
Abstract
There is a need for novel nanomaterials with properties not yet exploited in regenerative nanomedicine. Based on lessons learned from the oldest metazoan phylum, sponges, it has been recognized that two previously ignored or insufficiently recognized principles play an essential role in tissue regeneration, including biomineral formation/repair and wound healing. Firstly, the dependence on enzymes as a driving force and secondly, the availability of metabolic energy. The discovery of enzymatic synthesis and regenerative activity of amorphous biosilica that builds the mineral skeleton of siliceous sponges formed the basis for the development of successful strategies for the treatment of osteochondral impairments in humans. In addition, the elucidation of the functional significance of a second regeneratively active inorganic material, namely inorganic polyphosphate (polyP) and its amorphous nanoparticles, present from sponges to humans, has pushed forward the development of innovative materials for both soft (skin, cartilage) and hard tissue (bone) repair. This energy-rich molecule exhibits a property not shown by any other biopolymer: the delivery of metabolic energy, even extracellularly, necessary for the ATP-dependent tissue regeneration. This review summarizes the latest developments in nanobiomaterials based on these two evolutionarily old, regeneratively active materials, amorphous silica and amorphous polyP, highlighting their specific, partly unique properties and mode of action, and discussing their possible applications in human therapy. The results of initial proof-of-concept studies on patients demonstrating complete healing of chronic wounds are outlined.
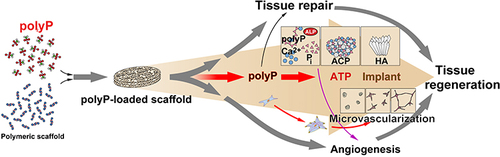
Graphical Abstract
Introduction
Over the course of its life, the integrity of any metazoan, from the simplest organisms to humans, is compromised to varying degrees by injury or chronic degenerative changes, leading to dysfunction. In humans, when the physiological repair capacity is insufficient, medical intervention is required to prevent a collapse of the fragile cellular homeostasis and the tuned organization of cells and tissues. There is a need for new concepts in regenerative therapy. Animals that have physiologically extensive regenerative abilities, such as sponges, are possible models for copying these concepts from nature. During the evolution of the metazoans from their basal phylum, the sponges (Porifera), with their ~40,000 predicted protein-coding loci,Citation1 to the “crown taxa”, humans, with 20,000 to 25,000 genes,Citation2 a gradual decrease in the genetic complexity is observed. This fact supports the view that the basal animals like sponges have a higher redundancy potential, combined with a variety of alternative metabolic circuits that provide them with a higher evolutionary stability. In turn, the regeneration capability of basal animals is more extensive compared to the “more advanced” taxa, including humans.
In animals, particularly in humans, some defects are not adequately repaired especially when physiological functional restoration is not supported by the inherent regeneration potential. If tissue/cell homeostasis is not maintained, the repair processes are insufficient with the result that the affected, damaged tissue is only replaced by tissue-like entities that lack physiological and biochemical functions. Or, as in present-day solutions, the defective parts are replaced or augmented by largely inert or only partially biological substituting materials. For example, the deficiencies of the bone-supporting implants can be overcome with tissue regenerating materials that restore not only the mechanical but also the biological and functional properties of the damaged tissue in the defect region. It is important to emphasize that any regeneration process requires an optimal physiological functioning of the cells, which depends on molecular and mechanical signals from the environment.Citation3 Both the cells in the tissue surrounding the defect, and particularly the cells within the areas to be repaired, must be supplied with growth factors and nutrients that support the anabolic repair pathways and, importantly, with metabolic energy (ATP or another energy-carrying molecule).Citation4 Not only intracellularly but also extracellularly, a sufficient supply of biochemically useful energy is important for the regeneration of the extracellular matrix (ECM). The number of cells in these regions is often low, for example in cartilage, which contains only ~10% chondrocytes.Citation5 During regeneration, biochemical reactions in the ECM are driven by the cell-based synthesis of fibrous mats (collagen) or hydrogels (hydrogel-forming polypeptides). These reactions are anabolic reactions that require the conversion of chemical fuels like glucose into ATP (energy). In addition, enzymes are needed that break down larger precursor molecules in order to drive anabolic ones in bone, cartilage, and muscle regeneration.Citation6 Accordingly, the two cornerstones, enzymes and energy, contribute significantly to the regeneration pathways. The initial basis for this gain came from studies from the basal animals, the sponges.
This review has a special focus on the regenerative abilities of two inorganic biomaterials in the formation and repair of both soft and hard tissues, amorphous silica (“biosilica”) and amorphous polyphosphate (polyP). Both biomaterials or biopolymers are regeneratively active only in their amorphous form and, in accordance with their physiological function, exhibit properties that the conventional organic biopolymers used in regenerative medicine do not have, such as in the case of polyP, the delivery of metabolic energy required for energy-dependent repair processes, as discussed in this review. In particular, the role that sponges, as the most basal phylum, have played in understanding the critical function of enzymes in the energy-dependent tissue regeneration/repair processes, including the formation of biomineralized structures, is discussed, while previous reviews on these biomaterials primarily focus on either only one these materialsCitation7,Citation8 or on their physicochemical properties or more mechanistic aspects.Citation9
Regeneration Capacity Across Animal Taxa
The basic principles of regeneration processes have been enigmatic for a long time. In 1606, it was stressed that bone formation and the regeneration of this organ depend on spiritual forces, virtus naturalis, based and localized in the lower body area, which is characterized by a “warm and humid” local environment.Citation10 Later, especially by Trembley, first evidence-based experiments were performed, which revealed remarkable regenerative abilities especially in the more basal Metazoa such as the phylum Cnidaria.Citation11 The sponges (Porifera), the basal phylum of Metazoa, traditionally served as a model system for regeneration.Citation12 Animal taxa that evolved later from sponges have a reduced regeneration capacity mainly due to the presence of lower amounts of stem cells.Citation13 Thomas Hunt Morgan introduced the perspective of functional genomics by studying the regeneration of segments in earthworms as a model.Citation14
In , a summary of the regeneration capacity across various animal taxa is shown. The ability to regenerate damaged or lost tissue greatly varies among different species and can include the whole body, the primary body axis, or only certain structures or tissues.Citation15 Whole-body regeneration is particularly found in the basal metazoans such as sponges, cnidarians and ctenophores.Citation16,Citation17 The regeneration of the sponge body or sponge tissue after an injury can either start from small body fragments or occur via aggregation of dissociated sponge cells.Citation16 Sponges as sessile marine organisms are particularly prone to wounding, eg, by grazers or mechanical injury, which may lead to the occurrence of chronic wounds if repair fails and homeostasis is not retained. Investigations of the response of the sponge Aplysina aerophoba to damage caused mechanically or by a spongivorous opisthobranch using differential gene expression analysis based on RNA-sequence data revealed a repair mechanism involving enzymes/proteins such as metalloproteases, transglutaminases, and integrins, and signaling pathways (Wnt and mitogen-activated protein kinase – MAPK), similar to that involved in wound healing in higher animals, including humans.Citation18
Table 1 Regeneration Capacity Across Various Animal Taxa
Taking Concepts from Nature as a Blueprint for Regeneration
Intense efforts over the past 40 years have uncovered, in a scientific, causal-analytical manner, the strategies of how living systems function with the aim of exploiting them to meet the urgent needs in engineering tissue replacement biomaterials. Biomimetic and bioinspired approaches based on these achievements have contributed considerably to a paradigm change in the field of tissue engineering.Citation20–22 Studies on sponges, the evolutionarily oldest metazoans, showed that these animals produce a range of organic biomolecules that they effectively defend against bacteria and viruses.Citation23–25 Then, the application of molecular biology techniques made it possible to show – what was not expected before – that these basal animals have in their protein toolkit, eg, building blocks even for basic immune defense systems that are successfully used by humans against foreign invaders. The existence of the Rhesus factorCitation26 or the polymorphic immunoglobulin molecules,Citation27 members of the acquired immune system, are mentioned here as examples. Even more, analyses of the formation of the skeletal system of the siliceous sponges, made of amorphous silica (“biosilica”), enabled the discovery of principles underlying tissue regeneration including biomineral formation, which were later found also in higher vertebrates including humans. In particular, it was discovered that biomineral formation is driven by enzymes, just like the biological formation of organic biomolecules and the functioning of metabolic pathways.Citation28,Citation29 Thus, sponges served as a model to progress advances in osteochondral regeneration.Citation30 In addition, due to their excellent mechanical but also light transmitting properties,Citation31 which are particularly evident in the siliceous sponge spicules, sponges can be considered and used as blueprints for the design of functional composite materials not only for nanobiomedical but also for technological applications.
In general, scaffolds formed from natural biopolymers provide an excellent matrix and niche for cell attachment, growth, migration, and differentiation, mimicking the ECM of native tissues (for a recent review, see Ref.Citation32). In these properties, natural biomaterials differ from bioinert synthetic materials used in tissue engineering and regenerative medicine that do not show bioactivity. Only by modifying them according to the design of natural polymers can synthetic bioinspired materials with biological activities be obtained.Citation33 Natural biomimetic biomaterials also have the advantage that they are non-toxic and do not exhibit genotoxic or teratogenic effects. In contrast to the natural biomaterials currently used, which are organic biopolymers such as polysaccharides (eg, hyaluronic acid, alginate, chitosan, and cellulose), polyesters (eg, polylactic acid and polyhydroxyalkanoates), and polypeptides/proteins (eg, collagen, gelatin, and fibroin),Citation32 the two inorganic materials described here, biosilica and polyP, although occurring physiologically, are also readily available by chemical methods, thereby overcoming the problem of batch variability shown by many natural polymers.
Sponges (Porifera): Their Seminal Contributions Towards Understanding Biomineralization
Based on the lessons learned from sponges, the two inorganic polymeric materials amorphous biogenic biosilica (“biosilica”) and inorganic polyphosphate (polyP) have attracted increasing interest in regenerative medicine due to their unique properties. While biosilica forms a sponge skeleton, polyP is a ubiquitous polymer, found from bacteria to men, in sponges particularly in the environment of biosilica. Findings in sponges have been of crucial importance for an understanding of the biological mineralization processes taking place in organisms, as compared to abiotic mineralization.Citation34,Citation35 While abiotic mineralization solely relies on chemical processes, biomineralization is based on chemical reactions that take advantage of genuine biochemical conversion processes. Biochemical reactions are principally catalyzed by enzymes, following the rules of thermodynamics. They often link individual processes together, where endergonic, non-spontaneous processes can be driven by exergonic, energy-releasing reactions.
More specifically, two principles have been revealed in sponges as are outlined in this review: First, that the vast majority of biochemical reactions, including biomineralization reactions, are enzyme driven and dependent. From sponges, the first enzyme that mainly contributes to inorganic mineral formation was discovered: silicatein.Citation28,Citation36,Citation37 While chemical reactions only strive to reach an equilibrium, biological processes proceed in open systems and are attuned to non-equilibria allowing both continuous and discontinuous flux of matter and energy.Citation38
Biosilica, enzymatically synthesized in vivo in sponges (), has become a paragon for a mineralic biomaterial. Later, it became increasingly aware that enzymes are also involved in the synthesis and degradation of the inorganic scaffold of other skeletal systems such as bone.Citation39 Of note here is the alkaline phosphatase (ALP), a ubiquitous, membrane-bound tetrameric enzyme that is attached via glycosyl-phosphatidylinositol moieties to the outer cell surface and involved in osteoid formation and mineralization.Citation40 The tartrate-resistant acid phosphatase and cathepsin K, an osteoclast-specific enzyme, are involved in bone resorption.Citation41 Furthermore, it has been suggested that bone formation also involves a carbonic anhydrase that synthesizes amorphous Ca-carbonate bioseeds during the course of Ca-phosphate bone mineral deposition.Citation42
Figure 1 Sponges, the earliest animals on Earth, have proven to be a valuable model system for understanding the basic principles of biomineralization. In these organisms, in siliceous sponges, the first enzyme involved in mineral formation (formation of the sponge skeletal elements, the spicules) was discovered: silicatein. Centers for ATP consumption were found adjacent to the spicules. Subsequently, the polymer inorganic polyphosphate (polyP) was identified as an extracellular storage for metabolic energy, which serves to generate ATP by successive enzymatic degradation via alkaline phosphatase (ALP) and adenylate kinase (ADK). In contrast, mineralization processes require higher temperatures for the chemical reactions instead of enzymes and ATP. The process of biomineralization is facilitated by organic templates/sheets, often collagen, secreted from the cells and undergoing modifications (often glycosylation), allowing the formation of supramolecular assemblies, and finally biomineral deposition.

In contrast to enzymatic, biotic biomineralization, abiotic mineralization, eg, geothermal mineralization, is driven by heat derived from, eg, cosmic rays/ionizing radiation (). In addition, it was shown that, in addition to enzymes, bone biomineralization as well as the organization of the bone architecture also require energy in the form of ATP,Citation43,Citation44 without specifying the extent.Citation45,Citation46 In fact, the energy issue was largely ignored until approaches to regenerate or repair tissue through biochemically based substitution therapy were addressed. Both osteoblasts and osteoclasts release ATP into the extracellular space in amounts that depend on the proliferation and differentiation state of the cells.Citation43 Despite a variety of ATP export channels into the extracellular space, the ATP concentration there is very low at ~10 nM, in contrast to the large intracellular pool at ~100 µM (see Ref.Citation5). Since the level of ATP in human blood is also low at ~100 nM, an additional energy source for ATP generation had to be postulated.
Based on the therapeutic success with polyP, after application of this polymer for the repair in different organs, bones, and chronic wounds,Citation30,Citation47–49 it was proposed and then proved that polyP with its energy-rich acid anhydride linkages could serve as a source for metabolically useful energy in the form of ATP and/or ADP,Citation50,Citation51 reviewed in Ref.Citation5 As reported later, the energy stored in polyP can be converted to ADP/ATP in a stepwise enzymatic reaction chain.
In addition to the two corner stones, (i) enzymes and (ii) metabolic energy, organic templates are usually required for the deposition of minerals during biomineralization. In bone, there are the collagen matrices built from collagen bundles that act as a platform for the apposition of mineral aggregates. Their post-translationally modified side chains, processed by hydroxylation and glycosylation, provide suitable deposition sites.Citation52
Sponge Biosilica
The sponges (Porifera) share a common body plan with the other, more evolved metazoans.Citation53 This fundamental finding, based on extensive studies of the (expressed) sponge genome, together with the discovery that these animals synthesize their mineral skeletons with the help of enzymes, as mentioned above,Citation28,Citation36,Citation37 formed the reason why these animals have become metazoic model taxa for the development of biomimetic materials and processes applicable for both nanobiomedical and engineering purposes for the benefit of humans. Equally important was the discovery that biomineral formation is an energy-consuming process that requires an energy (ATP) source.
Based on their skeletons, the Porifera are subdivided into the classes of the siliceous Demospongiae and Hexactinellida and the calcareous sponges, the class Calcarea.Citation54 The siliceous spicules of most Demospongiae remain individualized and the secreted cellular tissue units are formed around them. In these sponges, the spicules are linked together with organic molecules (collagen-related spongin and lectin) that form a bulky extracellular matrix.Citation55 It is the siliceous scaffold that directs the particular, species-characteristic form of the sponge tissue. The siliceous skeleton of the hexactinellids is composed of discrete spicules, which often fuse to a basket-like scaffold as in the hexactinellid Euplectella aspergillum ().Citation56 In particular, the siliceous skeleton of this class of sponges transmits light along the outer surface of the cage (), but also within the inner fan pocket (). Particularly impressive are the central giant spicules of the deep-sea sponge Monorhaphis chuni with a length of up to 3 m (diameter 1.7 cm), which represent the largest biosilica structures on Earth ().Citation57 Light can pass efficiently along these giant spicules (). These spicules are composed of up to 800 lamellae, each 5–10 μm thick, which are arranged concentrically around the axial canal (). In the center, the axial canal harbors a 10 µm thick proteinaceous filament, the axial filament ().
Figure 2 The siliceous sponges and their silica skeletons. (A–C) The hexactinellid sponge E. aspergillum; in (B), the silica cage is illuminated with a laser beam. (D and E) The hexactinellid Monorhaphis chuni with its up to 3 m large giant basal spicule; in (E), the silica rod is illuminated with green laser light. (F) Cross break through a giant basal spicule showing the central axial canal (ac) and the surrounding lamellae (lam). (G and H) A tylostyle spicule harboring (G) the central axial filament (af) of (H) the demosponge S. domuncula. (I) A broken aster spicule from the demosponge G. cydonium exposing the axial filament (af) in the axial canal (ac). (J) Cross break of a tissue unit from S. domuncula showing the immunostained silica-forming enzyme, silicatein, in the axial filament (af) and also on the surface (su) of the tylostyle. (K) Cryosection through a S. domuncula tissue unit; the transition in color from blue to orange reflects the increase in ATP level around the spicules (sp).

Enzymes Involved in Sponge Biomineralization
It is a distinctive feature of both classes of siliceous sponges, the demosponges and the hexactinellids, that their skeletal elements, composed of amorphous biosilica, are synthesized by an enzyme, silicatein. With silicatein, the first enzyme was discovered that catalyzes the formation of an inorganic “polymeric” material, here biosilica, from an inorganic precursor, here monomeric silicic acid.Citation37,Citation58–60
Silicatein is an enzyme protein found exclusively in Porifera. Three isoforms of silicatein are found in the axial filaments.Citation28,Citation37 Molecular sequence analyses revealed that the silicatein family of proteins originates from the cathepsin family of proteases, more precisely from cathepsin L.Citation61 Like the silicateins, the cathepsins are hydrolytic enzymes.Citation62 Since sponges are suspension/filter feeders, our group had proposed and then identified this major catabolic enzyme, cathepsin L.
Among the siliceous sponges, the demosponge Suberites domuncula () has been used in most cell biological and molecular studies, because this sponge species can be readily kept in aquaria and allowed the establishment of a cell culture, the primmorphs.Citation63 In the demosponges, the axial filament is voluminous ( and ). The silicatein filaments determine the morphology of the spicules, either rod-shaped (as in S. domuncula) or star-shaped (as in Geodia cydonium).
