
Abstract
Purpose
Asthma is a chronic heterogeneous respiratory disease resulting from a complex interplay between genetic variations and environmental exposures. There are sex disparities in the prevalence and severity of asthma in males and females. Asthma prevalence is higher in males during childhood but increases in females in adulthood. The mechanisms underlying these sex differences are not well understood; nevertheless, genetic variations, hormonal changes, and environmental influences are thought to play important roles. This study aimed to identify sex-specific genetic variants associated with asthma using CLSA genomic and questionnaire data.
Methods
First, we conducted a genome-wide SNP-by-sex interaction analysis on 23,323 individuals, examining 416,562 single nucleotide polymorphisms (SNPs) after quality control, followed by sex-stratified survey logistic regression of SNPs with interaction p-value less than 10¯5.
Results
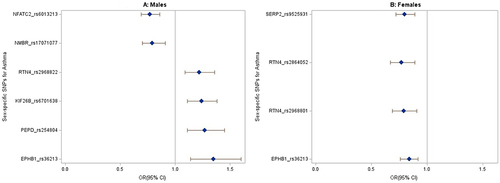
Out of the 49 SNPs with interaction p-value less than 10−5, a sex-stratified survey logistic regression showed that five male-specific SNPs (rs6701638, rs17071077, rs254804, rs6013213, and rs2968822) in/near KIF26B, NMBR, PEPD, RTN4, and NFATC2 loci, and three female-specific SNPs (rs2968801, rs2864052, and rs9525931) in/near RTN4, and SERP2 loci were significantly associated with asthma after Bonferroni correction. An SNP (rs36213) in the EPHB1 gene was significantly associated with an increased risk of asthma in males [OR=1.35, 95% CI (1.14, 1.60)] but with a reduced risk of asthma in females [OR=0.84, 95% CI (0.76, 0.92)] after Bonferroni correction.
Conclusion
We discovered novel sex-specific genetic markers in/near the KIF26B, RTN4, EPHB1, NMBR, SERP2, PEPD, and NFATC2 genes that could potentially shed light on the sex differences in asthma susceptibility in males and females. Future mechanistic studies are required to understand better the underlying sex-related pathways of the identified loci in asthma development.
Introduction
Asthma is a chronic inflammatory respiratory disease that affects both children and adults. It is characterized by reversible airflow obstruction, airway remodeling, hyperresponsiveness, shortness of breath, wheezing, and mucus production. In 2019, there were more than 262 million asthma cases globally, and the age-standardized point prevalence was highest in the high-income North American region.Citation1 In Canada, between 2011 and 2012, an estimated 3.8 million people one year and older lived with asthma.Citation2 Asthma prevalence and incidence differ in childhood and adulthood, and sex differences in prevalence, incidence, susceptibility, and severity have long been recognized.Citation3–5 During childhood, the prevalence of asthma is higher in males than in females, while in puberty and adulthood, females are more likely to develop asthma than males.Citation6
Sexual dimorphism has been reported to influence the severity, pathogenesis, pathophysiology, progression, and susceptibility of various human diseases and treatments, including asthma.Citation7–10 Women with asthma tend to have higher mortality and morbidity,Citation11 greater perception of symptom bothersomeness, and poorer quality of life than men with asthma.Citation12,Citation13 Studies using antigen-challenged mouse asthma models have shown that female mice had greater levels of antigen-specific serum IgE, eosinophils, and Th2 cytokine than male mice in the lung tissue and bronchoalveolar lavage fluid.Citation14,Citation15 Furthermore, airway hyperresponsiveness has been reported to be greater in male mice than in female mice at six weeks of age and reduced at twelve weeks of age.Citation16
Although the underlying biological mechanism of respiratory diseases and sex differences in risk are not fully understood, it is commonly believed that anatomical differences, genetic predisposition, hormonal changes, environmental exposures, and gene and environment interactions play an important role.Citation4,Citation17–19
Genetic predisposition plays a vital role in asthma pathogenesis. More than a hundred independent genetic loci have been associated with asthma.Citation20,Citation21 Studies have shown that several gene polymorphisms and gene expressions associated with asthma are sex-specific.Citation22–25 For instance, a large genome-wide association study (GWAS) using EVE Asthma Genetics Consortium discovered six sex-specific asthma risk loci, with two male-specific SNPs in/near IRF and RAB11FIP2 gene and four female-specific SNPs in/near RAP1GAP2, C6orf118, ERBB4, and AK057517.Citation25 Other sex-specific polymorphisms in many genes have been identified, eg, a male-specific SNP in the LCORL gene was significantly associated with childhood asthma,Citation22 and a female-specific polymorphism in the COX-2−765 gene was significantly associated with bronchial asthma.Citation26 Most GWAS of asthma generally include limited information on social and environmental factors and do not have enough sample size to investigate sex-specific genetic effects; this study aims to identify the sex-specific genetic polymorphisms significantly associated with asthma in middle-aged and older Canadian adults using the Canadian Longitudinal Study on Aging (CLSA) data (Baseline Comprehensive and Genomic datasets).
