure to 60 Hz magnetic fields does not alter clinical progression of LGL leukemia in Fischer rats. Carcinogenesis. 17:2681–2687.
- Schurkes C, Brock W, Abel J, Unfried K. 2004. Induction of 8-hydroxydeoxyguanosine by man-made vitreous fibres and crocidolite asbestos administered intraperitoneally in rats. Mutat Res. 553:59–65.
- Segura-Gonzalez M, Urias-Rocha J, Castelan-Pedraza J. 2015. Malignant mesothelioma of the tunica vaginalis: a rare neoplasm-case report and literature review. Clin Genitourin Cancer. 13:e401–e405.
- Semenzato G, Marino F, Zambello R. 2012. State of the art in natural killer cell malignancies. Int J Lab Hematol. 34:117–128.
- Sharpe RM, McNeilly AS. 1979. The effect of induced hyperprolactinaemia on Leydig cell function and LH-induced loss of LH-receptors in the rat testis. Mol Cell Endocrinol. 16:19–27.
- Shiga A, Narama I. 2015. Hepatic lesions caused by large granular lymphocyte leukemia in Fischer 344 rats: similar morphologic features and morphogenesis to those of nodular regenerative hyperplasia (NRH) in the human liver. Toxicol Pathol. 43:852–864.
- Shimokawa I, Yu BP, Higami Y, Ikeda T, Masoro EJ. 1993. Dietary restriction retards onset but not progression of leukemia in male F344 rats. J Gerontol. 48:B68–B73.
- Shipp A, Lawrence G, Gentry R, McDonald T, Bartow H, Bounds J, Macdonald N, Clewell H, Allen B, Van Landingham C. 2006. Acrylamide: review of toxicity data and dose-response analyses for cancer and noncancer effects. Crit Rev Toxicol. 36:481–608.
- Skakkebaek NE, Holm M, Hoei-Hansen C, Jorgensen N, Rajpert-De Meyts E. 2003. Association between testicular dysgenesis syndrome (TDS) and testicular neoplasia: evidence from 20 adult patients with signs of maldevelopment of the testis. APMIS. 111:1–9. discussion 9–11.
- Sokol L, Loughran TP, Jr. 2006. Large granular lymphocyte leukemia. Oncologist. 11:263–273.
- Song SY, Kim WS, Ko YH, Kim K, Lee MH, Park K. 2002. Aggressive natural killer cell leukemia: clinical features and treatment outcome. Haematologica. 87:1343–1345.
- Stefanski S, Elwell M, Stromberg P. 1990. Spleen, lymph nodes, and thymus. In: Boorman G, Eustis S, Elwell M, Montgomery C, Jr, MacKenzie W, editors. Pathology of the Fischer rat. Reference and Atlas.New York: Academic Press. p. 369–394.
- Stefanski SA, Greenwell A, Merrick BA, Brown TT, Reynolds SH. 1995. Proliferating cell nuclear antigen staining of Fischer-344/N rat spleens affected by large granular lymphocyte leukemia. Toxicol Pathol. 23:1–6.
- Steinbach T, Maronpot R, Hardisty J. 2015. Human relevance of rodent Leydig cell tumors. In: Harbison R, Bourgeois M, Johnson G, editors. Hamilton & Hardy's industrial toxicology. New York: John Wiley & Sons. p. 1189–1196.
- Steinway SN, LeBlanc F, Loughran TP. 2014. The pathogenesis and treatment of large granular lymphocyte leukemia. Blood Rev. 28:87–94.
- Stromberg PC, Grants IS, Kociba GJ, Krakowka GS, Mezza LE. 1990. Serial syngeneic transplantation of large granular lymphocyte leukemia in F344 rats. Vet Pathol. 27:404–410.
- Stromberg PC, Rojko JL, Vogtsberger LM, Cheney C, Berman R. 1983d. Immunologic, biochemical, and ultrastructural characterization of the leukemia cell in F344 rats. J Natl Cancer Inst. 71:173–181.
- Stromberg PC, Vogtsberger LM. 1983a. Pathology of the mononuclear cell leukemia of Fischer rats. I. Morphologic studies. Vet Pathol. 20:698–708.
- Stromberg PC, Vogtsberger LM, Marsh LR. 1983c. Pathology of the mononuclear cell leukemia of Fischer rats. III. Clinical chemistry. Vet Pathol. 20:718–726.
- Stromberg PC, Vogtsberger LM, Marsh LR, Wilson FD. 1983b. Pathology of the mononuclear cell leukemia of Fischer rats. II. Hematology. Vet Pathol. 20:709–717.
- Stromberg PC, Vogtsberger LM, McMurray DN, Marsh LR, Kotur MS, Brown CA. 1985. Behavior of transplanted large granular lymphocyte leukemia in Fischer 344 rats. Lab Invest. 53:200–208.
- Suzuki R, Suzumiya J, Nakamura S, Aoki S, Notoya A, Ozaki S, Gondo H, Hino N, Mori H, Sugimori H, et al. 2004. Aggressive natural killer-cell leukemia revisited: large granular lymphocyte leukemia of cytotoxic NK cells. Leukemia. 18:763–770.
- Taheri ZM, Mehrafza M, Mohammadi F, Khoddami M, Bahadori M, Masjedi MR. 2008. The diagnostic value of Ki-67 and repp86 in distinguishing between benign and malignant mesothelial proliferations. Arch Pathol Lab Med. 132:694–697.