Cross fractures through the spicules revealed that not only the central silicatein rod but also the surface of the spicules reacts with antibodies against silicatein (). Interestingly, when cryosections through S. domuncula are studied, a regional distribution of ATP is measured (). The highest ATP levels are found in the vicinity of the spicules. The latter finding reflects the crucial role of ATP in the biomineralization processes and the dependence of the formation of skeletal elements on metabolic activity.Citation64 Biomineralization requires not only an enzyme that lowers the activation energy of this process but also metabolic energy in the form of ATP.
The other class of sponges, the calcareous sponges, Calcarea, are stabilized by calcareous spicules that are formed of calcite.Citation65 Following the general rule that any physiological crystalline structure is built from amorphous precursors, calcite formation starts with amorphous Ca-carbonate (ACC).Citation66 Our group found that the Ca-carbonate-based spicules are synthesized enzymatically by a carbonic anhydrase,Citation29 a finding that has been suggested earlier.Citation67 Also of interest is the finding that the ACC precursor is stabilized by polyP.Citation68 Consequently, incubation of ACC with the polyP-hydrolyzing enzyme ALP causes the transformation of the ACC phase into calcite.Citation69
The Substrate and Mechanism of Silicatein Reaction
Sponges take up silicon from seawater after conversion of SiO2 to soluble, biological biosilica (SiO2•nH2O) and from there to readily soluble H4SiO4 (aq).Citation70 Silicic acid is transported via a Si transporter into the sponge cells,Citation71 where it serves as a substrate for silicatein for the formation of amorphous biosilica. The synthetic silicon compound tetraethyl orthosilicate (TEOS), as a silicic acid precursor, was used for the functional analyses.Citation28,Citation58,Citation72
Silicatein catalyzes the (poly)condensation/polymerization of its physiological substrate, ortho-silicic acid, at low concentrations (<1 mM).Citation59,Citation73 Higher concentrations are required for chemical, non-enzymatic condensation reactions (>1 mM).Citation73–75 In bioinspired fabrication processes, silicatein acts as a hydrolase (silicic acid esterase) that facilitates the hydrolysis of alkoxysilane compounds (cleavage of Si–O–C “ester” bonds), eg, of TEOS.Citation28,Citation37,Citation76
Among demosponges, the first deduced amino acid sequences of silicateins were published for S. domunculaCitation28,Citation77 and T. aurantium.Citation37 The catalytic triad (catalytic center) of silicatein consists of the amino acids Ser, His, and Asn. The cathepsins have a Cys residue instead of Ser. Another characteristic sequence of silicatein is a Ser-rich cluster.
The mechanism of biosilica formation with silicatein has been outlined.Citation78 In the marine environment, the concentration of silicon is around ~10 mg mL‒1. Silicic acid is taken up by cells via a specific transporter where it acts as a substrate for silicatein ().Citation71 The reaction starts with nucleophilic attack of the catalytic triad Ser-OH group at a silicic acid substrate molecule, supported by hydrogen bridge formation to the His imidazole group in the catalytic center (). The third amino acid of the catalytic triad of silicatein, the Asn residue (not shown), binds to the OH leaving group of the substrate, which is released as a water molecule. This step enables the formed Ser-bound silicic acid, again facilitated by H-bridge formation to His, to undergo two condensation reactions, resulting in the formation of an enzyme-bound disilicic acid and then a trisilicic acid species, which is then released from the enzyme after cyclization as cyclotrisilicic acid (cyclotrisiloxane). Next, a purely chemical condensation process takes place, initiated by preferential addition of further silicic acid species to this cyclic silicon compound. The subsequent steps of silica formation basically follow the Stöber synthesis, allowing a controlled growth of spherical silica particles to condensed silica deposits by hydrolysis of alkyl silicates with subsequent polycondensation of the produced silicic acid units.Citation79
Figure 3 Physiological silicic acid polymerase reaction and bioinspired hydrolase reaction catalyzed by the biosilica-forming enzyme silicatein. Silicatein mediates the condensation of monomeric ortho-silicic acid [Si(OH)4] to polymeric amorphous silica (n•SiO2; formation of Si–O–Si bonds). In a second further reaction, silicatein acts as a hydrolase, which catalyzes the hydrolytic cleavage of the alkoxysilane compound TEOS [Si(OR)4; R = ethyl] (cleavage of Si–O–C bonds) allowing a bioinspired synthesis of artificial amorphous silica-based (nano)materials for biomedical applications. Shown in the center (enclosed in a ring) are the two amino acids of the catalytic triad, Ser and His, which interact with silicic acid and mediate the condensation. (A) Silicatein core protein of an aster from G. cydonium. (B) Complete sponge aster after incubation of silicatein with TEOS. (C) A spicule from S. domuncula with the axial canal (ac) centered by the silicatein-containing axial filament (af). (D) An axial filament with thorny silica (sil) protrusions. (E) Silicatein deposits layered onto thorny template, after reaction with TEOS. (F) Microspheres lacking silicatein after incubation with TEOS. (G) Silicatein-containing microspheres after TEOS incubation.
![Figure 3 Physiological silicic acid polymerase reaction and bioinspired hydrolase reaction catalyzed by the biosilica-forming enzyme silicatein. Silicatein mediates the condensation of monomeric ortho-silicic acid [Si(OH)4] to polymeric amorphous silica (n•SiO2; formation of Si–O–Si bonds). In a second further reaction, silicatein acts as a hydrolase, which catalyzes the hydrolytic cleavage of the alkoxysilane compound TEOS [Si(OR)4; R = ethyl] (cleavage of Si–O–C bonds) allowing a bioinspired synthesis of artificial amorphous silica-based (nano)materials for biomedical applications. Shown in the center (enclosed in a ring) are the two amino acids of the catalytic triad, Ser and His, which interact with silicic acid and mediate the condensation. (A) Silicatein core protein of an aster from G. cydonium. (B) Complete sponge aster after incubation of silicatein with TEOS. (C) A spicule from S. domuncula with the axial canal (ac) centered by the silicatein-containing axial filament (af). (D) An axial filament with thorny silica (sil) protrusions. (E) Silicatein deposits layered onto thorny template, after reaction with TEOS. (F) Microspheres lacking silicatein after incubation with TEOS. (G) Silicatein-containing microspheres after TEOS incubation.](/cms/asset/1b03aff9-dbfa-4312-ae03-fa2d131c3fbf/dijn_a_12300375_f0003_c.jpg)
The biosilica synthesized by silicatein is deposited onto a structured organic template, the enzyme ( and ) which also forms a mature siliceous spicule. Consequently, silicatein existing in the axial canal (the axial filament) predetermines the morphology of the spicules ( and ). If the spicules are thorned, the surface of the silicatein axial filament shows thorny protrusions (). Using TEOS as a synthetic substrate, silicatein has been used to coat various surfaces/templates (), such as microspheres ( and ).
Biosilica: Bioinspired Application as a Morphogenetic Polymer
The field of application of (bio)silica and the bioinspired/biomimetic materials and biomineral structures based on it is wide, ranging from the use in regenerative medicine, improved bacterial fermentation to optical fibers and even as a construction model for architectural buildings.
Biosilica has been considered as a paradigm for biological mineral morphogenesis and evolution not only in spongesCitation80 but also in diatoms.Citation81 In both systems, the morphology of the structures formed is driven by genetically controlled mineralization. The morphogenetic biosilica effect is attributed to its gene-inducing effect.Citation82 In a series of applied experiments, it could be demonstrated that biosilica is a potent material for osteogenic stimulation and differentiation.
Biosilica: A Generic Template for Bone and Tooth Repair
The property of biosilica to be morphogenetically active and supports and likewise accelerates regeneration/repair processes in humans was proven in vitro and in animals (rabbits).Citation80,Citation83 In a first bioinspired approach, the sponge biosilica was applied in in vitro cell systems to evaluate and then to document an increased mineralization potential, first by measuring the expression of respective genes, like the structural proteins within the tooth matrix proteins, amelogenin, ameloblastin, and enamelin, which were strongly upregulated.Citation80 The rationale for this line of study came from Carlisle in her report in 1972.Citation84 Carlisle showed that chicks fed a silicon-rich diet grew and developed substantially faster than control animals. These data were confirmed laterCitation85 by demonstrating that the bone strength of broilers is strongly enhanced by dietary supplementation with bioavailable silicon. In continuation, in a series of in vitro studies, it was measured that both biosilica and silicatein induce the growth of mineralizing cells on silicatein/biosilica-coated matrices and cause under these conditions a strong increase in hydroxyapatite (HA) mineral formation;Citation86 in the absence of biosilica, no HA mineral nodules were detected.
Consequently, animal studies were performed using New Zealand White rabbits by inserting 600-µm-large microspheres pressed into pellets () and implanted into the anterior patella (between the medial and lateral femoral condyles). As an active ingredient, either silicatein (0.8 µg g‒1) or biosilica (100 µg g‒1) was added (). The biosilica pellets, added into PLGA (poly(d,l-lactide-co-glycolide))-based microspheres, were implanted.Citation83 After termination of the study, the implants reacted strongly with OsteoImage (), reflecting the increased regeneration/mineralization activity caused by biosilica.Citation87 The PLGA-based microspheres, supplemented with biosilica, were placed into the drilled space of the patellar grooves of the rabbits ().Citation83 After a total healing period of 100 days, the progression of bone regeneration was inspected by in vivo staining the samples with oxytetracycline dihydrate. The intensity of the blue fluorescence along a bone slide reflects the degree of regeneration.Citation88 The images showed that in the control samples with microspheres, supplemented with β-TCP (β-tricalcium phosphate), only a homogenous tissue is visible around the microspheres that did not brightly stain with oxytetracycline dihydrate and was not flashing under ultraviolet light (). In contrast, in the sections with the implanted biosilica-supplemented spheres, the microspheres are surrounded by bone tissue and this area lights brightly up almost homogenously in blue ().
Figure 4 Morphogenetic activity of biosilica in animal experiments (rabbits) - regeneration of bone in holes drilled into patellar grooves. (A) Microspheres embedded in pellets and (B) biosilica prepared by silicatein for the implant experiments. (C) The microspheres were fabricated together with biosilica. Staining of the mineral nodules formed in rabbits after insertion for 5 days. Staining with OsteoImage (fluorescence flashing). In the (D) femoral implant experiments, new bone formation was detected after in vivo staining with (E and F) oxytetracycline dihydrate under UV light. A striking difference was found between β-TCP controls (E) and biosilica-supplemented microspheres (F). Adapted from Bone, volume 67, Wang SF, Wang XH, Draenert FG, et al. Bioactive and biodegradable silica biomaterial for bone regeneration. 292–304. © 2014, with permissions from Elsevier Inc.Citation83

In general, crystalline biominerals such as bone HA are formed from the amorphous precursors.Citation66 The mechanism underlying the morphogenetic activity of biosilica is not fully understood. The release of orthosilicic acid from amorphous biominerals could contribute to the stimulating effect on the mineralization of bone-forming cells as in bioglass. The mechanism proposed for bioglass involves an exchange of Na+ and Ca2+ with H+ ions, leading to the formation of silanol groups and an increase in the surrounding concentration of OH– ions.Citation89 The cleavage of Si–O–Si bonds by OH– then results in the release of orthosilicic acid and the formation/re-condensation of further silanol groups, which – similar to biosilica – leads to a hydrated, silica-rich layer on the glass surface, which is a suitable matrix for mixed carbonated HA deposition by invading Ca2+, PO43–, OH– and CO32– ions. It is known that low silicate concentrations (0.05–0.5 mM) promote HA nucleation.Citation90 Silica has also been shown to stabilize and prevent the crystallization of ACC, the precursor of amorphous calcium phosphate (ACP), and of crystalline HA.Citation91 In addition, modeling studies showed that the silanol groups of cyclic trisilicic acid motifs on the silica surface stereochemically mimic the Ca2+-binding HA nucleation site on bone sialoprotein (BSP).Citation92–94
These data underscore that biosilica shows morphogenetic activity on bone-forming cells not only in vitro but also in vivo, in animal experiments.Citation95
Inorganic Polyphosphate
The second physiological inorganic polymer, identified in sponges,Citation96 that has attracted increasing attention, is polyphosphate (polyP). Originally, polyP was identified in bacteria and yeast (for a review, see Ref.Citation97), and later in animals, like in spongesCitation96 and higher vertebrates/humans.Citation98 This apparently ubiquitous polymerCitation99,Citation100 exhibits a unique property that no other inorganic material useful for human therapy possesses - the delivery of metabolic energy. PolyP () has emerged as a prime example of a physiological polymer that not only fulfills the structural but also the energy-supplying requirements for a successful biomedical regeneration process.
Figure 5 (A) Chemical structure of polyphosphate (polyP). (B) Intracellular synthesis of polyP. Glucose is the body’s main source of energy. After cellular uptake, glucose is metabolized to pyruvate during glycolysis, which is then channeled via the voltage-dependent anion channel (VDAC) to the mitochondrial intermembrane space and then to the matrix, where it undergoes oxidative metabolism. The generated reduced coenzymes (NADH and FADH2) drive redox reactions and the electron transport chain builds an electrochemical transmembrane proton gradient whose energy is converted to ATP. ATP is released into the cytoplasm via the adenine nucleotide translocator-2 (ANT2) and VDAC. Subsequently, ATP is channeled through the vacuolar transporter-chaperone complex (VTC) in the acidocalcisomal membrane, which functions in yeast as a polyP polymerase. From the acidocalcisomes in the megakaryocyte-platelets, polyP is released to the extracellular space either as soluble Na-polyP or as membrane-associated Ca-polyP nanoparticles (Ca-polyP-NP). Adapted from Müller WEG, Schröder HC, Wang XH. Inorganic polyphosphates as storage for and generator of metabolic energy in the extracellular matrix. Chem Rev. 2019;119:12337–12374. © 2019 American Chemical Society. Creative Commons.Citation5

Physiologically, polyP is stored intracellularly in organelles, now termed acidocalcisomes, which have been intensively studied in trypanosomatids, protozoan parasites.Citation101,Citation102 Earlier they have been identified as metachromatic granulesCitation103 or volutin granules.Citation104 In the blood platelets, polyP is accumulated in the dense granules, as identified by Ruiz et al.Citation105 The discovery that ATP and polyP harbor metabolic energy came from Meyerhof and Lohmann (cited in Ref.Citation106). The unequivocal identification of the chemical structure was described by Lohmann and Langen.Citation107,Citation108
Recently, as described later, the proof-of-concept of the therapeutic benefits in the clinic for polyP could even be successfully provided. As a hydrogel, the natural polymer not only provides a platform for cell proliferation, cell differentiation, and cell migration but also provides the metabolic energy required for maintaining the molecular and supramolecular organization of the extracellular matrix and cell function.Citation48,Citation49
Polyphosphate: Cell-Based Synthesis
The reaction chain by which polyP is synthesized in bacterial cells is fairly well understood (reviewed in Ref.Citation109). Less is known about polyP synthesis in higher vertebrate cells.Citation98,Citation110 For the synthesis of polyP in mammals, ATP is required, which is generated intracellularly in the mitochondria ().Citation111–113 There, in the respiratory chain, ATP formation is linked with complex V, the F1Fo-ATPase. For the chemical synthesis of the phosphoanhydride bonds in ATP, heating to >100°C for ~10 h is required.Citation114 Accordingly, for an intracellular, physiological synthesis of ATP at 37°C, with its high-energy anhydride bonds, an activation energy of 110 kJ mol‒1 has to be expended, as determined using Arrhenius plot.Citation115 Therefore, the reactions in the mitochondria must be mediated by enzymes. It is the ATP synthase, localized in the inner mitochondrial membrane, which catalyzes the synthesis of ATP from ADP. In turn, the biosynthetic pathway for the formation of polyP with its >30 high-energy anhydride bonds must involve enzymes as well. In mammalian systems, the genuine polyP-synthesizing enzyme has not yet been discovered. Experimental evidence in yeast suggests that polyP is formed enzymatically from ATP during import into the acidocalcisomes ().Citation116,Citation117 Indicative is the fact that, in the dense granules of the platelets that correspond to the yeast acidocalcisomes, the concentrations of ADP (600 mM), ATP (400 mM), and pyrophosphate (300 mM) are exceedingly high,Citation118 while the polyP content is comparably low (130 mM; based on Pi).Citation119 Therefore, it might be assumed that phosphatases or phosphotransferases present in plateletsCitation120,Citation121 could be involved in polyP synthesis through backward reactions from ATP, driven by these enzymes. From the platelets, polyP is released in two forms, either in a soluble form, as a chain with Na+ as counterion, or in an “insoluble” form, as a NP with Ca2+ as counterion ().Citation118,Citation122 Since polyP compartmentalized into the acidic dense granules (pH 5.4) is released in a controlled manner,Citation123,Citation124 it is likely that the Ca2+ gradient (Ca2+ concentration in the dense granules is 2.2 M)Citation119 determines the formation of the two forms of polyP as proposed.Citation118
After full platelet activation, the concentration of polyP in the blood is relatively high with 0.5 to 3 μg mL‒1.Citation118,Citation125 There, the polymer has a physiological chain length of ~50 phosphate (Pi) units. In the particulate Ca2+ form, the polyP chains are longer with ~250 Pi units. However, this value is very variable since there are high levels of ALP in the blood, which hydrolyzes the polymer from the chain termini as an exopolyphosphatase.Citation126
A biocompatible polymer can be particularly beneficial to patients when transported within the body to the target site for tissue repair. Here, too, polyP follows the route of a biomimetic medical compound. In vivo, under physiological conditions, polyP is efficiently distributed to the injured sites in the body with the blood circulation. There, the polymer is delivered by the blood platelets to the damaged tissue regions, where the polymer initiates the regeneration process.Citation127 In fact, platelets with the stored polyP are hallmarks of regeneration.Citation128 It was Julius Bizzozero who discovered platelets as small medicinal pellets of a size between 1 and 2 µm.Citation129 He described them as the initiators of blood clotting. PolyP is a major constituent of platelets.Citation105 Using these vehicles, polyP is distributed throughout the body in order to onset regeneration/repair. The distribution of polyP is flanked by macrophages, which bind or internalize polyP, independently of their signaling roles, via their P2Y1 and RAGE receptors.Citation130 Besides acting on the clotting cascade and enhancing hemostasis, platelets are activated and release polyP into the extracellular space when certain growth factors, such as epidermal growth factor or platelet-derived growth factor, are released.Citation122 In this environment, platelets bind directly to exposed collagen fibers, as well as to von Willebrand factor, fibronectin, and other adhesive proteins.Citation131 This efficient distribution mechanism of polyP in the body contributes to the prominent position that polyP has achieved in regenerative medicine, as described later.