Methods and Materials
Study Population
This study included 26,622 individuals with complete genotyping information from the Canadian Longitudinal Study for Aging (CLSA) comprehensive cohort.Citation27 The CLSA comprehensive cohort comprises over 30,000 middle-aged and older Canadian adults between the ages of 45 and 85 years. Baseline data on a wide range of variables, including socio-demographic and socio-economic factors, information on lifestyle and health behaviors, physical measurements including height, weight, pulmonary function test, and biospecimens (blood and urine), were obtained through in-person home interviews and visits to data collection sites (DCS).Citation28,Citation29
Definition of Asthma
A positive response to the following question from the CLSA questionnaire determined the presence of self-reported physician-diagnosed asthma, “Has a doctor ever told you that you have asthma?” However, we excluded the following participants from this study: participants with missing responses, participants who self-reported a physician diagnosis of COPD, and participants who self-reported a physician diagnosis of asthma and COPD.
Covariates
The following potential confounders were considered: age and principal components of genetic ancestry were included as continuous variables. Categorical variables included were age groups (45 to 54 years, 55 to 64 years, 65 to 74 years, and over 75 years), biological sex (male vs female), smoking status (current, never, and former smokers), marital status (single/never married, married/common-law relationship, widowed/divorced/separated), education level (less than post-secondary education, post-secondary but not university education, and university education/others), total personal income and total household income (Less than $20,000, $20,000 to less than $50,000, $50,000 to less than $100,000, $100,000 or more), province of recruitment (Prairies, British Columbia, Eastern provinces, Ontario and Quebec), retirement status (Retired completely vs retired/partly retired), homeownership (owned vs rented/others), and urban/rural dwelling (urban core vs rural/not urban core).
Genotyping, Sample, and Marker Quality Control
Genotyping of 794,409 genetic markers was undertaken using Affymetrix UK Biobank Axiom array.Citation27 Detailed sample and markers quality control have been described previously.Citation27 Samples and markers that failed the quality control requirements were excluded. Samples with inconsistent sex information (discrepancy between reported sex and sex determined from genetic data) and high rates of genotype missingness (> 5%). SNPs in sex chromosomes, SNPs with low genotype call rates (< 99%), SNPs with minor allele frequency < 1% (MAF < 0.01), and SNPs deviating from the Hardy Weinberg equilibrium threshold of 1e-10 were excluded. Using PLINK’s Indep-pairwise command (Indep-pairwise 50, 5, 0.5), we generated a subset of SNPs in approximate linkage equilibrium. Post-quality control, we had 2,799 asthma cases, 20,524 controls, and 416,562 SNPs for genome-wide SNP-by-sex interaction analysis.
Statistical Analysis
Descriptive statistics, mean (SE), and frequency (%) for continuous and categorical variables were presented to describe the study population. Characteristics between male and female participants were compared using chi-square tests and Student’s t-test. Trimmed inflation and analytic weights provided in the CLSA data (CLSA Sample Weights Version 1.2) were used for descriptive and regression analyses.
In order to identify sex-specific SNPs associated with asthma, first, we performed a multivariate logistic regression using PLINK 1.90b6.2Citation30,Citation31 to identify significant interaction terms between a sex variable, and each of the SNPs in the regression analysis of asthma after controlling for age, sex, smoking status, and the first four principal components of genetic ancestry. Second, we performed a sex-stratified analysis for those SNPs with an SNP-by-sex interaction p-value less than 10−5 using survey-specific logistic regression (Proc Surveylogistic) in SAS 9.4 version. This approach allowed us to include sampling weights, complex survey design variables and adjust for potential confounders.