- Takeda M, Kasai T, Enomoto Y, Takano M, Morita K, Kadota E, Iizuka N, Maruyama H, Nonomura A. 2012. Genomic gains and losses in malignant mesothelioma demonstrated by FISH analysis of paraffin-embedded tissues. J Clin Pathol. 65:77–82.
- Tanigawa H, Onodera H, Maekawa A. 1987. Spontaneous mesotheliomas in Fischer rats – a histological and electron microscopic study. Toxicol Pathol. 15:157–163.
- Testa JR, Cheung M, Pei J, Below JE, Tan Y, Sementino E, Cox NJ, Dogan AU, Pass HI, Trusa S, et al. 2011. Germline BAP1 mutations predispose to malignant mesothelioma. Nat Genet. 43:1022–1025.
- Thomas J, Haseman JK, Goodman JI, Ward JM, Loughran TP, Jr, Spencer PJ. 2007. A review of large granular lymphocytic leukemia in Fischer 344 rats as an initial step toward evaluating the implication of the endpoint to human cancer risk assessment. Toxicol Sci. 99:3–19.
- Thurman JD, Bucci TJ, Hart RW, Turturro A. 1994. Survival, body weight, and spontaneous neoplasms in ad libitum-fed and food-restricted Fischer-344 rats. Toxicol Pathol. 22:1–9.
- Tomatis L, Aitio A, Wilbourn J, Shuker L. 1989. Human carcinogens so far identified. Jpn J Cancer Res. 80:795–807.
- Trabert B, Zugna D, Richiardi L, McGlynn KA, Akre O. 2013. Congenital malformations and testicular germ cell tumors. Int J Cancer. 133:1900–1904.
- Triola MF. 2010. Elementary statistics. 11th ed. Boston: Pearson Education.
- Tsitouridis I, Maskalidis C, Panagiotidou D, Kariki EP. 2014. Eleven patients with testicular leydig cell tumors: clinical, imaging, and pathologic correlation. J Ultrasound Med. 33:1855–1864.
- Turek FW, Desjardins C. 1979. Development of Leydig cell tumors and onset of changes in the reproductive and endocrine systems of aging F344 rats. J Natl Cancer Inst. 63:969–975.
- Turturro A, Blank K, Murasko D, Hart R. 1994. Mechanisms of caloric restriction affecting aging and disease. Ann N Y Acad Sci. 719:159–170.
- Ulbright TM, Srigley JR, Hatzianastassiou DK, Young RH. 2002. Leydig cell tumors of the testis with unusual features: adipose differentiation, calcification with ossification, and spindle-shaped tumor cells. Am J Surg Pathol. 26:1424–1433.
- Unluer E, Ozcan D, Altin S. 1990. Malignant Leydig cell tumour of the testis: a case report and review of the literature. Int Urol Nephrol. 22:455–460.
- Versnel MA, Hagemeijer A, Bouts MJ, van der Kwast TH, Hoogsteden HC. 1988. Expression of c-sis (PDGF B-chain) and PDGF A-chain genes in ten human malignant mesothelioma cell lines derived from primary and metastatic tumors. Oncogene. 2:601–605.
- Vukina J, Chism DD, Sharpless JL, Raynor MC, Milowsky MI, Funkhouser WK. 2015. Metachronous bilateral testicular Leydig-like tumors leading to the diagnosis of congenital adrenal hyperplasia (adrenogenital syndrome). Case Rep Pathol. 2015:459318.
- Ward J, Rehm S, Reynolds C. 1990. Tumours of the hematopoietic system. In: Turusov V, Mohr U, editors. Pathology of tumours in laboratory animals. Vol. 1. Tumours of the rat. Lyon: International Agency for Research on Cancer. p. 625–657.
- Ward JM, Reynolds CW. 1983. Large granular lymphocyte leukemia. A heterogeneous lymphocytic leukemia in F344 rats. Am J Pathol. 111:1–10.
- Weisburger EK. 1983. History of the bioassay program of the National Cancer Institute. Prog Exp Tumor Res. 26:187–201.
- Weisburger JH, Rivenson A, Reinhardt J, Braley J, Pittman B, Zang E. 2002. On the occurrence of Leydig cell tumors in the F344 rat. Cancer Lett. 182:213–216.
- Winstanley AM, Landon G, Berney D, Minhas S, Fisher C, Parkinson MC. 2006. The immunohistochemical profile of malignant mesotheliomas of the tunica vaginalis: a study of 20 cases. Am J Surg Pathol. 30:1–6.
- Woodward PJ, Schwab CM, Sesterhenn IA. 2003. From the archives of the AFIP: extratesticular scrotal masses: radiologic-pathologic correlation. Radiographics. 23:215–240.
- Woodward PJ, Sohaey R, O'Donoghue MJ, Green DE. 2002. From the archives of the AFIP: tumors and tumorlike lesions of the testis: radiologic-pathologic correlation. Radiographics. 22:189–216.
- Xiao SY, Rizzo P, Carbone M. 2000. Benign papillary mesothelioma of the tunica vaginalis testis. Arch Pathol Lab Med. 124:143–147.
- Yen CH, Lee CT, Su CJ, Lo HC. 2012. Malignant mesothelioma of the tunica vaginalis testis: a malignancy associated with recurrent epididymitis? World J Surg Oncol. 10:238.
- Zhang Q, Jing W, Ouyang J, Zeng H, George SK, Liu Z. 2014. Six cases of aggressive natural killer-cell leukemia in a Chinese population. Int J Clin Exp Pathol. 7:3423–3431.