Polyphosphate: Chemical Synthesis
Chemically, polyP can be prepared in sufficiently large quantities.Citation132 Na-polyP with its energy-rich phosphoanhydride bonds () is obtained by melting of NaH2PO4 at temperatures up to 700°C (). For the fabrication of Ca-polyP, both an enzymatic/wet chemical approachCitation133 and a calcination process have been introduced.Citation134 By shortening the high-temperature processing protocol, it is possible to prepare both amorphous Na-polyP and Ca-polyP-NP almost in parallel ( and ). The process runs at 700°C. At the end of the polycondensation reaction, polyP, with a chain length of ~50–100 Pi units, is supplemented with CaCl2 and heated for an additional period of time to obtain the amorphous Ca-polyP-NP with a diameter of ~100 nm (). When Na-polyP is synthesized, the material is ground to 50 µm powder (). Different steps of the procedure, starting from NaH2PO4, are summarized in .
Figure 6 Chemical preparation of polyP. PolyP is prepared by heating the starting material NaH2PO4 to 700°C. Both amorphous (A and B) Na-polyP, and (C and D) Ca-polyP-NP can be prepared in parallel. (A) Na-polyP is formed during heating to 700°C. The glass-like melt is (B) ground to Na-polyP powder. Ca-polyP-NP is prepared from Na-polyP by addition of CaCl2 (C and D). By this, polyP of a physiological chain length of ~50–100 Pi units (Na-polyP50–100) is obtained. (E-1–E-4) Preparation process of Na-polyP and Ca-polyP-NP. (E-1) Start of preparation; (E-2) sliding in the furnace; (E-3) The molten glass poured from the crucible onto (E-4) a steel plate (images with infrared camera).

Polyphosphate: A Biomimetic Molecule for Human Therapy
PolyP as a polyanion can be present in tissues as salt with various cations as counterions. The biological regeneration function of polyP differs depending on the counterion chosen. In addition, the functional activity of polyP depends on the form of the polyP salts, which can exist both in a soluble form, such as the sodium salt (Na-polyP), and in an insoluble nanoparticulate form that can be used as a storage (depot) form, such as salts with various divalent cations, eg, Ca2+ ions. The latter salts can also form a gel-like coacervate phase, a physiologically active form, as outlined later.
Cation-Specificity of Polyphosphate: A Smart Nano/Micro Biomaterial
Besides the thermal method described above, polyP can be fabricated in a biomimetic way as nanoparticles (NP) as in vivo in human cells using a wet chemical procedure.Citation118,Citation135 The biological activity of the polyP nanoparticles is determined, at least in part, by the counter-cations used for a particular scaffold (). For cartilage repair, Ca2+- and also Mg2+-polyP complexes are more suitable,Citation136 while for bone regeneration, the Ca-polyP forms might be favored due to their higher stability. In addition, the Sr2+-polyP complex shows strong regenerative activity both in vitro and in animal experiments.Citation137 Drugs that are effective, eg, against bone tumors/metastases, such as the bisphosphonate zoledronic acid, can be conveniently integrated into the Ca-polyP-NP and used as implant particles.Citation138 For another application target, chronic wound healing,Citation48,Citation49 a combination of Na-polyP and Ca-polyP-NP acts most efficiently on the regeneration process. In addition, polyP particles, when applied in an aerosol form, have a protective effect on the respiratory epithelium, after conversion into the coacervate form upon contact with mucin, the protein of the airway mucus.Citation139 As a result, viral particles such as SARS-CoV-2 are entrapped and inactivated, in addition to a direct inhibitory effect on this virus.Citation140,Citation141
Figure 7 PolyP is a genuine, smart nano/micro biomaterial whose properties and applications depend on the selected counterion. PolyP-NP, polyphosphate nanoparticle. Reproduced with permission from Wang XH, Schröder HC, Müller WEG. Amorphous polyphosphate, a smart bioinspired nano-/bio-material for bone and cartilage regeneration: Towards a new paradigm in tissue engineering. J Mat Chem B. 2018;6:2385–2412. © 2018 The Royal Society of Chemistry. Creative Commons.Citation142

Polyphosphate Coacervate Formation
It is an exceptional feature of polyP (as Na-polyP) to undergo coacervation in the presence of Ca2+ or other divalent cations, at physiological pH. In principle, polyP nanoparticles, including non-processed Ca-polyP-NP, are biologically inert. They have to be converted into a biologized form, a polyP-coacervate. During this phase transition, polyP provides its physiological functions. At pH 7, Ca2+ together with polyP forms a viscous aqueous phase, the coacervate ( and ). These aggregates are formed through liquid–liquid phase separation, resulting in a denser phase and a dilute phase that are in thermodynamic equilibrium.Citation143 In the coacervate phase, polyP shows morphogenetic activity as well as its function to generate ATP.Citation144 In our approach, the two phases and their interconversion were studied using in silico simulation studies, as well as (physico)chemical analyses. The data showed that Ca-polyP coacervate formation occurs at pH 7 and is slower, compared to Ca-polyP-NP formation at pH 10 ( and ). Interestingly, if CaCl2 is dropped into a Na-polyP solution at pH 7, the Ca-polyP coacervate initially forms and becomes then converted to NP at pH 10. Conversely, when Na-polyP is added to a CaCl2 solution, only the coacervate phase is obtained at both pH values.Citation144
Figure 8 PolyP / Ca-polyP phases. (A–D) At pH 7, the polyP coacervate is formed from Ca2+ and Na-polyP. During this process, the aqueous liquid-liquid gelatinous phase envelops bacteria such as (B) E. coli and/or (D) attracts mesenchymal stem cells (MSC) and allows them to nest and differentiate there. (E and F) NP formed from Ca2+ and Na-polyP at pH 10.

The coacervate is biocompatible and allows the infiltration of mesenchymal stem cells (MSC) into the gel matrix, where polyP promotes cell proliferation and differentiation;Citation143 the MSC become completely embedded in the matrix (). When the coacervate (Ca2+-polyP) forms during the liquid–liquid phase separation, different local densities arise causing turbulences during which bacteria, such as E. coli, are enwrapped () and then killed.Citation49
PolyP particles are taken up by endocytosis, as determined by inhibition studies with trifluoperazine dihydrochloride.Citation51 Intracellularly, the particles begin to transform into a coacervate. The free polyanionic polyP (not in a salt form) is not able to traverse cell membranes. Only after caging the polymer, eg, into a guanidinium/oligocarbonate vehicle, polyP can be channeled into the cells.Citation145
Supramolecular Extracellular Matrix Organization: Importance of Polyphosphate Coacervation and Energy Requirement
Biomimetic materials used for soft or hard tissue regeneration can be classified according to the four tissue categories: covering tissue, connective/and supporting tissue, muscle tissue, and nervous tissue.Citation146 The ECM of the different organs has a different percentage of cells, such as the liver with a high (>90%) and cartilage with a low cell density (<10%). A common feature of their ECMes is that the cells are embedded into a fibrillar, often collagenous, hydrogel, which allows architectural stability and simultaneous diffusion of nutrients as well as cell movement. To ensure biocompatibility, a physiological or close-to-physiological hydrogel is preferably used in regenerative medicine. Surely, such hydrogels can elicit signals stimulating tissue regeneration. Normally, hydrogels lack the ability to directly supply the cells with metabolic energy, preferably in the form of ATP.Citation5
Supramolecular hydrogels have been introduced into regenerative medicine because of their unique dynamic properties for self-healing and injectability based on the non-covalent crosslinking organization of their macromolecules. Hydrogels based on this fabrication technology are held together by forces arising from non-covalent bonds such as electrostatic interactions, hydrogen bonds, metal coordination, aromatic stacking, and hydrophobic and van der Waals forces ().Citation147,Citation148 The individual molecules are synthesized by covalent bond formation (“Molecular chemistry”) and become subsequently organized into complex supramolecular chemistry-based systems. There (usually) more than one chemical unit interacts via non-covalent bonds, as in the case of receptors or enzymes. Consequently, ligands or substrates in the next higher hierarchy level typically associate with receptors in turn via noncovalent bonds and form an active state of the system. In this complex arrangement, signal transduction pathways (with the respective ligands) are initiated or enzymatic reactions follow, through which new covalent bond(s) are introduced into the reaction chain.Citation148
Figure 9 Formation of a physiological hydrogel in mammalian cells. (A) The different steps of physiological hydrogel formation through the steps of molecular chemistry (formation of covalent bonds) and supramolecular chemistry (non-covalent bonds) to the biomimetic synthesis of a coacervate (pH 7) or NP (pH 10). During these reactions and pathways, polyP is distributed to the target tissues where it serves as a source for Pi delivery, metabolic energy supply, and bacterial enwrapping. (B) The bidirectional functional activity of polyP. Na-polyP forms at pH 7 a coacervate by compartmentation of polyP molecules after Na+/Ca2+ exchange. The transformation of Ca-polyP-NP, the storage (depot) form of the polymer, is facilitated in the presence of peptides. The coacervate is the functional form of polyP, which exhibits morphogenetic activity. During the progression of Ca-polyP-NP to the coacervate, the polymer is (partially) hydrolyzed by ALP under formation of ADP, which in turn is up-phosphorylated to ATP to bio-fuel the cells.

Due to its ability to form a coacervate promoting immigration and embedding of cells in the presence of Ca2+ or other divalent cations, as well as its ability to induce stem cell growth and differentiation, polyP is an excellent material for mimicking the conditions in the ECM of human tissues. In particular, polyP has the unique property of not only exerting a morphogenic effect but also providing the metabolic energy needed for tissue regeneration (). Organic polymers used with polyP included negatively charged polysaccharides such as N,O-carboxymethyl chitosan (N,O-CMC), alginate,Citation149 chondroitin sulfate,Citation150 hyaluronic acid,Citation136 and karaya gum,Citation151 as well as collagenCitation150 and poly(vinyl alcohol) (PVA).Citation151 These in vitro results were confirmed by animal experiments (rat muscle) showing a replacement of the implants by granulation tissue already after an implantation period of 2 to 4 weeks.Citation151 The most advanced polyP-based bio-ink, which provides metabolic energy in addition to morphogenetic activity, was developed using an N,O-CMC/alginate/gelatin hydrogel enriched with soluble Na-polyP- and Ca-polyP-NP (see below). This bio-ink enabled the successful 3D bioprinting of MSCs that remained functionally active and able to migrate, grow, and differentiate to mineralizing osteoblasts.Citation152
In tissues, the organization of both the intra- and extracellular supramolecular structures requires an energy input.Citation153,Citation154 Self-assembly reactions are usually considered to be independent of energy supply; however, the subsequent downstream processes certainly require ATP, such as the organization of collagen fibers into bundles.Citation155 Until recently, the source of metabolic energy in the ECM was uncertain. It should be mentioned that the concentration of ATP in the extracellular space is only ~10 nM, while intracellularly the ATP level is high, varying between 0.5 and 5 mM depending on the cell type.Citation156,Citation157 ATP is required for many functions in the ECM. There, ATP is involved in the cell-cell communication, particularly in the nervous system, but also for the activation of T cell receptor (TCR) signaling.Citation158 ATP is also required, as a chemotactic agent, for endothelial cells to direct them to micro-vessels along an ATP gradient.Citation51 In addition, ATP-consuming kinase reactions also run extracellularly and chaperones are also present (reviewed in Ref.Citation5).
Physiological Polyphosphate: A Key in Regenerative Medicine
Autologous platelet concentrate, platelet-rich plasma, is rich in polyP and is used for the prevention and treatment of bone and cartilage defects and for wound closure, especially in small ulcers.Citation159 Interestingly, fibrin (which contains platelets with their polyP) shows the same accelerating effect on regeneration processes.Citation160
So far, the main focus of the polyP-correlated biomimetic application for human therapy is in the field of repair of osteochondral (bone and cartilage) and skin and soft tissue (chronic wounds). The common denominator for these targets are the MSC which promote both bone/cartilageCitation161,Citation162 and wound repair.Citation163 Furthermore, in vitro studies have shown that polyP is a potent inducer of cell proliferation and differentiationCitation164–166 as well as mineralization.Citation167 There are two features that qualify the metabolizable polyP as a distinguished polymer in regenerative medicine; first, it is based on phosphate, an osteoconductive biomaterial required for bone cell growth,Citation168 and second, the inherent function of polyP to dissipate metabolic activity.Citation5,Citation99
Principal Distinctive Feature of Polyphosphate: Generation of Metabolic Energy
The distinguished feature of polyP is that its orthophosphate units are condensed via energy-rich phosphoanhydride bonds, forming a linear polymer with a physiological chain length of ~50–100 Pi units.Citation169 The energy (Gibbs free energy; ΔG0) stored in each bond and released during hydrolysis from the ends of the polymer is about −30 kJ mol‒1.Citation170 The law of energy-constancy in a closed system implies that energy is converted into different forms, while the sum of energy remains constant. The energy simply converts from one form to another. In a biochemical reaction, a part of the energy is dissipated as heat and the rest is used as metabolic energy that drives biochemical reactions.Citation171 In turn, ΔG0 for ADP hydrolysis is about −30 kJ mol‒1 and for ATP (both energy-rich bonds), it is about −60 kJ mol‒1 (). In contrast, polyP with a chain length of, eg, 40 Pi units (39 energy-rich bonds) comprises a ΔG0 of approximately −1170 kJ mol‒1. Experimental evidence from SaOS-2 cells, after incubation with polyP, showed that an increased amount of energy-rich nucleotides (ADP and ATP) accumulates extracellularly.Citation50 During the incubation of the cells with polyP, vesicles with both ALP and adenylate kinase (ADK) activities are released. These two enzymes are plasma membrane associated.Citation172,Citation173 Inhibition studies indicate that initially ADP is formed (during degradation of polyP with ALP), which is further up-phosphorylated to ATP by ADK.Citation51 So far, a quantitative evaluation of the yield of ATP production from polyP is missing.
Figure 10 Role of ATP during bone mineralization. (A) PolyP acts as an extracellular generator for ATP (for polyP40: ΔG0 −1170 kJ mol−1). A polyP molecule can generate a multifold of the energy present in ADP (−30 kJ mol−1) or ATP (−60 kJ mol−1). (B) Bone mineralization is an energy-dependent process and proceeds extracellularly (Scheme). This process can be divided (left panel) into an ATP generation section and a bioseed formation for initiating of bone/HA synthesis (right panel). On the surface of osteoblasts, polyP is enzymatically hydrolyzed by ALP to form ADP from AMP. During cleavage of the high-energy acid anhydride bonds in polyPn, Gibbs free energy is released; in each step, the polyP chain is shortened by one Pi unit (polyPn-1). The enzyme ADK forms one mole of ATP from two moles of ADP. This process is coupled to the Fam20C kinase, which phosphorylates the dentin matrix protein 1 (MP-1), among other proteins. The phosphorylated MP-1 is an essential key in bone mineralization. The molecule acts as seed for the deposition of ACP (amorphous Ca-phosphate). Subsequently, after hydrolysis of polyP via ALP, ACP transforms into crystalline hydroxyapatite (HA).

Kinases: An Essential Transfer Element
The outlined generation of ATP from polyP would cease when an equilibrium defined by the equilibrium constants between polyP, AMP, ADP, and ATP is reached. Consequently, metabolic energy must flow out of this equilibrium to keep the system running (). At this point, kinases achieve a crucial function. Extracellularly, a number of proteins involved in mineralization are phosphorylated, in particular osteopontin, dentin matrix protein-1, and dentin sialophosphoprotein.Citation174,Citation175 As the only (major) enzyme that catalyzes the phosphorylation of these proteins, the Golgi-localized protein kinase Fam20C, a serine kinase, has been identified.Citation176 Failure of the function of Fam20C often leads to lethal osteosclerotic bone dysplasia, known as the Raine syndrome.Citation177
Repair of Osseous Hard Tissues by Polyphosphate
Already Lowenstam suggested that the calcareous invertebrate skeletons from the Early Cambrian ≈560 mya are made of ACC, a view that has been later corroborated by dissolution experiments.Citation178,Citation179 In sponges, the class of calcareous sponges (Calcarea) forms their hard skeletons of Ca-carbonate, which is individually composed of calcite formed enzymatically with the sponge-specific carbonic anhydrase.Citation29 The inorganic biomineral of the otoliths in the human inner ear also completely consists of Ca-carbonate, which is present in the form of calcite.Citation180 It is not yet clear which enzymatic processes are involved in Ca-carbonate biosynthesis in human systems. Experiments with human osteogenic SaOS-2 cells suggested that in this system the carbonic anhydrase CA-IX, a cytosolic enzyme localized on cell membranes, is upregulated after exposure of SaOS-2 cells to bicarbonate.Citation142 HA/bone formation proceeds through the phases of hyaline cartilage (hyaline, translucent cartilage), “smooth” cartilage (smooth type of connective tissue), endochondral ossification (embryonic bone tissue formation), appositional growth (increase in diameter), and finally, maturation of the mineral deposits to crystalline HA ().Citation181 Element mapping using SaOS-2 cells by scanning electron microscopy of the surfaces of mineral deposits showed that the Ca minerals show signals for phosphorus, as expected, but initially also for the element carbon (). These data corroborate the assumption that mineralization in human bone also passes through the Ca-carbonate stage before the conversion to Ca-phosphate can take place.Citation42 Therefore, it was proposed that the initial step in bone formation starts with the deposition of Ca-carbonate, perhaps catalyzed by the carbonic anhydrase. The matrix onto which bone mineral deposition takes place is a collagen meshwork. Surely, the first mineral deposits are amorphous.Citation66 The transformation of the enzymatically formed (carbonic anhydrase-mediated) amorphous Ca-carbonate (ACC) towards amorphous Ca-phosphate (ACP) proceeds as an exergonic reaction and is not enzyme-driven.Citation182,Citation183 After that, ACP transforms into crystalline hydroxyapatite ().