A purposeful model selection method was used to identify potential confounders for inclusion in the final model of the sex-stratified analysis. The covariates with a p-value ≤ 0.20 from the univariate analysis were entered into an interim multivariate model. The least significant covariate was then removed one at a time until only covariates with significant p-values (p ≤ 0.05) and clinically important factors remained in the model. The following covariates were included in the final model of the sex-stratified analysis for asthma: age, smoking status, province of recruitment, marital status, total personal income, and retirement status. The “sex variable” was included as a domain factor in the sex-stratified multivariate survey logistic regression to examine one SNP at a time after controlling for the first four principal components, age, smoking status, province of recruitment, marital status, total personal income, and retirement status.
Three inheritance models (dominant, recessive, and additive) for each variant were evaluated. However, only the inheritance model with the smallest AIC value (the best-fitted model) was selected and presented. Bonferroni correction was applied to control for multiple comparisons. The association’s strength was reported as an odds ratio with 95% confidence intervals. We created regional association plots using LocuszoomCitation32 for the significant SNPs from the sex-stratified analysis. All statistical analysis was performed using Plink 1.90b6.2Citation30 and SAS 9.4 (SAS Institute Inc, Cary, NC).
Results
Population Characteristics
compares the characteristics of the study population between males and females. Female subjects were more likely to have self-reported physician-diagnosed asthma than males (14.0% vs 10.6%). The mean age of females was significantly greater than that of males (60.0 vs 59.1, p ≤ 0.0001). The distribution of smoking status, marital status, urban or rural dwelling, homeownership, total household, personal income, retirement status, province of recruitment, and highest education status also differed significantly between males and females. The proportion of current and former smokers was significantly greater in males than in females. The percentage of widowed, separated, or divorced females (21.6%) was significantly higher than that of males (9.6%).
Table 1 Characteristics of the Study Population
Results of SNP-by-Sex Interaction Analysis
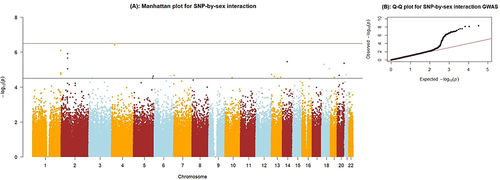
After quality control, 416,562 SNPs were included in the GWAS of SNP by sex interaction analysis. As indicated in , 49 variants showed a p-value less than 10−5 for the interaction term. The polymorphisms of rs7676077 and rs6701638 in/near the SLC34A2 and KIF26B genes (interaction p-values = 3.84×10−7 and 7.88×10−7 respectively) were close to reaching the genome-wide significance p-value threshold (). There was no evidence of population stratification, as indicated by the genomic inflation factor (λ = 1.025) in the Q-Q plot shown in .
Table 2 49 Signals of the SNP-by-Sex Interactions with p-value Less Than 10−5 for Asthma
Figure 1 (A) Manhattan plot for the SNP-by-sex interaction GWAS. The X-axis indicates the chromosomal positions of each variant and the Y-axis shows the negative Log p-values. The blue horizontal line indicates the suggestive significance threshold (1x10−5),and the red horizontal line shows the genome-wide signficance threshold (5x10−8). (B) Quantile-Quantile plot showing the distribution of observed versus expected p-values. The genomic inflation factor, lambda (λ) = 1.025.
Results of Sex Stratified Analysis
presents regional plots of the significant sex-specific SNPs from the sex-stratified multivariate survey logistic regression. After adjusting for multiple comparisons using Bonferroni correction at α ≤ 0.001 (0.05/49) significance level, we identified five male-specific SNPs (rs6701638, rs2968822, rs17071077, rs254804, and rs6013213) in/near KIF26B, RTN4, NMBR, PEPD, and NFATC2 loci and three female-specific SNPs (rs2968801, rs2864052 and rs9525931) in/near RTN4, and SERP2 genes (, and ). The polymorphism rs36213 in the EPHB1 gene was significantly associated with an increased risk of asthma [OR=1.35, p= 0.0004, 95% CI (1.14, 1.60)] in males and a reduced risk of asthma [OR=0.84, p= 0.0003, 95% CI (0.76, 0.92)] in females (, and ).