Figure 11 Ca-phosphate/bone formation. (A) Steps during bone formation: amorphous Ca-carbonate (ACC), amorphous Ca-phosphate (ACP) and finally crystalline Ca-phosphate/hydroxyapatite (HA). The different phases of bone initiation, growth and maturation are outlined in the upper trace. At least the final step, the transition from ACC to ACP, is driven enzymatically by ALP. There, the enzyme ALP hydrolyzes polyP and, by this, eliminates the stabilizing polymer polyP from the amorphous ACC and ACP phases. This process leads to the maturation of ACP to crystalline HA. During this process, the initial endergonic reactions are pulled by the subsequent exergonic processes. (B) Element mapping of the surfaces of Ca mineral deposits formed on human osteoblast-like SaOS-2 cells (B-1; SEM) for the elements oxygen (B-2), phosphorus (B-3) and carbon (B-4). Adapted with permission from Wang XH, Schröder HC, Müller WEG. Amorphous polyphosphate, a smart bioinspired nano-/bio-material for bone and cartilage regeneration: Towards a new paradigm in tissue engineering. J Mat Chem B. 2018;6:2385–2412. © 2018 The Royal Society of Chemistry. Creative Commons.Citation142

A huge amount of phosphate is required for bone formation. Based on in vitro data, it had been suggested that it is the organic phosphate, β-glycerophosphate, which, after hydrolysis by the ALP, provides the inorganic phosphate for bone formation.Citation184 Later, when the crucial role of the platelets in bone synthesis and bone remodeling was revealed,Citation185 it became evident that polyP is the main source of Pi.Citation186 In this study, it was also reported that the gene expression of ALP,Citation187 a marker enzyme for bone formation, is upregulated after polyP exposure.Citation186 The enzyme-driven mineralization chain then proceeds to the ACP step and allows an appositional growth of the bone.Citation188 In ACP, polyP acts as a stabilizing polymer of the amorphous phase during its transition to crystalline HA.Citation167 After hydrolysis of polyP by the ALP, maturation from amorphous to crystalline Ca-phosphate can take place.Citation69
Following these new scientific insights, polyP has been tested for its regenerative activity in vivo . Using the rat calvarial defect system as a model of bone repair, the polymer was integrated into PLGA microspheres. After eight weeks, the material was fully resorbed and the bone defect was completely restored, while β-TCP caused only limited repair. A comparative gene expression study revealed that polyP also significantly upregulates the expression of the collagen gene, in contrast to the β-TCP material.Citation189
As mentioned above, crystalline biomaterials have only limited morphogenetic, regenerative activity, while amorphous materials exhibit regenerative potential.Citation66 This differential effect can be demonstrated with the polyP-stabilized amorphous phase of Ca-phosphate, ACP. In the presence of small amounts of polyP (5% by weight), ACP still proceeds to the crystalline Ca-phosphate state (CCP). However, after increasing the polyP level to 15%, the ACP mineral remains frozen in the amorphous state.Citation167 Therefore, a comparative animal study was performed with crystalline Ca-phosphate (containing 5% polyP), cCa(5)polyP, versus amorphous ACP/15% polyP particles (aCa(15)polyP).Citation30 The Ca-phosphate formulations were integrated into PLGA microspheres. The starting material cCa(5)polyP shows a hexagonal columnar crystal morphologyCitation190 (). In the in vivo model (rabbit calvarial bone defect model), cCa(5)polyP only marginally repaired the bone defect after 6 weeks (). Interesting was the fact that the association (acceptance) of the microspheres to the tissue environment with cCa(5)polyP was very poor or even absent ().Citation30 In comparison to crystalline ACP, the amorphous ACP (aCa(15)polyP) deposits appear as globular spheres (). Those ACP deposits accelerate the processes of migration, microvascularization, and mineralization in SaOS-2 cells. The repair process was dramatic; the newly formed tissue covered the regenerating zone and showed only remnants of the microspheres (). Even more, the microspheres in the regenerating area of the aCa(15)polyP-treated animal group readily attracted and interacted with the newly formed collagen bundles (). These data show that aCa(15)polyP combines osteoinductive activity with the morphogenetic potential to build the HA calvaria.
Figure 12 Bone healing with crystalline Ca-phosphate (cCa(5)polyP; containing 5% polyP). (A) Crystalline Ca-phosphate formed during aging in the presence of only 5% polyP. (B and C) Healing studies of encapsulated Ca-phosphate microspheres (MS) in vivo using the rabbit calvarial bone defect model. (B) After 6 weeks of insertion of MS filled with cCa(5)polyP, only a slightly repaired bone defect was seen in animals; (C) the interaction between the microspheres (crystalline phosphate implant) and the adjacent bone is poor. In contrast to the crystalline Ca-phosphate, containing only 5% polyP, (D) addition of 15% polyP to the starting substrates for the preparation of Ca-phosphate, [(NH4)2 HPO4 and CaCl2], freezes the product Ca-phosphate at the amorphous phase. (E) Those MS elicit a strong healing process, the MS fused with adjacent regenerating bone and (F) formed a tight collagen (co) around the MS. Adapted with permission from Acta Biomater. Volume 118, Müller WEG, Ackermann M, Al-Nawas B, etal. Amplified morphogenetic and bone forming activity of amorphous versus crystalline calcium phosphate/polyphosphate. 233–247, copyright 2020, with permission from Elsevier.Citation30
![Figure 12 Bone healing with crystalline Ca-phosphate (cCa(5)polyP; containing 5% polyP). (A) Crystalline Ca-phosphate formed during aging in the presence of only 5% polyP. (B and C) Healing studies of encapsulated Ca-phosphate microspheres (MS) in vivo using the rabbit calvarial bone defect model. (B) After 6 weeks of insertion of MS filled with cCa(5)polyP, only a slightly repaired bone defect was seen in animals; (C) the interaction between the microspheres (crystalline phosphate implant) and the adjacent bone is poor. In contrast to the crystalline Ca-phosphate, containing only 5% polyP, (D) addition of 15% polyP to the starting substrates for the preparation of Ca-phosphate, [(NH4)2 HPO4 and CaCl2], freezes the product Ca-phosphate at the amorphous phase. (E) Those MS elicit a strong healing process, the MS fused with adjacent regenerating bone and (F) formed a tight collagen (co) around the MS. Adapted with permission from Acta Biomater. Volume 118, Müller WEG, Ackermann M, Al-Nawas B, etal. Amplified morphogenetic and bone forming activity of amorphous versus crystalline calcium phosphate/polyphosphate. 233–247, copyright 2020, with permission from Elsevier.Citation30](/cms/asset/144a06a7-12ff-4b90-bc67-0c821b68a835/dijn_a_12300375_f0012_c.jpg)
Repair of Soft Tissue: Chronic Wounds
It is established that the supply of blood, and hence platelets, is crucial for bone tissue development and repair, and for healing of osteoporotic fractures.Citation191–193 During these processes, platelets surround the affected site and become activated, and release growth factors and certainly also polyP. Even more obvious is the role of platelets in wound healing. It has been reported that platelet deficiency plays an adverse, critical role in skin healing and that its failure prevents healing of chronic wounds.Citation194 In all phases of wound repair, hemostasis, followed by inflammatory reactions, up to the proliferative and remodeling phase, platelets decisively promote the regeneration process.Citation195
Wound healing is energy-intensive. Most of the metabolic energy is consumed for positioning of new cells in the wound area.Citation196 The epithelial cell motility, which is controlled by different members of the Rho family of GTPases, which act as molecular switches, consumes energy-rich nucleotides.Citation197 In addition, wound healing is decisively dependent on new vessel formation, beginning with a polyP-induced microvascular network,Citation198 which is required for optimal tissue perfusion and oxygenation. Finally, ATP acts as a stimulus for the endothelial cells to follow an ATP gradient and guide the cells via an autocrine sensing mechanism during the initial chemotactic formation of 100 µm circular endothelial cell rings.Citation51,Citation199,Citation200 This ATP originates from glycolysis and not from the respiratory chain.Citation201
Chronic wounds are becoming an increasingly common clinical problem, especially in patients with diabetes mellitus.Citation202 In these patients, severe hyperglycemia due to an impaired insulin supply leads to reduced gene expression of enzymes involved in mitochondrial oxidative phosphorylation.Citation203,Citation204 In addition, an imbalance between angiogenic factors, angio-inhibitory factors, and abnormal apoptotic potential is observed, which reduces the proliferation of cells and their remodeling capacity. In diabetic wounds, the cells remain in a chronic inflammatory state and do not switch to the proliferation and remodeling phase. In turn, the use of drugs that generate metabolic energy in the wound is indicated. Besides ATPCitation205 and a hydrogel supplemented with 0.02% adenine,Citation206 it turned out that the physiological polymer polyP accelerates the healing rate of both experimental wounds in diabetic animalsCitation207 and chronic wounds in patients.Citation48,Citation49
In animal studies, polyP was applied into 8-mm wounds of diabetic mice and normal non-diabetic animals punched in the interscapular region down to the underlying fascia muscularis.Citation207 In both animal groups, the polymer was applied once. While in normal mice (untreated), the physiological regeneration proceeded by 31% re-epithelialization during a 7-day healing period, the polyP-treated animals showed a strong acceleration of the regeneration potential to 72% during 7 days. In diabetic controls, re-epithelialization was even less, only about 20% by day 7, while with polyP-NP, healing was greatly accelerated and was complete after 14 days.
These very promising results prompted our group to conduct off-label studies in patients. Depending on the depth of the wound, either more superficial or deeper to the hypodermis along the muscle or down even to the bone, the active substance has to be integrated into a hydrogel or alternatively into a collagen-based mat.Citation208 In the first human trial with chronic wounds, a hydrogel (with hydroxyethyl cellulose) containing 600 µg mL‒1 Na-polyP (to elicit immediate regenerative activity) together with 60 µg mL‒1 Ca-polyP-NP (acting as a depot) was prepared ().Citation49 The Na-polyP in the gel formulation undergoes direct coacervation after application onto the wound, taking up the Ca2+ from the wound bed (). The Ca-polyP-NP ingredient (depot form) must form a coacervate before becoming active ( and ). A clinical example is illustrated in . The 55-year-old female patient was suffering from a ventral ulcer on the tibia for 3 months. This patient had been treated with cortisone over a long period due to a number of pre-existing illnesses. Treatment of the wound with the hydrogel started on 10.11. (). After 8 weeks of treatment, the wound area was reduced by 87% (). The patient was released from the clinics 10 weeks later ().
Figure 13 Application of polyP for the treatment of chronic wounds. (A) Application of the polymer in (A-1) a hydrogel supplemented with Na-polyP and Ca-polyP-NP to cure more superficial wound. (A-2) Formation of a coacervate after contact with Ca2+ in the wound bed. (A-3) The presence of Ca-polyP-NP served as a depot for retarded polyP release. The chronic wound was treated with (A-4) the hydrogel. (A-5) After a 8 weeks of treatment, the wound dimension shrunk by 87%. (A-6) After a total of 18 weeks, the patient could go back home (B) For deeper wounds, (B-1) collagen-based mats were fabricated after compressing the collagen fibers. (B-2) Orientation of Ca-polyP-NP on the collagen bundles. (B-3) Every second day, a wetting solution composed of Na-polyP and Ca-polyP-NP was used and dropped onto the wound to keep it moist. There the Na-polyP underwent coacervation. Treatment success with the polyP-containing mats. (B-4) Initial state after debridement; (B-5) a strong reduction was observed after 6 weeks of treatment; (B-6) final state prior to patient discharge. (A) Adapted with permission from J Mat Sci Technol. Volume 135, Müller WEG, Schepler H, Neufurth M, et al. The physiological polyphosphate as a healing biomaterial for chronic wounds: Crucial roles of its antibacterial and unique metabolic energy supplying properties. 170–185. Copyright 2023. Creative Commons.Citation49 (B) Adapted with permission from Schepler H, Neufurth M, Wang SF, et al. Acceleration of chronic wound healing by bio-inorganic polyphosphate: In vitro studies and first clinical applications. Theranostics. 2022;12:18–34. © 2022 Ivyspring. Creative Commons.Citation48

Deep wounds can extend through the epidermis, dermis, hypodermis, and even to the cutaneous trunci muscle covering the skeletal muscle. These wounds require a wound dressing that elicits low antigenicity, suitable biocompatibility, and hemostatic properties to allow for cell proliferation and adhesion. One choice is collagen mats, which imitate the ECM, since collagen stabilizing the skin is present in the mats with an abundance of 60% to 70%. The collagen-based mats used here for application to deep wounds of patientsCitation48 were modified by compressing the collagen fibers in order to transfer the unstable collagen sheets to a strong wound-covering layer. For this purpose, bovine collagen samples were processed through a controlled pH shift series. The mats were then supplemented with 8 mg of Ca-polyP-NP per 1 mL collagen solution (10 mg mL‒1). The thickness of the mats was 1 to 1.3 mm (). The NP within the mats were oriented along the orientation of the collagen bundles (). Every second day, after application onto the wounds, the mats were moisturized with 1 mL of wetting solution composed of 300 μg g‒1 Na-polyP and 30 μg g‒1 (w/w final) Ca-polyP-NP to improve the healing process.Citation48 After moisturizing the mats, a coacervate developed from the Na-polyP component (). As with the hydrogel treatment, the results of the off-label studies with the wound mats have been very encouraging. An example is shown in . The 79-year-old male patient suffered from a chronic wound resulting from surgical resection of a squamous cell carcinoma. The treatment of the wound, with a dimension of ~20 cm2, started in April 2021 (). After debridement by surgery and ultrasound, the wound was covered with the polyP-collagen mat. After 6 weeks of treatment, the wound diameter had shrunken to 37% (), and after a further 12 weeks, the patient could be released home ().
Repair of Soft Tissue: Respiratory Epithelium
The epithelium of the respiratory tract (from the trachea to the bronchi) has an important barrier function against inhaled pathogens. The mucus gel layer produced by the goblet cells of the respiratory epithelium plays a crucial role in this defense mechanism by the entrapment of respiratory bacteria and viruses such as the coronavirus SARS-CoV-2. This epithelium is prone to damage not only by these pathogenic agents but also by environmental factors such as pollution by particulate matter. PolyP was found to induce the expression of mucins, the main protein component of the mucus.Citation139 In addition, it was found that polyP exhibits a strong antiviral activity against SARS-CoV-2.Citation140,Citation141 The finding that polyP elicits regenerative activity in the epithelial respiratory tract was not unexpected. The way it inhibits SARS-CoV-2 was surprising. Based on modeling studies, it has been demonstrated experimentally by in vitro studies and also supported by in vivo studies that polyP covers the tips of the coronavirus SARS-CoV-2 viral envelope spike proteins by binding to an unusually conserved cationic groove composed of basic amino acids (Arg, Lys, and His).Citation139,Citation140 Interestingly, the spacing of the basic amino acids on the spike protein perfectly matches the distances of 2.0 to 2.5 Å between the Pi units within the polyP molecule. In subsequent studies, a complete prevention of viral infection by polyP was experimentally proven. Due to this property, polyP can be classified as a component of the innate immune system and as a “smart” biomaterial (see ).Citation209
Application of Biosilica and Polyphosphate in Emerging Tissue Engineering Techniques
The two biomaterials, silica and polyP, can also contribute significantly to the field of tissue regeneration through the application of new tissue manufacturing techniques such as 3D printing technology. Among the existing 3D fabrication techniques such as selective laser sintering, stereolithography, fusion deposition modeling, digital light processing, and 3D bioprinting (for a review, see Ref.Citation210,Citation211), the latter in particular is one of the most advanced techniques. The development of 3D bioprinting is undergoing rapid development. This extrusion-based additive manufacturing technique is not only suitable for the creation of customized bone implants but also the fabrication of even large-sized tissue constructs and complex organs.
Nevertheless, this technique still has a number of limitations that await novel solutions. These include, for example, the fabrication of hollow structures such as blood vessels that would collapse as a result of gravitational forces during the layer-by-layer printing process. This makes, among others, the production of vascularized tissues difficult. One way to circumvent this problem is to use embedded bioprinting,Citation212 a gel-in-gel manufacturing technique in which the bio-ink is printed into a microgel or granular support bath. In the case of cortical bone implants, the printed material should have the required porosity and mechanical properties. Recapitulating the different mechanical properties and biological functions of the different parts of bone is a challenging task. Another requirement specifically for 3D cell printing (3D bioprinting) of tissues is the availability of a suitable bio-ink. This bio-ink, which usually consists of a hydrogel, certain other factors such as growth factors and the cells, must be able to protect the cells suspended in the hydrogel not only during the printing process but also afterwards, after curing (usually achieved by printing in a hardening solution containing Ca2+ ions), to maintain essential cell functions such as vitality, proliferation, migration, and differentiation abilities. Furthermore, the cells must be able to be printed in a density that corresponds to the high cell densities of native tissues and organs.
The introduction of the two biomaterials, silica and polyP, brought decisive progress, particularly with regard to the latter properties. Firstly, both amorphous biomaterials are regeneratively active themselves, they support cell vitality and promote cell proliferation/differentiation and do not require the addition of growth factors. The activity of such biodegradable growth factors/proteins is often difficult to control. Secondly, polyP in particular is able to provide the metabolic energy required for cell function, supporting cell migration and the synthesis, organization, and maintenance of the ECM structure. Cells can be printed at sufficiently high concentrations required for tissue regeneration/repair and formation of functional tissue constructs.