Table 3 Result of Sex Stratified Analysis for Asthma
Figure 2 Regional plots for the nine sex-specific SNPs show their respective locations. (A) SNP rs6701638 in KIF26B gene. (B) SNP rs17071077 near NMBR gene. (C) SNP rs254804 near PEPD gene. (D) SNP rs6013213 in NFATC2 gene. (E) SNP rs2968822 in RTN4 gene. (F) SNP rs2968801 near RTN4 gene. (G) SNP rs2864052 in RTN4 gene. (H) SNP rs9525931 near SERP2 gene. (I) SNP rs36213 in EPHB1 gene.
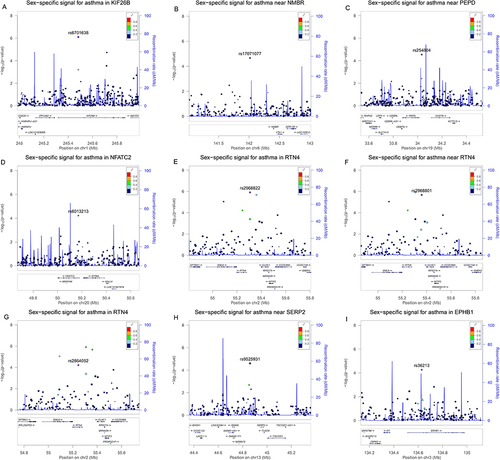
Figure 3 The forest plots show the odds ratio distribution for the sex-specific variants associated with asthma from the sex-stratified analysis. (A) Males. (B) Females. Only significant SNPs after Bonferroni adjustment were plotted. The blue diamonds represent effect sizes (Odds ratio), and the lines extending from the blue diamonds are the 95% confidence intervals.
In the additive genetic model, males with homozygous GG of rs6701638 (minor allele: G) in the KIF26B gene, homozygous GG of rs254804 (minor allele: G) near the PEPD gene, and homozygous TT of rs2968822 (minor allele: T) in RTN4 gene exhibited an increased risk of asthma (ORs: 1.24, 1.27, and 1.22) than those with wild-type (AA for rs6701638, TT for rs254804 and CC for rs2968822). In addition, males with homozygous GG of rs17071077 (minor allele: G) near the NMBR gene and homozygous AA of rs6013213 (minor allele: A) in the NFATC2 gene showed reduced risk of asthma (ORs: 0.79 and 0.77) compared to males with wild-type (AA for rs17071077 and GG for rs6013213).
Under the dominant genetic model, females with homozygous GG or heterozygous GA for rs2968801 (minor allele: G) and homozygous AA or heterozygous AG for rs2864052 (minor allele: A) in the RTN4 gene had a reduced risk of asthma (ORs: 0.79 and 0.77, respectively) compared to females with the wild-type (AA for rs2968801 and GG for rs2864052). Under the additive genetic model, females carrying homozygous GG for rs9525931 (minor allele: G) near the SERP2 gene demonstrated a lower risk of asthma (OR: 0.80) compared to females with the wild-type (AA for rs9525931).
Discussion
In this study, we identified sex-specific SNPs associated with asthma among middle-aged and older Canadians. Five male-specific SNPs (rs6701638, rs17071077, rs254804, rs6013213 and rs2968822) and three female-specific SNPs (rs2968801, rs2864052 and rs9525931) were significantly associated with asthma. Three of the five male-specific SNPs were associated with an increased risk of asthma, with an odds ratio ranging from 1.22 to 1.27. The other two male-specific SNPs were associated with a reduced risk of asthma (OR = 0.79 and OR = 0.77). All three female-specific SNPs were associated with a reduced risk of asthma, with ORs ranging from 0.77 to 0.80. The polymorphism rs36213 was significantly associated with an increased risk of asthma (OR = 1.35) in males but with a reduced risk of asthma (OR = 0.84) in females. In previous studies, male and female-specific genetic markers have been associated with asthma.Citation22,Citation25 However, these studies did not consider the influence of socio-demographic, socio-economic, and lifestyle factors.