Taking advantage of these unique properties of silica and polyP/polyP-NP, the first bioprintable biosilica and polyP-containing hydrogels were developed. Embedding SaOS-2 cells in an alginate hydrogel resulted in a 3D printable material that showed increased HA formation and expression of bone morphogenetic protein-2 (BMP-2) and osteoprotegerin (OPG) genes.Citation213
The first cell-free, polyP-enriched bio-ink suitable for 3D printing consisted of Ca-polyP-NP embedded in a poly(ε-caprolactone) (PCL) matrix. The scaffolds printed with this bio-ink supported the ingrowth of SaOS-2 cells, accompanied by increased expression of the cell-attracting chemokine stromal cell-derived factor-1 (SDF-1).Citation214 The first cell-containing polyP-bio-ink consisted of Na-polyP, gelatin, and alginate complexed by Ca2+ ions. The cells (SaOS-2) embedded in this ink remain alive and grow efficiently in the 3D printed construct.Citation215 The best results were achieved – as described above – with a bio-ink based on N,O-CMC/alginate/gelatin containing both soluble polyP (Na-polyP) and Ca-polyP-NP. MSC contained in this matrix remained functionally active and showed increased cell migration and propensity to grow and differentiate into mineralizing osteoblasts.Citation152
As a further method, the electrospinning method has been used to fabricate advanced biosilica- or polyP-containing scaffold materials. Electrospun nanofiber mats prepared from poly(d,l-lactide) (PLA) and modified by enzymatically (silicatein)-formed biosilica were found to promote the growth and mineralization of SaOS-2 cells.Citation216 In addition, the synergistic effect of amorphous Ca-polyP-NP and retinol on cell growth has been exploited by manufacturing electrospun PLA fiber mats containing both components in view of their beneficial effects as antimicrobial and regeneratively active wound dressing materials.Citation217
Preferred Fields of Application
Both biomaterials, biosilica and polyP, have a wide range of potential applications. The most important ones identified are summarized in . Although there are some potential applications in the technical field, the preferred application of both materials is in tissue engineering and regenerative medicine. For the latter applications, the amorphous nature of the material is necessary. As shown in animal studies (rabbits; see ), silica encapsulated in PLGA microspheres alone or together with silicatein strongly stimulates bone regeneration after implantation in the femur compared to β-TCP controls.Citation83 As stated, a similar mineralization-promoting effect was also found with polyP, both with soluble polyPCitation189 and with polyP-NP, but also with polyP-stabilized ACCCitation68,Citation218 and polyP-stabilized ACP.Citation30,Citation69,Citation167 The effect of the polyP-NP is significantly influenced by the countercation of the polyanionic polyP. While Ca, Sr, and Mg are the preferred counterions in inducing bone regeneration,Citation135,Citation137,Citation142,Citation189 the Mg- and Ca-salts of the polymer are more suitable for cartilage regeneration.Citation136 Using the calvarial bone defect model, polyP-stabilized ACCCitation218 and polyP-stabilized ACPCitation30 encapsulated in PLGA and embedded in cranial defects of rats or rabbits significantly increased the formation of new bone mineral. β-TCP and crystalline Ca-phosphate, used as a controls, were much less active.
Table 2 Preferred Applications of Amorphous Biosilica and Amorphous polyP
Instead of encapsulation in microspheres, silica and polyP can also be administered after integration into a hydrogel-forming organic polymer matrix that can be hardened in the presence of divalent cations. In this way, biohybrid materials are created that combine the regenerative properties of the inorganic biomaterials/biopolymers with the advantageous mechanical and water-binding properties of the organic polymers. SaOS-2 cells embedded in an alginate-based hydrogel matrix containing biosilica formed by silicateinCitation213 or grown on 3D printed biosilica-alginate scaffoldsCitation219 showed increased growth, mineralization, and gene expression of BMP-2, collagen type 1, and OPG, but not of RANKL (receptor activator of NF-κB ligand). Increased osteogenic activity was also found with enzymatically formed biosilica on a chitosan-graft-polycaprolactone matrix.Citation234 Organic polymers used together with polyP mainly included negatively charged polysaccharides, as well as collagen and PVA. In the presence of divalent metal cations such as Ca2+ (used for bone implants) or Mg2+ ions (for cartilage), these materials can be cured by forming metal bridges to produce materials with suitable hardness and viscoelastic properties for potential use in bone and cartilage regeneration/repair. PVA is hardened by intermolecular cross-linking by freeze-thawing.Citation151 The addition of Ca2+ to the polyP-containing matrices can also lead to the in situ formation of Ca-polyP-NP,Citation150,Citation151 which upon contact with the body fluid are converted into a biologically active coacervate, as confirmed in animal studies.Citation151 Furthermore, due to their regenerative activity, biosilica and polyP can also be used in dentistry for tooth restoration/repair,Citation226 but also as dental sealants.Citation80,Citation227 Other identified potential applications of both polymeric biomaterials include their use in drug delivery, biologization of inert materials and protection against biofouling, including caries prevention ().
The most advanced application of polyP and polyP-NP in regenerative medicine is in the field of wound healing, specifically for the treatment of chronic wounds, where this polymer has already been successfully used in initial human studies, as described here. The material can be in the form of a wound gelCitation49 or incorporated into a collagen-based wound mat.Citation48,Citation217 In addition, the use of the remarkable antiviral activity of polyP,Citation125 especially against SARS-CoV-2,Citation139,Citation140 also proven in vivo, Citation141 in the form of a nasal spray is promising.
Conclusion and Outlook
At first glance, both the soft tissues, such as cartilage or fibrous connective tissue such as the dermal layer of the skin, and the bones or teeth, the hard tissue, are largely composed of non-cellular ECM structures that regulate almost all cellular functions. Especially during regeneration, these tissues are subject to continuous remodeling mediated by enzymes such as the catabolic-acting matrix-degrading enzymesCitation235 or the anabolic-acting ALP.Citation236 The dysregulation of the energy metabolism in cells of the ECM, such as fibroblasts (dermis layer of the skin), chondrocytes (cartilage), or bone cells, such as osteoblasts, osteocytes, and osteoclasts, disrupts the balance between various growth factors, proteoglycans, mineralizing cells, and enzymes.Citation237 Focusing on cells of the mineralized tissue, the central role of enzymes was first discovered in sponges, as summarized in this review. Furthermore, the connection of the metabolism of the ECM and the skeletal elements with bioenergetics was first disclosed in these evolutionary oldest animals. The mineralized elements in sponges are formed in an ATP-rich environment. A suitable biomaterial, in turn, must provide these two landmarks, enzymes and metabolic energy, to enable regeneration, morphogenesis, and repair of damaged tissue (). Here again, the sponges offered a solution, directing the bioinspired/biomimetic studies towards an inorganic biopolymer, polyP, which from then on became a distinguished biomaterial for engineering scaffolds and supplements for successful repair of damaged tissue with graded structural, mechanical, and compositional properties. The anionic polyP, in combination with cationic ions, acts as a structural and reinforcing biomaterial that tends to transform into a hydrogel-like coacervate in the presence of peptides.Citation143 In this water-rich phase, polyP undergoes enzymatic degradation and processing into ADP and ATP.Citation5
Figure 14 A suitable biomaterial is formed enzymatically and drives the expression of genes encoding for enzymes in the surrounding tissue. During regeneration, the structural proteins and the newly developed cells require metabolic energy for functional arrangement and positioning.

Also, wound healing is an energy-intensive process. Sponges are already provided with basal mechanisms involved in wound healing, including enzymes and signaling pathways that are found in higher animals and humans.Citation18 Sponges as sessile animals are sensitive to injury and tend to develop chronic wounds when repair is compromised. However, they can survive even in adverse conditions with low oxygen levels, possibly by using polyP accumulated in these animals.Citation238
Although biomineral formation in sponges and human bone shares some similarities in terms of enzyme involvement and energy requirements, it should be noted that both the chemical nature and the steps in which enzymes are involved are different. The sponge biosilica consists of covalently linked SiO4 tetrahedra that share an edge with each other (some edges of the tetrahedra remain unlinked as free silanol Si-OH groups) and is an amorphous material, while HA is a crystalline material composed of PO43‒, OH‒, CO32‒, and Ca2+ units linked together by ionic forces. In nature, in living organisms, both are composites, with silica as silica spicules consisting of multiple silica lamellae surrounding a central axial filament of silicatein and separated from each other by thin layers made of silicatein and some other proteins.Citation239 Bone, on the other hand, consists mainly of HA and collagen, which forms the matrix for the deposition of the HA crystals, but also of some other proteins involved in the regulation of the biomineralization process. The steps in which enzymes are involved in mineral deposition and the nature of the protein framework for biomineral deposition are also different. In the sponge spicules, the protein scaffold around which silica is deposited, itself carries the enzymatic, silica-forming activity, while in bone, the proteinaceous collagen scaffold is enzymatically inactive and enzymes (ALP) are only involved in providing the required energy and phosphate (released from polyP or some other sources, eg, ATP), as well as in the formation of the ACC precursor of ACP and crystalline HA (carbonic anhydrase). As a regeneratively active biomineral for human tissue regeneration/repair, silica consists only of the inorganic amorphous material.
Biosilica derived from sponges has a wide range of bioinspired applications, ranging from orthopedic implants, as summarized in this review, to dental sealantsCitation80 and light transmission systems.Citation31 The exploitation in human therapy is remarkable and based on the morphogenetic and regenerative activity of the material. Closely related to biosilica are bioactive glasses, also an amorphous material that was introduced in 1971.Citation240,Citation241 However, the chemical composition of biosilica is different from the CaO-SiO2-P2O5 composition of bioactive glasses prepared by sol–gel methods. Biosilica formed enzymatically by silicatein consists largely of a (water-rich) network of covalently linked SiO2 units, while 45S5 bioglass, for example, consists of 24.5 wt% CaO, 45.0 wt% SiO2, 6.0 wt% P2O5, and 24.5 wt% Na2O.Citation242,Citation243 As shown here, the morphogenetic activity of biosilica can be exploited in implants as demonstrated by the differential expression of various genes required for cell growth and differentiation in response to the biomineral.Citation80 In addition, from the natural biosilica, the blueprint, the fabrication of distinctive hierarchical porous structures could be learned, which even enabled the production of fibers with specific optical properties.Citation244
PolyP has a unique property to act both as a structural and degradable biomaterial for bone regeneration in orthopedics and simultaneously as a generator for extracellular metabolic energy. The application of polyP is even more advanced in the area of wound healing, especially the healing of chronic wounds. Healing of chronic wounds caused by metabolic diseases (diabetic foot ulcers), inadequate blood flow (venous leg ulcers), sustained pressure (bedsores), infections or cancer is still an unsolved medical challenge and a major cost driver in medical care. Initial applications in human patients with chronic wounds led to promising results, as described here. PolyP can be administered both in soluble form (as Na-polyP, immediately active) and as Ca-polyP-NP (depot form, prolonged effect), either as a wound creme or integrated into a collagen mat. Since the proof of concept for this biopolymer could be elaborated in human studies, its central position for physiological regeneration has been established.
Future tasks will also include using the two biomaterials silica and polyP to develop stimuli-responsive materials that are sensitive to specific signals from the tissue environment (pH, temperature, presence of certain biomolecules, etc.). Such materials may be endowed with the ability to decompose, swell or shrink, undergo sol–gel transition, or release certain drugs, growth factors, or other biomolecules in response to pH or other changes in the surrounding tissue.Citation245 In particular, pH changes (acidosis) that occur, for example, in the course of wound healing,Citation246 at tumor sitesCitation247 or during ischemiaCitation248 can be used to trigger the release of these molecules from the implanted or administered material. In recent years, a number of pH-dependent delivery systems have been developed, based, for example, on N,O-CMC,Citation249 alginate-dialdehyde-gentamicin/chitosan,Citation250 or poly(N-isopropylacrylamide-co-acrylic acid) hydrogels.Citation251 PolyP, as described in this review, is particularly suitable for the development of such delivery systems because it can transform from the nanoparticulate state (polyP-NP) to the coacervate state in a pH-dependent manner. Using bisphosphonate zoledronic acid, Ca-polyP-NP carrying this drug for potential treatment of tumors/metastases has already been synthesized.Citation138 A challenging task will be the development of silica- or polyP-based materials that carry multiple bioactive molecules or drugs, released at different times or on demand, and controlled by specific biochemical signals during multi-step repair processes such as the phases of wound healing (hemostasis, inflammation, proliferation, and maturation/remodeling) or the stages of bone regeneration (hematoma formation, inflammation, soft callus formation, hard callus formation, and remodeling). This could be realized by incorporating these biomolecules into various carrier systems such as core-shell particles, and nanoparticles (polyP-NP) with different counterions or nano/microspheres. Recently, a core-shell drug delivery system has been developed consisting of a polyP-NP core and a polyP-coacervate shell loaded with two different bioactive molecules (dexamethasone and ascorbic acid) that are released with different kinetics.Citation228 Such systems would also be of interest for the long-term treatment of osteoporosis caused by a reduced ratio of expression of the two cytokines OPG and RANKL, leading to increased osteoclastogenesis/bone resorption, or rheumatoid arthritis, which is associated with overexpression of inflammatory cytokines such as tumor necrosis factor α or interleukins 1, 6, or 7 causing bone destruction. Both polyP and biosilica have been shown to increase OPG expression,Citation9,Citation213 and polyP to decrease proinflammatory cytokine levels.Citation141 Finally, the beneficial effects of the regenerative activity of silica and polyP in other pathological conditions such as neurodegeneration need to be elucidated.
As summarized in this review, bioinspired/biomimetic thinking has the potential to reconcile the present-day demands of our comparatively very young species with its deficits. The human species (Homo sapiens) with an age of ~0.3 million years can benefit by learning from other metazoan taxa such as sponges (age 800 million years),Citation252 and also from the world of microorganisms, which can be traced back to over 3465 million years of the Earth’s history.Citation253 Keeping in mind that every organism on earth has a long, rich history driven by adaptation and survival of the fittest.Citation254 The biosilica and polyP materials discussed here have the groundbreaking potential for further scientific, biomedical, and technical innovations. Surely, these two materials will push ahead sustainable progress of our society in science, medicine, and technology.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
We thank our colleagues Prof. Dr W. Müller-Klieser and Prof. Dr S. Walenta (Institute of Pathophysiology; University Medical Center Mainz; Germany) for providing us with the cryosections through the sponge for the visualization of the ATP accumulation. W.E.G. M. is the holder of an ERC Advanced Investigator Grant (Grant No.: 268476). In addition, W.E.G. M. has obtained three ERC-PoC Grants (Si-Bone-PoC, Grant No.: 324564; MorphoVES-PoC, Grant No.: 662486; and ArthroDUR, Grant No.: 767234). In addition, this work was supported by Grants from the European Commission (Grant Nos.: 604036 and 311848), the International Human Frontier Science Program and the BiomaTiCS research initiative of the University Medical Center, Mainz. Further support came from the Grant of the BMBF (SKIN-ENERGY, Grant No.: 13GW0403A/B) and the Grant of the BMWi (Grant No.: ZF4294002AP9).
References
- Kenny NJ, Francis WR, Rivera-Vicéns RE, et al. Tracing animal genomic evolution with the chromosomal-level assembly of the freshwater sponge Ephydatia muelleri. Nat Commun. 2020;11(1):3676. doi:10.1038/s41467-020-17397-w
- Pertea M, Salzberg SL. Between a chicken and a grape: estimating the number of human genes. Genome Biol. 2010;11(5):206. doi:10.1186/gb-2010-11-5-206
- Williams D. The essential materials paradigms for regenerative medicine. JOM. 2011;63(4):51–55. doi:10.1007/s11837-011-0067-5
- Chiang B, Essick E, Ehringer W, et al. Enhancing skin wound healing by direct delivery of intracellular adenosine triphosphate. Am J Surg. 2007;193(2):213–218. doi:10.1016/j.amjsurg.2006.08.069
- Müller WEG, Schröder HC, Wang XH. Inorganic polyphosphates as storage for and generator of metabolic energy in the extracellular matrix. Chem Rev. 2019;119(24):12337–12374. doi:10.1021/acs.chemrev.9b00460
- Trackman PC. Diverse biological functions of extracellular collagen processing enzymes. J Cell Biochem. 2005;96(5):927–937. doi:10.1002/jcb.20605
- Wang XH, Schröder HC, Müller WEG. Enzyme-based biosilica and biocalcite: biomaterials for the future in regenerative medicine. Trends Biotechnol. 2014;32(9):441–447. doi:10.1016/j.tibtech.2014.05.004
- Wang Y, Li M, Li P, et al. Progress and applications of polyphosphate in bone and cartilage regeneration. Biomed Res Int. 2019;2019:5141204. doi:10.1155/2019/5141204
- Schröder HC, Wang XH, Neufurth M, Wang SF, Tan R, Müller WEG. Inorganic polymeric materials for injured tissue repair: biocatalytic formation and exploitation. Biomedicines. 2022;10(3):658. doi:10.3390/biomedicines10030658
- Vallesius F. Controversiarum medicarum et philosophicarum. W.C. Hanoviae: Marnium; 1606.
- Trembley A. Mémoires pour servir à l’histoire d’un genre de polypes d’eau douce à bra sen forme de cornes. Verbeek: Leide; 1744.
- Galtsoff PS. The amoeboid movement of dissociated sponge cells. Biol Bull. 1923;45(3):153–161. doi:10.2307/1536625
- Müller WEG. The stem cell concept in sponges (Porifera): metazoan traits. Semin Cell Dev Biol. 2006;17(4):481–491. doi:10.1016/j.semcdb.2006.05.006
- Morgan TH. Regeneration. New York/London: Macmillan; 1901.
- Bely AE, Nyberg KG. Evolution of animal regeneration: re-emergence of a field. Trends Ecol Evol. 2010;25(3):161–170. doi:10.1016/j.tree.2009.08.005
- Ereskovsky A, Borisenko IE, Bolshakov FV, Lavrov AI. Whole-body regeneration in sponges: diversity, fine mechanisms, and future prospects. Genes. 2021;12(4):506. doi:10.3390/genes12040506
- Ricci L, Srivastava M. Wound-induced cell proliferation during animal regeneration. Wiley Interdiscip Rev Dev Biol. 2018;7(5):e321. doi:10.1002/wdev.321
- Wu YC, Franzenburg S, Ribes M, Pita L. Wounding response in Porifera (sponges) activates ancestral signaling cascades involved in animal healing, regeneration, and cancer. Sci Rep. 2022;12(1):1307. doi:10.1038/s41598-022-05230-x
- Cary GA, Wolff A, Zueva O, Pattinato J, Hinman VF. Analysis of sea star larval regeneration reveals conserved processes of whole-body regeneration across the metazoa. BMC Biol. 2019;17(1):16. doi:10.1186/s12915-019-0633-9
- Rybicka-Jasińska K, Derr JB, Vullev VI. What defines biomimetic and bioinspired science and engineering?. Pure Appl Chem. 2021;93(11):1275–1292. doi:10.1515/pac-2021-0323
- Gough A, Soto-Gutierrez A, Vernetti L, Ebrahimkhani MR, Stern AM, Taylor DL. Human biomimetic liver microphysiology systems in drug development and precision medicine. Nat Rev Gastroenterol Hepatol. 2021;18(4):252–268. doi:10.1038/s41575-020-00386-1
- Benyus JM. Biomimicry: Innovation Inspired by Nature. Perennial, New York: HaperCollins Pub; 2002.
- Furth JJ, Cohen SS. Inhibition of mammalian DNA polymerase by the 5’-triphosphate of 1-β-D-arabinofuranosylcytosine and the 5’-triphosphate of 9-β-D-arabinofuranoxyladenine. Cancer Res. 1968;28(10):2061–2067.
- Schabel FM. The antiviral activity of 9-β-D-arabinofuranosyladenine (ARA-A). Chemotherapy. 1968;13(6):321–338. doi:10.1159/000220567
- Müller WEG, Zahn RK, Bittlingmaier K, Falke D. Inhibition of herpesvirus DNA-synthesis by 9-ß-D-arabinofuranosyladenosine in vitro and in vivo. Ann New York Acad Sci. 1977;284(1):34–48. doi:10.1111/j.1749-6632.1977.tb21935.x
- Seack J, Pancer Z, Müller IM, Müller WEG. Molecular cloning and primary structure of a Rhesus (Rh)-like protein from the marine sponge Geodia cydonium. Immunogenetics. 1997;46(6):493–498. doi:10.1007/s002510050310
- Müller WEG, Blumbach B, Müller IM. Evolution of the innate and adaptive immune systems: relationships between potential immune molecules in the lowest metazoan phylum [Porifera] and those in vertebrates. Transplantation. 1999;68(9):1215–1227. doi:10.1097/00007890-199911150-00001
- Krasko A, Lorenz B, Batel R, Schröder HC, Müller IM, Müller WEG. Expression of silicatein and collagen genes in the marine sponge Suberites domuncula is controlled by silicate and myotrophin. Eur J Biochem. 2000;267(15):4878–4887. doi:10.1046/j.1432-1327.2000.01547.x
- Müller WEG, Wang XH, Grebenjuk V, et al. Common genetic denominators for Ca++-based skeleton in metazoan: role of osteoclast stimulating factor and carbonic anhydrase in a calcareous sponge. PLoS One. 2012;7(4):e34617. doi:10.1371/journal.pone.0034617
- Müller WEG, Ackermann M, Al-Nawas B, et al. Amplified morphogenetic and bone forming activity of amorphous versus crystalline calcium phosphate/polyphosphate. Acta Biomater. 2020;118:233–247. doi:10.1016/j.actbio.2020.10.023
- Müller WEG, Wendt K, Geppert C, Wiens M, Reiber A, Schröder HC. Novel photoreception system in sponges? Unique transmission properties of the stalk spicules from the hexactinellid Hyalonema sieboldi. Biosens Bioelectron. 2006;21(7):1149–1155. doi:10.1016/j.bios.2005.04.017
- Liu S, Yu JM, Gan YC, et al. Biomimetic natural biomaterials for tissue engineering and regenerative medicine: new biosynthesis methods, recent advances, and emerging applications. Mil Med Res. 2023;10(1):16. doi:10.1186/s40779-023-00448-w
- Omidian H, Wilson RL, Babanejad N. Bioinspired polymers: transformative applications in biomedicine and regenerative medicine. Life. 2023;13:1673.
- Weiner S, Dove PM. An overview of biomineralization processes and the problem of the vital effect. Rev Mineral Geochem. 2003;54(1):1–29. doi:10.2113/0540001
- Westbroek P. Biological metal accumulation and biomineralization in a geological perspective. In: Westbroek P, de Jong EW, editors. Biomineralization and Biological Metal Accumulation. Dordrecht, Holland: D. Reidel Publishing Co; 1983:1–11.
- Krasko A, Gamulin V, Seack J, Steffen R, Schröder HC, Müller WEG. Cathepsin, a major protease of the marine sponge Geodia cydonium: purification of the enzyme and molecular cloning of cDNA. Molec Marine Biol Biotechnol. 1997;6:296–307.
- Shimizu K, Cha J, Stucky GD, Morse DE. Silicatein α: cathepsin L-like protein in sponge biosilica. Proc Natl Acad Sci USA. 1998;95(11):6234–6238. doi:10.1073/pnas.95.11.6234
- Mărgineanu DG. Equilibrium and non-equilibrium approaches in biomembrane thermodynamics. Arch Internat Physiol Biochim. 1987;95:381–422.
- Seibel MJ. Biochemical markers of bone turnover. Part I Biochemistry and variability. Clin Biochem Rev. 2005;26(4):97–122.
- Harris H. The human alkaline phosphatases: what we know and what we don’t know. Clin Chim Acta. 1990;186(2):133–150. doi:10.1016/0009-8981(90)90031-M
- Schini M, Vilaca T, Gossiel F, Salam S, Eastell R. Bone turnover markers: basic biology to clinical applications. Endocr Rev. 2023;44:417–473.
- Müller WEG, Schröder HC, Schloßmacher U, Grebenjuk VA, Ushijima H, Wang XH. Induction of carbonic anhydrase in SaOS-2 cells, exposed to bicarbonate and consequences for calcium phosphate crystal formation. Biomaterials. 2013;34(34):8671–8680. doi:10.1016/j.biomaterials.2013.07.096
- Andrilli LHS, Sebinelli HG, Favarin BZ, et al. NPP1 and TNAP hydrolyze ATP synergistically during biomineralization. Purinergic Signal. 2023;19:353–366.
- Morrison MS, Turin L, King BF, Burnstock G, Arnett TR. ATP is a potent stimulator of the activation and formation of rodent osteoclasts. J Physiol. 1998;511(2):495–500. doi:10.1111/j.1469-7793.1998.495bh.x
- de Paula FJ, Rosen CJ. Bone remodeling and energy metabolism: new perspectives. Bone Res. 2013;1:72–84.
- Motyl KJ, Guntur AR, Carvalho AL, Rosen CJ. Energy metabolism of bone. Toxicol Pathol. 2017;45(7):887–893. doi:10.1177/0192623317737065
- Müller WEG, Wang SF, Ackermann M, et al. Biologization of allogeneic bone grafts with polyphosphate: a route to a biomimetic periosteum. Adv Funct Mater. 2019;29(44):1905220. doi:10.1002/adfm.201905220
- Schepler H, Neufurth M, Wang SF, .et al. Acceleration of chronic wound healing by bio-inorganic polyphosphate: in vitro studies and first clinical applications. Theranostics. 2022;12(1):18–34. doi:10.7150/thno.67148
- Müller WEG, Schepler H, Neufurth M, et al. The physiological polyphosphate as a healing biomaterial for chronic wounds: crucial roles of its antibacterial and unique metabolic energy supplying properties. J Mat Sci Technol. 2023;135:170–185.
- Müller WEG, Wang SF, Neufurth M, Kokkinopoulou M, Feng Q, Schröder HC. Polyphosphate as a donor of high-energy phosphate for the synthesis of ADP and ATP. J Cell Sci. 2017;130:2747–2756.
- Müller WEG, Ackermann M, Tolba E, et al. Role of ATP during the initiation of microvascularization. Acceleration of an autocrine sensing mechanism facilitating chemotaxis by inorganic polyphosphate. Biochem J. 2018;475(20):3255–3273. doi:10.1042/BCJ20180535
- Michaëlsson E, Malmström V, Reis S, Engström A, Burkhardt H, Holmdahl R. T cell recognition of carbohydrates on type II collagen. J Exp Med. 1994;180:745–749.
- Müller WEG, Wiens M, Adell T, Gamulin V, Schröder HC, Müller IM. Bauplan of Urmetazoa: basis for genetic complexity of Metazoa. Intern Rev Cytol. 2004;235:53–92.
- Uriz MJ, Turon X, Becerro MA, Agell G. Siliceous spicules and skeleton frameworks in sponges: origin, diversity, ultrastructural patterns, and biological functions. Microsc Res Tech. 2003;62:279–299.
- Müller WEG, Blumbach B, Wagner-Hülsmann C, Lessel U. Galectins in the phylogenetically oldest metazoa, the sponges [Porifera]. Trends Glycosci Glycotechnol. 1997;9(45):123–130. doi:10.4052/tigg.9.123
- Weaver JC, Aizenberg J, Fantner GE, et al. Hierarchical assembly of the siliceous skeletal lattice of the hexactinellid sponge Euplectella aspergillum. J Struct Biol. 2007;158(1):93–106. doi:10.1016/j.jsb.2006.10.027
- Wang XH, Schröder HC, Müller WEG. Giant siliceous spicules from the deep-sea glass sponge Monorhaphis chuni: morphology, biochemistry and molecular biology. Int Rev Cell Mol Biol. 2009;273:69–115.
- Zhou Y, Shimizu K, Cha JN, Stucky GD, Morse DE. Efficient catalysis of polysiloxane synthesis by silicatein α requires specific hydroxy and imidazole functionalities. Angew Chem Int Ed Engl. 1999;38:779–782.
- Schröder HC, Borejko A, Krasko A, Reiber A, Schwertner H, Müller WEG. Mineralization of SaOS-2 cells on enzymatically (Silicatein) modified bioactive osteoblast-stimulating surfaces. J Biomed Mat Res Part B Appl Biomater. 2005;75B:387–392.
- Schloßmacher U, Wiens M, Schröder HC, Wang XH, Jochum KP, Müller WEG. Silintaphin-1: interaction with silicatein during structure-guiding biosilica formation. FEBS J. 2011;278(7):1145–1155. doi:10.1111/j.1742-4658.2011.08040.x
- Müller WEG, Boreiko A, Wang XH, et al. Silicateins, the major biosilica forming enzymes present in demosponges: protein analysis and phylogenetic relationship. Gene. 2007;395(1–2):62–71. doi:10.1016/j.gene.2007.02.014
- Tabatabaei Dakhili SY, Caslin SA, Faponle AS, Quayle P, de Visser SP, Wong LS. Recombinant silicateins as model biocatalysts in organosiloxane chemistry. Proc Natl Acad Sci USA. 2017;114:E5285–E5291.
- Müller WEG, Wiens M, Batel R, et al. Establishment of a primary cell culture from a sponge: primmorphs from Suberites domuncula. Mar Ecol Progr Ser. 1999;178:205–219. doi:10.3354/meps178205
- Walenta S, Dötsch J, Mueller-Klieser W. ATP concentrations in multicellular tumor spheroids assessed by single photon imaging and quantitative bioluminescence. Eur J Cell Biol. 1990;52(2):389–393.
- Boury-Esnault N, Rützler K. Thesaurus of sponge morphology. Smithson Contrib Zool. 1997;596(596):1–55. doi:10.5479/si.00810282.596
- Weiner S, Mahamid J, Politi Y, Ma Y, Addadi L. Overview of the amorphous precursor phase strategy in biomineralization. Frontiers Mat Sci China. 2009;3:104–108.
- Jackson DJ, Macis L, Reitner J, Degnan BM, Wörheide G. Sponge paleogenomics reveals an ancient role for carbonic anhydrase in skeletogenesis. Science. 2007;316(5833):1893–1895. doi:10.1126/science.1141560
- Tolba E, Müller WEG, El-Hady BMA, et al. High biocompatibility and improved osteogenic potential of amorphous calcium carbonate/vaterite. J Mat Chem B. 2016;4(3):376–386. doi:10.1039/C5TB02228B
- Müller WEG, Neufurth M, Ushijima H, et al. Molecular and biochemical approach for understanding the transition of amorphous to crystalline calcium phosphate deposits in human teeth. Dental Mater. 2022;38:2014–2029.
- Crundwell FK. On the mechanism of the dissolution of quartz and silica in aqueous solutions. ACS Omega. 2017;2(3):1116–1127. doi:10.1021/acsomega.7b00019
- Schröder HC, Perović-Ottstadt S, Rothenberger M, et al. Silica transport in the demosponge Suberites domuncula: fluorescence emission analysis using the PDMPO probe and cloning of a potential transporter. Biochem J. 2004;381(3):665–673. doi:10.1042/BJ20040463
- Müller WEG, Schloßmacher U, Wang XH, et al. Poly(silicate)-metabolizing silicatein in siliceous spicules and silicasomes of demosponges comprises dual enzymatic activities (silica-polymerase and silica-esterase). FEBS J. 2008;275:362–370.
- Wang XH, Schröder HC, Brandt D, et al. Sponge biosilica formation involves syneresis following polycondensation in vivo. Chembiochem. 2011;12(15):2316–2324. doi:10.1002/cbic.201100345
- Belton DJ, Deschaume O, Patwardhan SV, Perry CC. A solution study of silica condensation and speciation with relevance to in vitro investigations of biosilicification. J Phys Chem B. 2010;114(31):9947–9955. doi:10.1021/jp101347q
- Belton DJ, Deschaume O, Perry CC. An overview of the fundamentals of the chemistry of silica with relevance to biosilicification and technological advances. FEBS J. 2012;279(10):1710–1720. doi:10.1111/j.1742-4658.2012.08531.x
- Cha JN, Shimizu K, Zhou Y, et al. Silicatein filaments and subunits from a marine sponge direct the polymerization of silica and silicones in vitro. Proc Natl Acad Sci USA. 1999;96:361–365.
- Müller WEG, Belikov SI, Tremel W, et al. Siliceous spicules in marine demosponges (example Suberites domuncula). Micron. 2006;37(2):107–120. doi:10.1016/j.micron.2005.09.003
- Schröder HC, Wang XH, Manfrin A, et al. Silicatein: acquisition of structure-guiding and structure-forming properties during maturation from the pro-silicatein to the silicatein form. J Biol Chem. 2012;287(26):22196–22205. doi:10.1074/jbc.M112.351486
- Stöber W, Fink A, Bohn E. Controlled growth of monodisperse silica spheres in the micron size range. J Colloid Interface Sci. 1968;26(1):62–69. doi:10.1016/0021-9797(68)90272-5
- Müller WEG, Boreiko A, Wang XH, et al. Morphogenetic activity of silica and bio-silica on the expression of genes, controlling biomineralization using SaOS-2 cells. Calcif Tissue Int. 2007;81(5):382–393. doi:10.1007/s00223-007-9075-4
- Görlich S, Pawolski D, Zlotnikov I, Kröger N. Control of biosilica morphology and mechanical performance by the conserved diatom gene Silicanin-1. Commun Biol. 2019;2(1):245. doi:10.1038/s42003-019-0436-0
- Ege D, Zheng K, Boccaccini AR. Borate Bioactive Glasses (BBG): bone regeneration, wound healing applications, and future directions. ACS Appl Bio Mater. 2022;5(8):3608–3622. doi:10.1021/acsabm.2c00384
- Wang SF, Wang XH, Draenert FG, et al. Bioactive and biodegradable silica biomaterial for bone regeneration. Bone. 2014;67:292–304. doi:10.1016/j.bone.2014.07.025
- Carlisle EM. Silicon: an essential element for the chick. Science. 1972;178(4061):619–621. doi:10.1126/science.178.4061.619
- Burton EJ, Scholey DV, Belton DJ, Bedford MR, Perry CC. Efficacy and stability of a novel silica supplement for improving bone development in broilers. Br Poult Sci. 2020;61(6):719–724. doi:10.1080/00071668.2020.1799328
- Wiens M, Wang XH, Schloßmacher U, et al. Osteogenic potential of bio-silica on human osteoblast-like (SaOS-2) cells. Calcif Tissue Int. 2010;87(6):513–524. doi:10.1007/s00223-010-9408-6
- Pettersson LF, Kingham PJ, Wiberg M, Kelk P. In vitro osteogenic differentiation of human mesenchymal stem cells from jawbone compared with dental tissue. Tissue Eng Regen Med. 2017;14:763–774.
- Pautke C, Vogt S, Kreutzer K, et al. Characterization of eight different tetracyclines: advances in fluorescence bone labeling. J Anat. 2010;217(1):76–82. doi:10.1111/j.1469-7580.2010.01237.x
- Hench LL. Bioceramics. J Am Ceram Soc. 1998;81(7):1705–1728. doi:10.1111/j.1151-2916.1998.tb02540.x
- Wang Y, Jiang S, Pan H, Tang R. Less is more: silicate in the crystallization of hydroxyapatite in simulated body fluids. CrystEngComm. 2016;18(3):379–383. doi:10.1039/C5CE01861G
- Kellermeier M, Melero-García E, Glaab F, et al. Stabilization of amorphous calcium carbonate in inorganic silica-rich environments. J Am Chem Soc. 2010;132(50):17859–17866. doi:10.1021/ja106959p
- Sahai N. Modeling apatite nucleation in the human body and in the geochemical environment. Am J Sci. 2000;305(6–8):661–672. doi:10.2475/ajs.305.6-8.661
- Sahai N, Tossell JA. Molecular orbital study of apatite (Ca5(PO4)3OH) nucleation at silica bioceramic surfaces. J Phys Chem B. 2000;104(18):4321–4322. doi:10.1021/jp9935889
- Salih E, Flückiger R. Complete topographical distribution of both the in vivo and in vitro phosphorylation sites of bone sialoprotein and their biological significance. J Biol Chem. 2004;279(19):19808–19815. doi:10.1074/jbc.M310299200
- Wysokowski M, Jesionowski T, Ehrlich H. Biosilica as a source for inspiration in biological materials science. Amer Mineral. 2018;103:665–691.
- Lorenz B, Batel R, Bachinski N, Müller WEG, Schröder HC. Purification and characterization of two exopolyphosphatases from the marine sponge Tethya lyncurium. Biochim Biophys Acta. 1995;1245(1):17–28. doi:10.1016/0304-4165(95)00067-L
- Kulaev IS. Biochemistry of inorganic polyphosphates. Rev Physiol Biochem Pharmacol. 1975;73:131–158.
- Kumble KD, Kornberg A. Inorganic polyphosphate in mammalian cells and tissues. J Biol Chem. 1995;270(11):5818–5822. doi:10.1074/jbc.270.11.5818
- Achbergerová L, Nahálka J. Polyphosphate--an ancient energy source and active metabolic regulator. Microb Cell Fact. 2011;10(1):63. doi:10.1186/1475-2859-10-63
- Desfougères Y, Saiardi A, Azevedo C. Inorganic polyphosphate in mammals: where’s Wally?. Biochem Soc Trans. 2020;48(1):95–101. doi:10.1042/BST20190328
- Docampo R, Huang G. Acidocalcisomes of eukaryotes. Curr Opin Cell Biol. 2016;41:66–72. doi:10.1016/j.ceb.2016.04.007
- Lander N, Cordeiro C, Huang G, Docampo R. Polyphosphate and acidocalcisomes. Biochem Soc Trans. 2016;44(1):1–6. doi:10.1042/BST20150193
- Babes V. Beobachtungen über die metachromatischen Körperchen, Sporenbildung, Verzweigung, Kolben- und Kapselbildung pathogener Bakterien. Zentralbl Bakteriol Parasitenkd Infektionskr Hyg. 1895;20:412–420.