The five male-specific polymorphisms, ie, rs6701638 in the KIF26B, rs254804 near the PEPD, rs17071077 near the NMBR, rs6013213 in the NFATC2, and rs2968822 near the RTN4 genes are located at 1q44, 19q13.11, 6q24.1, 20q13.2 and 2p16.1 genomic regions, respectively. The three female-specific polymorphisms, ie, rs2968801 near the RTN4, rs2864052 in the RTN4, and rs9525931 near the SERP2 genes, are located at 2p16.1 and 13q14.11 cytogenetic regions.
The polymorphism rs36213 (minor allele: G), which was significantly associated with an increased risk of asthma in males and a decreased risk of asthma in females, is an intronic variant in the EPH receptor B1 gene (EPHB1). EPHB1 is a receptor for Ephrin-B ligands (Ephrin-B1, Ephrin-B2, Ephrin-B3). EPH receptors are the largest family members of receptor tyrosine kinase (RTK), which are expressed in various immune cells, including CD4+ and CD8+ T cells, lymphocytes, monocytes, and granulocytes.Citation33–35 Furthermore, the EPH receptor-ephrin ligand interaction has been linked to immune cell activation, T cell differentiation, proliferation, and migration.Citation34,Citation35 Previous research has linked allergic rhinitis and asthma to EPH receptors, especially the EPHB2 gene, an important paralog of the EPHB1 gene.Citation36 Recently, animal studies have illustrated that sex hormones augmented the sex-specific effect of EPHB receptors and Ephrin-B ligands.Citation37–39 Wang et alCitation37 found that deleting EPHB4 from vascular smooth muscle cells of mice resulted in hypotension in males but not females. In a different experiment, female but not male Ephrin-B3 null mice had higher blood pressure and increased vascular smooth muscle cell contractions than wild-type.Citation38 In addition, estrogen enhanced increased vascular smooth muscle contraction in female Ephrin-b3 null mice compared with wild type, whereas testosterone reduced it.Citation38 These suggest that sex hormones may modulate ephrin-Eph receptor functions in males and females and, as such, may play crucial sex-specific role in asthma susceptibility.
This study also discovered three variants in/near the RTN4 gene, SNPs rs2968822 (minor allele: T), rs2968801 (minor allele: G), and rs2864052 (minor allele: A), associated with asthma in males and females, respectively. While the intronic SNP rs2968822 indicated an increased risk of asthma in males, Intergenic and intronic SNPs rs2968801 and rs2864052 showed a protective effect against asthma risk in females. RTN4 (Reticulon 4), commonly known as a neurite outgrowth inhibitor (Nogo) with three major isoforms, most notably NOGO-B (RTN4B), has previously been shown to be expressed in the lung tissue and airway epithelium.Citation40,Citation41 Numerous animal and human studies have examined the role of RTN4 in asthma and immune responses.Citation41–45 In an experimental study using mice, Wright et alCitation41 demonstrated that RTN4 inhibits Th2-mediated inflammation in the lungs, airway epithelium, and smooth muscles by showing that eosinophils levels and Th2 cytokines (IL-13, IL-4, and 1L-5) were higher in RTN-A/B knockout mice when compared to wild-type mice.Citation41 In addition, RTN4B has also been shown to enhance the production of the inflammatory cytokine after the stimulation of various nucleic acid-sensing toll-like receptors, including TLR9, TLR3, and TLR7, in macrophages and serum.Citation43 Studies have shown that males produce higher TNF-α (pro-inflammatory cytokine) and IL-10 (anti-inflammatory cytokine) than females after the stimulation of TLR3 and TLR9 or viral infection in peripheral blood mononuclear cells (PBMCs).Citation46 However, a review article reported that older men or men with androgen deficiency express higher pro-inflammatory cytokines and lower anti-inflammatory cytokines than older or menopausal women.Citation47 These suggest that sex hormones may distinctively impact RTN4-mediated expressions of Th2 immune responses, toll-like receptors, and the production of inflammatory and suppressive cytokines in males and females.