- Meyer A. Orientierende Untersuchungen über Verbreitung, Morphologie, und Chemie des Volutins. Bot Zeit. 1904;62:113–152.
- Ruiz FA, Lea CR, Oldfield E, Docampo R. Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes. J Biol Chem. 2004;279(43):44250–44257. doi:10.1074/jbc.M406261200
- Langen P, Hucho F. Karl Lohmann and the discovery of ATP. Angew Chem Int Ed Engl. 2008;47(10):1824–1827. doi:10.1002/anie.200702929
- Langen P, Liss E, Lohmann K. Art, Bildung und Umsatz der Polyphosphate der Hefe [Formation and turnover of polyphosphates in yeast]. In: Jacquinot P, editor. Acides Ribonucléiques Et Polyphosphates: Structure, Synthèse Et Fonctions. CNRS No 105. Paris: Centre National de la Recherche Scientifique; 1962:604–614.
- Lohmann K. Über das Vorkommen, und den Umsatz von Pyrophosphat in Zellen. I. Mitteilung: nachweis und Isolierung des Pyrophosphates. Biochem Z. 1928;202:466–493.
- Bowlin MQ, Gray MJ. Inorganic polyphosphate in host and microbe biology. Trends Microbiol. 2021;29(11):1013–1023. doi:10.1016/j.tim.2021.02.002
- Hooley P, Whitehead MP, Brown MR. Eukaryote polyphosphate kinases: is the ‘Kornberg’ complex ubiquitous?. Trends Biochem Sci. 2008;33(12):577–582. doi:10.1016/j.tibs.2008.09.007
- Pavlov E, Zakharian E, Bladen C, et al. A large, voltage-dependent channel, isolated from mitochondria by water-free chloroform extraction. Biophys J. 2005;88(4):2614–2625. doi:10.1529/biophysj.104.057281
- Pavlov E, Aschar-Sobbi R, Campanella M, Turner RJ, Gomez-Garcia MR, Abramov AY. Inorganic polyphosphate and energy metabolism in mammalian cells. J Biol Chem. 2010;285(13):9420–9428. doi:10.1074/jbc.M109.013011
- Abramov AY, Fraley C, Diao CT, et al. Targeted polyphosphatase expression alters mitochondrial metabolism and inhibits calcium-dependent cell death. Proc Natl Acad Sci USA. 2007;104(46):18091–18096. doi:10.1073/pnas.0708959104
- Budavari S. The Merck Index: An Encyclopedia of Chemicals, Drugs, and Biologicals. 11th ed. Rahway, NJ: Merck & Co; 1989.
- Soga N, Kinosita K, Yoshida M, Suzuki T. Efficient ATP synthesis by thermophilic Bacillus Fo F1-ATP synthase. FEBS J. 2011;278(15):2647–2654. doi:10.1111/j.1742-4658.2011.08191.x
- Hothorn M, Neumann H, Lenherr ED, et al. Catalytic core of a membrane-associated eukaryotic polyphosphate polymerase. Science. 2009;324(5926):513–516. doi:10.1126/science.1168120
- Guan Z, Chen J, Liu R, et al. The cytoplasmic synthesis and coupled membrane translocation of eukaryotic polyphosphate by signal-activated VTC complex. Nat Commun. 2023;14(1):718. doi:10.1038/s41467-023-36466-4
- Verhoef JJ, Barendrecht AD, Nickel KF, et al. Polyphosphate nanoparticles on the platelet surface trigger contact system activation. Blood. 2017;129(12):1707–1717. doi:10.1182/blood-2016-08-734988
- Morrissey JH, Smith SA. Polyphosphate as modulator of hemostasis, thrombosis, and inflammation. J Thromb Haemost. 2015;13(Suppl 1):S92–S97. doi:10.1111/jth.12896
- Polasek J. Platelet lysosomal acid phosphatase enzyme activity as a marker of platelet procoagulant activity. Blood Transfus. 2009;7(2):155–156. doi:10.2450/2008.0053-08
- Buxton IL, Kaiser RA, Oxhorn BC, Cheek DJ. Evidence supporting the nucleotide axis hypothesis: ATP release and metabolism by coronary endothelium. Am J Physiol Heart Circ Physiol. 2001;281(4):H1657–H1666. doi:10.1152/ajpheart.2001.281.4.H1657
- Weitz JI, Fredenburgh JC. Platelet polyphosphate: the long and the short of it. Blood. 2017;129(12):1574–1575. doi:10.1182/blood-2017-01-761593
- Sharda A, Flaumenhaft R. The life cycle of platelet granules. F1000Res. 2018;7:236. doi:10.12688/f1000research.13283.1
- Tyagi T, Jain K, Gu SX, et al. A guide to molecular and functional investigations of platelets to bridge basic and clinical sciences. Nat Cardiovasc Res. 2022;1(3):223–237. doi:10.1038/s44161-022-00021-z
- Lorenz B, Leuck J, Köhl D, Müller WEG, Schröder HC. Anti-HIV-1 activity of inorganic polyphosphates. J Acquir Immune Defic Syndr Hum Retrovirol. 1997;14(2):110–118. doi:10.1097/00042560-199702010-00003
- Lorenz B, Schröder HC. Mammalian intestinal alkaline phosphatase acts as highly active exopolyphosphatase. Biochim Biophys Acta. 2001;1547(2):254–261. doi:10.1016/S0167-4838(01)00193-5
- Ho-Tin-Noé B, Boulaftali Y, Camerer E. Platelets and vascular integrity: how platelets prevent bleeding in inflammation. Blood. 2018;131(3):277–288. doi:10.1182/blood-2017-06-742676
- Harrison P, Alsousou J, Andia I; Subcommittee on Platelet Physiology. The use of platelets in regenerative medicine and proposal for a new classification system: guidance from the SSC of the ISTH. J Thromb Haemost. 2018;16(9):1895–1900. doi:10.1111/jth.14223
- Bizzozero J. Ueber einen neuen Formbestandtheil des Blutes und dessen Rolle bei der Thrombose und der Blutgerinnung. Virchow’s Arch Pathol Anat Physiol Klin Med. 1882;90(2):261–332. doi:10.1007/BF01931360
- Roewe J, Stavrides G, Strueve M, et al. Bacterial polyphosphates interfere with the innate host defense to infection. Nat Commun. 2020;11(1):4035. doi:10.1038/s41467-020-17639-x
- Ruggeri ZM, Mendolicchio GL. Adhesion mechanisms in platelet function. Circ Res. 2007;100(12):1673–1685. doi:10.1161/01.RES.0000267878.97021.ab
- Jackson LE, Kariuki BM, Smith ME, Barralet JE, Wright AJ. Synthesis and structure of a calcium polyphosphate with an unique criss-cross arrangement of helical phosphate chains. Chem Mater. 2005;17(18):4642–4646. doi:10.1021/cm050984x
- Shiba T, Nishimura D, Kawazoe Y, et al. Modulation of mitogenic activity of fibroblast growth factors by inorganic polyphosphate. J Biol Chem. 2003;278(29):26788–26792. doi:10.1074/jbc.M303468200
- Lorenz B, Marmé S, Müller WEG, Unger K, Schröder HC. Preparation and use of polyphosphate-modified zirconia for purification of nucleic acids and proteins. Anal Biochem. 1994;16(1):118–126. doi:10.1006/abio.1994.1015
- Müller WEG, Tolba E, Schröder HC, et al. A new polyphosphate calcium material with morphogenetic activity. Mater Lett. 2015;148:166. doi:10.1016/j.matlet.2015.02.070
- Wang XH, Ackermann M, Tolba E, et al. Artificial cartilage bio-matrix formed of hyaluronic acid and Mg2+-polyphosphate. Eur Cell Mater. 2016;32:271–283. doi:10.22203/eCM.v032a18
- Müller WEG, Tolba E, Ackermann M, et al. Fabrication of amorphous strontium polyphosphate microparticles that induce mineralization of bone cells in vitro and in vivo. Acta Biomater. 2017;50:89–101. doi:10.1016/j.actbio.2016.12.045
- Müller WEG, Neufurth M, Wang SF, et al. Amorphous, smart, and bioinspired polyphosphate nano/microparticles: a biomaterial for regeneration and repair of osteo-articular impairments in-situ. Int J Mol Sci. 2018;19(2):427. doi:10.3390/ijms19020427
- Müller WEG, Schröder HC, Neufurth M, Wang XH. An unexpected biomaterial against SARS-CoV-2: bio-polyphosphate blocks binding of the viral spike to the cell receptor. Mater Today. 2021;51:504–524. doi:10.1016/j.mattod.2021.07.029
- Neufurth M, Wang XH, Tolba E, et al. The inorganic polymer, polyphosphate, blocks binding of SARS-CoV-2 spike protein to ACE2 receptor at physiological concentrations. Biochem Pharmacol. 2020;182:114215. doi:10.1016/j.bcp.2020.114215
- Ferrucci V, Kong DY, Asadzadeh F, et al. Long-chain polyphosphates impair SARS-CoV-2 infection and replication. Sci Signal. 2021;14(690):eabe5040. doi:10.1126/scisignal.abe5040
- Wang XH, Schröder HC, Müller WEG. Amorphous polyphosphate, a smart bioinspired nano-/bio-material for bone and cartilage regeneration: towards a new paradigm in tissue engineering. J Mat Chem B. 2018;6(16):2385–2412. doi:10.1039/C8TB00241J
- Müller WEG, Wang SF, Tolba E, et al. Transformation of amorphous polyphosphate nanoparticles into coacervate complexes: an approach for the encapsulation of mesenchymal stem cells. Small. 2018;14(27):1801170. doi:10.1002/smll.201801170
- Müller WEG, Neufurth M, Lieberwirth I, Wang SF, Schröder HC, Wang XH. Functional importance of coacervation to convert calcium polyphosphate nanoparticles into the physiologically active state. Mater Today Bio. 2022;16:100404. doi:10.1016/j.mtbio.2022.100404
- Fernandes-Cunha GM, McKinlay CJ, Vargas JR, Jessen HJ, Waymouth RM, Wender PA. Delivery of inorganic polyphosphate into cells using amphipathic oligocarbonate transporters. ACS Cent Sci. 2018;4(10):1394–1402. doi:10.1021/acscentsci.8b00470
- Barbeck M, Alkildani S, Jung O. Editorial of the Special Issue: “Soft and Hard Tissue Regeneration”. Biomedicines. 2022;10(2):356. doi:10.3390/biomedicines10020356
- Lehn JM. Supramolecular chemistry: receptors, catalysts, and carriers. Science. 1985;227(4689):849–856. doi:10.1126/science.227.4689.849
- Fasting C, Schalley CA, Weber M, et al. Multivalency as a chemical organization and action principle. Angew Chem Int Ed Engl. 2012;51(42):10472–10498. doi:10.1002/anie.201201114
- Müller WEG, Neufurth M, Wang S, Tolba E, Schröder HC, Wang XH. Morphogenetically active scaffold for osteochondral repair (polyphosphate/alginate/N,O-carboxymethyl chitosan). Eur Cell Mater. 2016;31:174–190. doi:10.22203/eCM.v031a12
- Müller WEG, Neufurth M, Ackermann M, et al. Fabrication of a new physiological macroporous hybrid biomaterial/bioscaffold material based on polyphosphate and collagen by freeze-extraction. J Mat Chem B. 2017;5(21):3823–3835. doi:10.1039/C7TB00306D
- Tolba E, Wang XH, Ackermann M, et al. In-situ polyphosphate nanoparticle formation in hybrid poly(vinyl alcohol)/karaya gum-hydrogels: a porous scaffold inducing infiltration of mesenchymal stem cells. Adv Sci. 2018;2018:1801452.
- Neufurth M, Wang SF, Schröder HC, Al-Nawas B, Wang XH, Müller WEG. 3D bioprinting of tissue units with mesenchymal stem cells, retaining their proliferative and differentiating potential, in polyphosphate-containing bio-ink. Biofabrication. 2022;14(1):015016. doi:10.1088/1758-5090/ac3f29
- Krieg E, Bastings MM, Besenius P, Rybtchinski B. Supramolecular polymers in aqueous media. Chem Rev. 2016;116(4):2414–2477. doi:10.1021/acs.chemrev.5b00369
- Samperi M, Pérez-García L, Amabilino DB. Quantification of energy of activation to supramolecular nanofibre formation reveals enthalpic and entropic effects and morphological consequence. Chem Sci. 2019;10(44):10256–10266. doi:10.1039/C9SC03280K
- Harris JR, Lewis RJ. The collagen type I segment long spacing (SLS) and fibrillar forms: formation by ATP and sulphonated diazo dyes. Micron. 2016;86:36–47. doi:10.1016/j.micron.2016.04.008
- Trautmann A. Extracellular ATP in the immune system: more than just a “danger signal”. Sci Signal. 2009;2(56):pe6. doi:10.1126/scisignal.256pe6
- Huang H, Zhang X, Li S, et al. Physiological levels of ATP negatively regulate proteasome function. Cell Res. 2010;20(12):1372–1385. doi:10.1038/cr.2010.123
- Schenk U, Westendorf AM, Radaelli E, et al. Purinergic control of T cell activation by ATP released through pannexin-1 hemichannels. Sci Signal. 2008;1(39):ra6. doi:10.1126/scisignal.1160583
- Etulain J. Platelets in wound healing and regenerative medicine. Platelets. 2018;29(6):556–568. doi:10.1080/09537104.2018.1430357
- Swieringa F, Spronk HMH, Heemskerk JWM, van der Meijden PEJ. Integrating platelet and coagulation activation in fibrin clot formation. Res Pract Thromb Haemost. 2018;2(3):450–460. doi:10.1002/rth2.12107
- Zuk PA, Zhu M, Mizuno H, et al. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng. 2001;7(2):211–228. doi:10.1089/107632701300062859
- Yang J, Zhang YS, Yue K, Khademhosseini A. Cell-laden hydrogels for osteochondral and cartilage tissue engineering. Acta Biomater. 2017;57:1–25. doi:10.1016/j.actbio.2017.01.036
- Laverdet B, Micallef L, Lebreton C, et al. Use of mesenchymal stem cells for cutaneous repair and skin substitute elaboration. Pathol Biol. 2014;62(2):108–117. doi:10.1016/j.patbio.2014.01.002
- Leyhausen G, Lorenz B, Zhu H, et al. Inorganic polyphosphate in human osteoblast-like cells. J Bone Mineral Res. 1998;13(5):803–812. doi:10.1359/jbmr.1998.13.5.803
- Schröder HC, Kurz L, Müller WEG, Lorenz B. Polyphosphate in bone. Biochemistry. 2000;65(3):296–303.
- Morimoto D, Tomita T, Kuroda S, et al. Inorganic polyphosphate differentiates human mesenchymal stem cells into osteoblastic cells. J Bone Miner Metab. 2010;28(4):418–423. doi:10.1007/s00774-010-0157-4
- Müller WEG, Tolba E, Schröder HC, Muñoz-Espí R, Diehl-Seifert B, Wang XH. Amorphous polyphosphate-hydroxyapatite: a morphogenetically active substrate for bone-related SaOS-2 cells in vitro. Acta Biomater. 2016;31:358–367. doi:10.1016/j.actbio.2015.11.060
- Bolander J, Ji W, Geris L, et al. The combined mechanism of bone morphogenetic protein- and calcium phosphate-induced skeletal tissue formation by human periosteum derived cells. Eur Cell Mater. 2016;31:11–25. doi:10.22203/eCM.v031a02
- Morrissey JH, Choi SH, Smith SA. Polyphosphate: an ancient molecule that links platelets, coagulation, and inflammation. Blood. 2012;119(25):5972–5979. doi:10.1182/blood-2012-03-306605
- Hill T, Morales M. On “High energy phosphate bonds” of biochemical interest 1. J Am Chem Soc. 1951;73(4):1656–1660. doi:10.1021/ja01148a072
- Lipman F. Metabolic generation and utilization of phosphate bond energy. In: Nord FF, Werkman CH, editors. Advances in Enzymology and Related Subjects of Biochemistry. Vol. 1. New York: Interscience Publishers Inc; 1941:99–162.
- Hessle L, Johnson KA, Anderson HC, et al. Tissue-nonspecific alkaline phosphatase and plasma cell membrane glycoprotein-1 are central antagonistic regulators of bone mineralization. Proc Natl Acad Sci USA. 2002;99(14):9445–9449. doi:10.1073/pnas.142063399
- Klepinin A, Zhang S, Klepinina L, et al. Adenylate kinase and metabolic signaling in cancer cells. Front Oncol. 2020;10:660. doi:10.3389/fonc.2020.00660
- Ishikawa HO, Xu A, Ogura E, Manning G, Irvine KD, Giniger E. The Raine syndrome protein FAM20C is a Golgi kinase that phosphorylates bio-mineralization proteins. PLoS One. 2012;7(8):e42988. doi:10.1371/journal.pone.0042988
- Tagliabracci VS, Xiao J, Dixon JE. Phosphorylation of substrates destined for secretion by the Fam20 kinases. Biochem Soc Trans. 2013;41(4):1061–1065. doi:10.1042/BST20130059
- Tagliabracci VS, Wiley SE, Guo X, et al. A single kinase generates the majority of the secreted phosphoproteome. Cell. 2015;161(7):1619–1632. doi:10.1016/j.cell.2015.05.028
- Raine J, Winter RM, Davey A, Tucker SM. Unknown syndrome: microcephaly, hypoplastic nose, exophthalmos, gum hyperplasia, cleft palate, low set ears, and osteosclerosis. J Med Genet. 1989;26(12):786–788. doi:10.1136/jmg.26.12.786
- Lowenstam HA. Minerals formed by organisms. Science. 1981;211(4487):1126–1131. doi:10.1126/science.7008198
- Ruben JA, Bennett AA. The evolution of bone. Evolution. 1987;41(6):1187–1197. doi:10.2307/2409087
- Mann S, Parker SB, Ross MD, Skarnulis AJ, Williams RJ. The ultrastructure of the calcium carbonate balance organs of the inner ear: an ultra-high resolution electron microscopy study. Proc R Soc Lond B Biol Sci. 1983;218:415–424.