An intergenic SNP rs9525931 near the Stress-Associated Endoplasmic Reticulum Protein 2 gene (SERP2) had a female-specific association with asthma. Variants near the SERP2 gene have previously been associated with FEV1 and FEV1/FVC ratio in a GWAS of lung function and COPD.Citation48 Furthermore, CpGs and SNPs mapped to the SERP2 gene have been reported to be associated with Alzheimer’s disease in females.Citation49
An intronic variant rs6013213 (minor allele: A) in the NFATC2 gene was protective against the risk of asthma in males only. NFATC2, a member of the nuclear factor of activated T cells (NFAT) family, regulates Th cell immune response, differentiation, and the expression of induced cytokines such as IL-2, IL-3, IL-4, IL-10, and TNF-alpha.Citation50 Previous genome-wide association studies have associated several variants in the NFATC2 gene with asthma and allergic disease, including hay fever, eczema, and allergic rhinitis.Citation51–55 NFATC2 mRNA levels in PBMCs of allergic asthmatics were higher than in healthy non-asthmatic controls.Citation56 Furthermore, NFATC2 mRNA has been demonstrated to correlate positively with IL-5-induced eosinophils in asthma.Citation56 Animal and human studies have shown that estrogen and cigarette smoke exposure increased NFAT mRNA expression in the airways of females.Citation57,Citation58 Furthermore, NFAT has been reported to have a regulatory function in estradiol-mediated MUC5AC mRNA and protein expression in airway epithelial cells.Citation58 MUC5AC is a marker for mucus production in the airway epithelial cells. Thus, it is reasonable to suggest that NFATC2 may influence sex-hormone-induced mucus secretion differently in male and female asthmatic bronchial epithelial cells.
The intergenic variant rs17071077 (minor allele: G) near the Neuromedin B Receptor (NMBR) gene exhibited a significant protective effect against the risk of asthma in males. NMBR is a G protein-coupled peptide receptor that binds with the regulatory neuropeptide Neuromedin B (NMB).Citation59 NMB/NMBR, widely expressed in the lungs, broncho-epithelial cells, pulmonary neuroendocrine cells, brain, and testis at the protein and mRNA levels,Citation59–61 has been shown to promote fetal lung development.Citation62 Several studies have shown that NMBR plays an innate immune defense role against respiratory viral infection by enhancing IFN-alpha and reducing the expression of IL-6.Citation63 Females have been shown to have greater Type I IFN responses and are less susceptible to viral infections than males.Citation64 We reasonably infer that any abnormality or polymorphic alterations in the NMBR gene may have a higher impact on males.
In our study, males with the intergenic variant rs254804 near the PEPD gene had a significantly higher risk of asthma susceptibility. PEPD is a gene that encodes prolidase, an enzyme involved in collagen metabolism, wound healing, inflammation, angiogenesis, and cell growth.Citation65 Numerous studies have shown that airway remodeling in asthma is largely due to the deposition of extracellular matrix protein, including collagen fibers, around the airway smooth muscle layers.Citation66,Citation67 This suggests that polymorphisms of the PEPD gene may contribute to airway remodeling. Patients with mutations in the PEPD gene and dysfunctional prolidase enzyme have been reported to have asthma and asthma-like airway disease.Citation68,Citation69 Studies have shown that serum prolidase activity was associated with bronchial asthma.Citation70,Citation71 Higher serum prolidase levels appear to be associated with higher oxidative stress and lower antioxidant levels.Citation72,Citation73 The homeostatic balance between the oxidant and antioxidant systems is impaired in asthma,Citation74 with oxidative stress increasing tissue damage, triggering the production of pro-inflammatory mediators, and exacerbating airway inflammation.Citation74 Studies have shown that males exhibit higher levels of oxidative stress, oxidative stress biomarkers, reactive oxygen species, and lower antioxidant capacity than females.Citation75
In our current study, an intron SNP rs6701638 in the Kinesin Family Member 26B gene (KIF26B) exhibited a significantly increased risk of asthma susceptibility in males but not females. Several variants of the genes in the 1q43-q44 region have been associated with asthma and atopic asthma.Citation76 White et al demonstrated that a variant in the KIF26B gene within the 1q43-44 region was associated with atopic asthma.Citation76 However, the association’s p-value in that study was not significant after multiple testing adjustments. KIF26B is a target of the Wnt5a-Ror signaling pathway.Citation77 Studies have revealed that non-canonical Wnt5a-Ror signaling activation reduces KIF26B protein expression levels. Susman et alCitation77 demonstrated that the KIF26B protein expression level increased in embryonic fibroblast of Wnt5a null mice compared to the wild type. Dysregulation of Wnt5a signaling has been implicated in disrupted alveologenesis and the development of chronic lung disease, including asthma.Citation78 Furthermore, loss of Wnt5a in male mice resulted in abnormal reproductive organ development.Citation79 Taking together, we suggest that the alteration of the Wnt5a-Ror-KIF26B signaling pathway may have a deleterious impact in males and could play a crucial role in sex-specific asthma pathogenesis.