- Setiawati R, Rahardjo P. Bone development and growth. In: Yang HS, editor. Osteogenesis and Bone Regeneration. Intechopen; 2019. doi:10.5772/intechopen.82452
- Müller WEG, Neufurth M, Huang J, et al. Nonenzymatic transformation of amorphous CaCO3 into calcium phosphate mineral after exposure to sodium phosphate in vitro: implications for in vivo hydroxyapatite bone formation. ChemBioChem. 2015;16(9):1323–1332. doi:10.1002/cbic.201500057
- Tadier S, Rokidi S, Rey C, Combes C, Koutsoukos PG. Crystal growth of aragonite in the presence of phosphate. J Crystal Growth. 2017;458:44–52. doi:10.1016/j.jcrysgro.2016.10.046
- Boskey AL, Guidon P, Doty SB, Stiner D, Leboy P, Binderman I. The mechanism of β-glycerophosphate action in mineralizing chick limb-bud mesenchymal cell cultures. J Bone Miner Res. 1996;11(11):1694–1702. doi:10.1002/jbmr.5650111113
- Sharif PS, Abdollahi M. The role of platelets in bone remodeling. Inflamm Allergy Drug Targets. 2010;9(5):393–399. doi:10.2174/187152810793938044
- Müller WEG, Wang XH, Diehl-Seifert B, et al. Inorganic polymeric phosphate/polyphosphate as an inducer of alkaline phosphatase and a modulator of intracellular Ca2+ level in osteoblasts (SaOS-2 cells) in vitro. Acta Biomater. 2011;7(6):2661–2671. doi:10.1016/j.actbio.2011.03.007
- Khersonsky O, Tawfik DS. Enzyme promiscuity: a mechanistic and evolutionary perspective. Annu Rev Biochem. 2010;79(1):471–505. doi:10.1146/annurev-biochem-030409-143718
- Lotsari A, Rajasekharan AK, Halvarsson M, Andersson M. Transformation of amorphous calcium phosphate to bone-like apatite. Nat Commun. 2018;9(1):4170. doi:10.1038/s41467-018-06570-x
- Wang XH, Wang SF, He F, et al. Polyphosphate as a bioactive and biodegradable implant material: induction of bone regeneration in rats. Adv Engin Mat. 2016;18(8):1406–1417. doi:10.1002/adem.201600057
- Meldrum FC, Cölfen H. Controlling mineral morphologies and structures in biological and synthetic systems. Chem Rev. 2008;108(11):4332–4432. doi:10.1021/cr8002856
- Mayr-Wohlfart U, Waltenberger J, Hausser H, et al. Vascular endothelial growth factor stimulates chemotactic migration of primary human osteoblasts. Bone. 2002;30(3):472–477. doi:10.1016/S8756-3282(01)00690-1
- Martineau I, Lacoste E, Gagnon G. Effects of calcium and thrombin on growth factor release from platelet concentrates: kinetics and regulation of endothelial cell proliferation. Biomaterials. 2004;25(18):4489–4502. doi:10.1016/j.biomaterials.2003.11.013
- Marenzana M, Arnett TR. The key role of the blood supply to bone. Bone Res. 2013;1(3):203–215. doi:10.4248/BR201303001
- Bilgen F, Ural A, Bekerecioglu M. Platelet-rich fibrin: an effective chronic wound healing accelerator. J Tissue Viability. 2021;30(4):616–620. doi:10.1016/j.jtv.2021.04.009
- Schultz GS, Chin GA, Moldawer L, Diegelmann RF. Principles of wound healing. In: Fitridge R, Thompson M, editors. Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists, Chapter 23. Adelaide (AU): University of Adelaide Press; 2011. Available from: https://www.ncbi.nlm.nih.gov/books/NBK534261. Accessed January 31, 2024.
- Rodriguez-Diaz A, Toyama Y, Abravanel DL, et al. Actomyosin purse strings: renewable resources that make morphogenesis robust and resilient. HFSP J. 2008;2(4):220–237. doi:10.2976/1.2955565
- Jacinto A, Martinez-Arias A, Martin P. Mechanisms of epithelial fusion and repair. Nat Cell Biol. 2001;3(5):E117–E123. doi:10.1038/35074643
- Santamaría R, González-álvarez M, Delgado R, Esteban S, Arroyo AG. Remodeling of the microvasculature: may the blood flow be with you. Front Physiol. 2020;11:586852. doi:10.3389/fphys.2020.586852
- Schwiebert EM, Zsembery A. Extracellular ATP as a signaling molecule for epithelial cells. Biochim Biophys Acta. 2003;1615(1–2):7–32. doi:10.1016/S0005-2736(03)00210-4
- De Bock K, Georgiadou M, Schoors S, et al. Role of PFKFB3-driven glycolysis in vessel sprouting. Cell. 2013;154(3):651–663. doi:10.1016/j.cell.2013.06.037
- Leung SWS, Shi Y. The glycolytic process in endothelial cells and its implications. Acta Pharmacol Sin. 2022;43(2):251–259. doi:10.1038/s41401-021-00647-y
- Spampinato SF, Caruso GI, De Pasquale R, Sortino MA, Merlo S. The treatment of impaired wound healing in diabetes: looking among old drugs. Pharmaceuticals. 2020;13(4):60. doi:10.3390/ph13040060
- Sreekumar R, Halvatsiotis P, Schimke JC, Nair KS. Gene expression profile in skeletal muscle of type 2 diabetes and the effect of insulin treatment. Diabetes. 2002;51(6):1913–1920. doi:10.2337/diabetes.51.6.1913
- Patti ME, Butte AJ, Crunkhorn S, et al. Coordinated reduction of genes of oxidative metabolism in humans with insulin resistance and diabetes: potential role of PGC1 and NRF1. Proc Natl Acad Sci USA. 2003;100(14):8466–8471. doi:10.1073/pnas.1032913100
- Sarojini H, Bajorek A, Wan R, et al. Enhanced skin incisional wound healing with intracellular ATP delivery via macrophage proliferation and direct collagen production. Front Pharmacol. 2021;12:594586. doi:10.3389/fphar.2021.594586
- Yang JY, Chen CC, Chang SC, et al. ENERGI-F703 gel, as a new topical treatment for diabetic foot and leg ulcers: a multicenter, randomized, double-blind, Phase II trial. EClinicalMedicine. 2022;51:101497. doi:10.1016/j.eclinm.2022.101497
- Müller WEG, Relkovic D, Ackermann M, et al. Enhancement of wound healing in normal and diabetic mice by topical application of amorphous polyphosphate. Polymers. 2017;9(12):300. doi:10.3390/polym9070300
- Stan D, Tanase C, Avram M, et al. Wound healing applications of creams and “smart” hydrogels. Exp Dermatol. 2021;30(9):1218–1232. doi:10.1111/exd.14396
- Rosso F, Marino G, Giordano A, Barbarisi M, Parmeggiani D, Barbarisi A. Smart materials as scaffolds for tissue engineering. J Cell Physiol. 2005;203(3):465–470. doi:10.1002/jcp.20270
- Budharaju H, Suresh S, Sekar MP, et al. Ceramic materials for 3D printing of biomimetic bone scaffolds – current state-of-The-art & future perspectives. Mater Des. 2023;231:112064. doi:10.1016/j.matdes.2023.112064
- Ananth KP, Jayram ND. A comprehensive review of 3D printing techniques for biomaterial-based scaffold fabrication in bone tissue engineering. Ann 3D Print Med. 2024;13:100141. doi:10.1016/j.stlm.2023.100141
- Budharaju H, Sundaramurthi D, Sethuraman S. Embedded 3D bioprinting – an emerging strategy to fabricate biomimetic & large vascularized tissue constructs. Bioact Mater. 2024;32:356–384. doi:10.1016/j.bioactmat.2023.10.012
- Müller WEG, Schröder HC, Feng QL, Schlossmacher U, Link T, Wang XH. Development of a morphogenetically active scaffold for three-dimensional growth of bone cells: biosilica-alginate hydrogel for SaOS-2 cell cultivation. J Tissue Eng Regen Med. 2015;9(11):E39–E50. doi:10.1002/term.1745
- Neufurth M, Wang XH, Wang SF, et al. 3D printing of hybrid biomaterials for bone tissue engineering: calcium-polyphosphate microparticles encapsulated by polycaprolactone. Acta Biomater. 2017;64:377–388. doi:10.1016/j.actbio.2017.09.031
- Neufurth M, Wang XH, Schröder HC, et al. Engineering a morphogenetically active hydrogel for bioprinting of bioartificial tissue derived from human osteoblast-like SaOS-2 cells. Biomaterials. 2014;35(31):8810–8819. doi:10.1016/j.biomaterials.2014.07.002
- Müller WEG, Tolba E, Schröder HC, Diehl-Seifert B, Link T, Wang XH. Biosilica-loaded poly(ϵ-caprolactone) nanofibers mats provide a morphogenetically active surface scaffold for the growth and mineralization of the osteoclast-related SaOS-2 cells. Biotechnol J. 2014;9(10):1312–1321. doi:10.1002/biot.201400277
- Müller WEG, Tolba E, Dorweiler B, Schröder HC, Diehl-Seifert B, Wang XH. Electrospun bioactive mats enriched with Ca-polyphosphate/retinol nanospheres as potential wound dressing. Biochem Biophys Rep. 2015;3:150–160. doi:10.1016/j.bbrep.2015.08.007
- Wang XH, Ackermann M, Wang SF, et al. Amorphous polyphosphate/amorphous calcium carbonate implant material with enhanced bone healing efficacy in a critical-size defect in rats. Biomed Mater. 2016;11(3):035005. doi:10.1088/1748-6041/11/3/035005
- Schloßmacher U, Schröder HC, Wang XH, et al. Alginate/silica composite hydrogel as a potential morphogenetically active scaffold for three-dimensional tissue engineering. RSC Adv. 2013;3(28):11185–11194. doi:10.1039/c3ra23341c
- Braun S, Rappoport S, Zusman R, Avnir D, Ottolenghi M. Biochemically active sol-gel glasses: the trapping of enzymes. Mater Lett. 1990;10(1–2):1–5. doi:10.1016/0167-577X(90)90002-4
- Nassif N, Roux C, Coradin T, Rager MN, Bouvet OMM, Livage J. A sol–gel matrix to preserve the viability of encapsulated bacteria. J Mat Chem. 2003;13(2):203–208. doi:10.1039/b210167j
- Müller WEG, Engel S, Wang XH, et al. Bioencapsulation of living bacteria (Escherichia coli) with poly(silicate) after transformation with silicatein-α gene. Biomaterials. 2008;29(7):771–779. doi:10.1016/j.biomaterials.2007.10.038
- Müller WEG, Wang XH, Proksch P, et al. Principles of biofouling protection in marine sponges: a model for the design of novel biomimetic and bio-inspired coatings in the marine environment?. Mar Biotechnol. 2013;15(4):375–398. doi:10.1007/s10126-013-9497-0
- Müller WEG, Tolba E, Schröder HC, et al. A new printable and durable N,O-carboxymethyl chitosan-Ca2+ -polyphosphate complex with morphogenetic activity. J Mat Chem B. 2015;3(8):1722–1730. doi:10.1039/C4TB01586J
- Wang SF, Neufurth M, Schepler H, et al. Acceleration of wound healing through amorphous calcium carbonate, stabilized with high-energy polyphosphate. Pharmaceutics. 2023;15(2):494. doi:10.3390/pharmaceutics15020494
- Ackermann M, Tolba E, Neufurth M, et al. Biomimetic transformation of polyphosphate microparticles during restoration of damaged teeth. Dent Mater. 2019;35(2):244–256. doi:10.1016/j.dental.2018.11.014
- Müller WEG, Neufurth M, Tolba E, et al. A biomimetic approach to ameliorate dental hypersensitivity by amorphous polyphosphate microparticles. Dent Mater. 2016;32(6):775–783. doi:10.1016/j.dental.2016.03.027
- Müller WEG, Tolba E, Wang SF, et al. Nanoparticle-directed and ionically forced polyphosphate coacervation: a versatile and reversible core-shell system for drug delivery. Sci Rep. 2020;10(1):17147. doi:10.1038/s41598-020-73100-5
- Neufurth M, Wang XH, Tolba E, et al. Modular small diameter vascular grafts with bioactive functionalities. PLoS One. 2015;10(7):e0133632. doi:10.1371/journal.pone.0133632
- Ackermann M, Wang XH, Wang SF, et al. Collagen-inducing biologization of prosthetic material for hernia repair: polypropylene meshes coated with polyP/collagen. J Biomed Mater Res B. 2018;106(6):2109–2121. doi:10.1002/jbm.b.34016
- Smith SA, Morrissey JH. Polyphosphate as a general procoagulant agent. J Thromb Haemost. 2008;6(10):1750–1756. doi:10.1111/j.1538-7836.2008.03104.x
- Smith SA, Mutch NJ, Baskar D, Rohloff P, Docampo R, Morrissey JH. Polyphosphate modulates blood coagulation and fibrinolysis. Proc Natl Acad Sci USA. 2006;103(4):903–908. doi:10.1073/pnas.0507195103
- Moon JH, Park JH, Lee JY. Antibacterial Action of Polyphosphate on Porphyromonas gingivalis. Antimicrob Agents Chemother. 2011;55(2):806–812. doi:10.1128/AAC.01014-10
- Wiens M, Elkhooly TA, Schröder HC, Mohamed TH, Müller WEG. Characterization and osteogenic activity of a silicatein/biosilica-coated chitosan-graft-polycaprolactone. Acta Biomater. 2014;10(10):4456–4464. doi:10.1016/j.actbio.2014.06.036
- Phang JM, Liu W, Zabirnyk O. Proline metabolism and microenvironmental stress. Annu Rev Nutr. 2010;30(1):441–463. doi:10.1146/annurev.nutr.012809.104638
- Zhou Z, Fan Y, Jiang Y, et al. Mineralized enzyme-based biomaterials with superior bioactivities for bone regeneration. ACS Appl Mater Interfaces. 2022;14(32):36315–36330. doi:10.1021/acsami.2c05794
- Yang J, Ueharu H, Mishina Y. Energy metabolism: a newly emerging target of BMP signaling in bone homeostasis. Bone. 2020;138:115467. doi:10.1016/j.bone.2020.115467
- Imsiecke G, Münkner J, Lorenz B, Bachinski N, Müller WEG, Schröder HC. Inorganic polyphosphates in the developing freshwater sponge Ephydatia muelleri: effect of stress by polluted waters. Environ Toxicol Chem. 1996;15(8):1329–1334. doi:10.1002/etc.5620150811
- Müller WEG, Wang XH, Sinha B, Wiens M, Schröder HC, Jochum KP. NanoSIMS: insights into the organization of the proteinaceous scaffold within hexactinellid sponge spicules. ChemBioChem. 2010;11(8):1077–1082. doi:10.1002/cbic.201000078
- Hench LL, Splinter RJ, Allen WC, Greenlee TK. Bonding mechanisms at the interface of ceramic prosthetic materials. J Biomed Mater Res Symp. 1971;5:117–141.
- Zhu H, Zheng K, Boccaccini AR. Multi-functional silica-based mesoporous materials for simultaneous delivery of biologically active ions and therapeutic biomolecules. Acta Biomater. 2021;129:1–17. doi:10.1016/j.actbio.2021.05.007
- Hench LL. The story of Bioglass. J Mater Sci Mater Med. 2006;17(11):967–978. doi:10.1007/s10856-006-0432-z
- Rahaman MN, Day DE, Bal BS, et al. Bioactive glass in tissue engineering. Acta Biomater. 2011;7(6):2355–2373. doi:10.1016/j.actbio.2011.03.016
- Moffa M, Camposeo A, Fasano V, et al. Biomineral amorphous lasers through light-scattering surfaces assembled by electrospun fiber templates. Laser Photonics Rev. 2018;12(1):1700224. doi:10.1002/lpor.201700224
- Szwed-Georgiou A, Płociński P, Kupikowska-Stobba B, et al. Bioactive materials for bone regeneration: biomolecules and delivery systems. ACS Biomater Sci Eng. 2023;9(9):5222–5254. doi:10.1021/acsbiomaterials.3c00609
- Schneider LA, Korber A, Grabbe S, Dissemond J. Influence of pH on wound-healing: a new perspective for wound-therapy?. Arch Dermatol Res. 2007;298(9):413–420. doi:10.1007/s00403-006-0713-x
- Raghunand N, Gatenby RA, Gillies RJ. Microenvironmental and cellular consequences of altered blood flow in tumours. Br J Radiol. 2003;76(Spec No 1):S11–S22. doi:10.1259/bjr/12913493
- Yan GX, Kléber AG. Changes in extracellular and intracellular pH in ischemic rabbit papillary muscle. Circ Res. 1992;71(2):460–470. doi:10.1161/01.RES.71.2.460
- Gan Q, Zhu J, Yuan Y, Liu H, Zhu Y, Liu C. A proton-responsive ensemble using mesocellular foam supports capped with N,O-carboxymethyl chitosan for controlled release of bioactive proteins. J Mater Chem B. 2015;3(11):2281–2285. doi:10.1039/C5TB00219B
- Tao B, Deng Y, Song L, et al. BMP2-loaded titania nanotubes coating with pH-responsive multilayers for bacterial infections inhibition and osteogenic activity improvement. Colloids Surf B Biointerfaces. 2019;177:242–252. doi:10.1016/j.colsurfb.2019.02.014
- Banerjee I, Mishra D, Das T, Maiti TK. Wound pH-responsive sustained release of therapeutics from a poly(NIPAAm-co-AAc) hydrogel. J Biomater Sci Polym Ed. 2012;23(1–4):111–132. doi:10.1163/092050610X545049
- Schäcke H, Müller IM, Müller WEG. Tyrosine kinase from the marine sponge Geodia cydonium: the oldest member belonging to the receptor tyrosine kinase class II family. In: Müller WEG, editor. Use of Aquatic Invertebrates as Tools for Monitoring of Environmental Hazards. Stuttgart, New York: Gustav Fischer Verlag; 1994:201–211.
- Schopf JW. Microfossils of the early Archean apex chert: new evidence of the antiquity of life. Science. 1993;260(5108):640–646. doi:10.1126/science.260.5108.640
- Müller WEG, Brümmer F, Batel R, Müller IM, Schröder HC. Molecular biodiversity. Case study: porifera (sponges). Naturwissenschaften. 2003;90(3):103–120. doi:10.1007/s00114-003-0407-6