In our study, most sex-specific polymorphisms associated with asthma susceptibility may directly or indirectly interact with sex hormones in modulating immunoregulation, immune cell population, airway remodeling, oxidative stress, and lung function differently in males and females.
This study has strengths and limitations. A major strength was the size of the study population and the inclusion of lifestyle, socio-economic and socio-demographic factors in our analysis. Regarding the limitations, we used all asthma cases in our analysis without stratifying asthma by the age of onset (childhood onset and adulthood onset); this could potentially mask the identification of sex-specific loci associated with childhood and adulthood onset asthma. We reckon that misclassification of asthma is possible with self-reported physician-diagnosed asthma. Misclassification of the outcome variable may result in a bias towards the null. This may lead to an underestimation of the observed association. However, self-reported physician diagnosis of asthma has been used in genome-wide association and large population-based studies to identify sex-specific genetic and clinical characteristics of asthma.Citation22,Citation80 Our genome-wide SNP-by-sex interaction testing found no variants that met the genome-wide significance p-value criterion (p ≤ 5×10−8). Genome-wide interaction testing generally requires a very large sample size to achieve adequate statistical power to detect genome-wide significant interaction p-value. There was no independent replication cohort in our study to corroborate our findings. Future replication of these findings in an independent cohort will increase their generalizability. Our work lacks functional follow-up and enrichment analysis of sex-specific genetic variants associated with asthma; hence we cannot demonstrate a causal relationship. Future experimental investigations can address the underlying biological processes for the sex differences.
In conclusion, we found evidence of sex-specific polymorphisms associated with asthma susceptibility in/near the KIF26B, RTN4, EPHB1, NMBR, SERP2, PEPD, and NFATC2 genes. Most of the identified loci may potentially play direct or indirect roles in the immune modulatory mechanism, oxidative stress, and airway remodeling. Future studies aimed at finding sex-specific expressions of these loci and the related pathways of action would give functional insights into their sex-specific involvement in asthma pathogenesis.
Data Sharing Statement
The datasets used in this study are not readily available. Data are only available from the Canadian Longitudinal Study on Aging (www.clsa-elcv.ca) for researchers who meet the criteria for access to de-identified CLSA data.
Ethics Approval
This CLSA project received ethics approval at two levels. Consent to participate was obtained for all participants under the CLSA harmonized multi-university ethics process approved by the Hamilton Integrated Research Ethics Board (HiREB), Hamilton Health Sciences/McMaster University. Simon Fraser University (SFU) was a participating institution in the CLSA data collection, and the SFU Office of Research Services Ethics Committee reviewed all consent material prior to data collection (SFU ORS #2018s0139). This study was also conducted following the principles of the Declaration of Helsinki, and ethical approval was obtained from the University of Alberta Health Research Ethics Board (Pro00091377_REN3) and Memorial University Health Research Ethics Board (HREB # 2019.072).
Disclosure
The authors declare no conflicts of interest in this work.
Acknowledgment
This research was made possible using the data/biospecimens collected by the Canadian Longitudinal Study on Aging (CLSA). The Government of Canada provides funding for the Canadian Longitudinal Study on Aging (CLSA) through the Canadian Institutes of Health Research (CIHR) under grant reference: LSA 94473 and the Canada Foundation for Innovation, as well as the following provinces, Newfoundland and Labrador, Nova Scotia, Quebec, Ontario, Manitoba, Alberta, and British Columbia. This research has been conducted using CLSA’s Baseline Comprehensive Dataset version 4.0 and Genomics Dataset version 3.0, under Application Number 19CA006. The CLSA is led by Drs. Parminder Raina, Christina Wolfson, and Susan Kirkland. The opinions expressed in this manuscript are solely those of the author and do not represent those of the Canadian Longitudinal Study on Aging.
Additional information
Funding
References
- Safiri S, Carson-Chahhoud K, Karamzad N, et al. Prevalence, deaths, and disability-adjusted life-years due to asthma and its attributable risk factors in 204 countries and territories, 1990–2019. Chest. 2022;161(2):318–329. doi:10.1016/j.chest.2021.09.042
- Report from the Canadian Chronic Disease Surveillance System: Asthma and Chronic Obstructive Pulmonary Disease (COPD) in Canada, 2018. Available from: https://www.canada.ca/content/dam/phac-aspc/documents/services/publications/diseases-conditions/asthma-chronic-obstructive-pulmonary-disease-canada-2018/pub-eng.pdf. Accessed May 5, 2023.
- Postma DS. Gender differences in asthma development and progression. Gend Med. 2007;4:S133–S146. doi:10.1016/S1550-8579(07)80054-4
- Chowdhury NU, Guntur VP, Newcomb DC, Wechsler ME. Sex and gender in asthma. Eur Respir Rev. 2021;30(162):210067. doi:10.1183/16000617.0067-2021
- Ekpruke CD, Silveyra P. Sex differences in airway remodeling and inflammation: clinical and biological factors. Front Allergy. 2022;29(3):875295. doi:10.3389/falgy.2022.875295
- Fuseini H, Newcomb DC. Mechanisms driving gender differences in asthma. Curr Allergy Asthma Rep. 2017;17(3):19. doi:10.1007/s11882-017-0686-1
- Koleade A, Farrell J, Mugford G, Gao Z. Female-specific risk factors associated with risk of ACO (asthma COPD overlap) in aboriginal people. J Asthma. 2020;57(9):925–932. doi:10.1080/02770903.2019.1621890
- Wadden D, Allwood Newhook LA, Twells L, Farrell J, Gao Z. Sex-specific association between childhood BMI trajectories and asthma phenotypes. Int J Pediatr. 2018;2:e9057435.
- Gao Z, Dosman JA, Rennie DC, et al. Gender-specific associations between polymorphisms in the Toll-like receptor (TLR) genes and lung function among workers in swine operations. J Toxicol Environ Health A. 2018;81(22):1186–1198. doi:10.1080/15287394.2018.1544523
- Amegadzie JE, Gamble JM, Farrell J. Gender differences in inhaled pharmacotherapy utilization in patients with obstructive airway diseases (OADs): a population-based study. COPD. 2020;30(15):2355–2366. doi:10.2147/COPD.S264580
- Pignataro FS, Bonini M, Forgione A, Melandri S, Usmani OS. Asthma and gender: the female lung. Pharmacol Res. 2017;1(119):384–390. doi:10.1016/j.phrs.2017.02.017
- Chhabra SK, Chhabra P. Gender differences in perception of dyspnea, assessment of control, and quality of life in asthma. J Asthma. 2011;48(6):609–615. doi:10.3109/02770903.2011.587577
- Zillmer LR, Gazzotti MR, Nascimento OA, Montealegre F, Fish J, Jardim JR. Gender differences in the perception of asthma and respiratory symptoms in a population sample of asthma patients in four Brazilian cities. J Bras Pneumol. 2014;40:591–598. doi:10.1590/S1806-37132014000600002
- Blacquière MJ, Hylkema MN, Postma DS, Geerlings M, Timens W, Melgert BN. Airway inflammation and remodeling in two mouse models of asthma: comparison of males and females. Int Arch Allergy Immunol. 2010;153(2):173–181. doi:10.1159/000312635
- Takeda M, Tanabe M, Ito W, et al. Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma. Respirology. 2013;18(5):797–806. doi:10.1111/resp.12078
- McKenzie R, Burton MD, Royce SG, Tang MLK. Age and sex influences on airway hyperresponsiveness. J Asthma. 2010;47(6):651–654. doi:10.3109/02770901003692801
- Gao Z, Dosman JA, Rennie DC, et al. Association of Toll-like receptor 2 gene polymorphisms with lung function in workers in swine operations. Ann Allergy Asthma Immunol. 2013;110(1):44–50.e1. doi:10.1016/j.anai.2012.11.003
- Gao Z, Dosman JA, Rennie DC, et al. Effects of tumor necrosis factor (TNF) gene polymorphisms on the association between smoking and lung function among workers in swine operations. J Toxicol Environ Health A. 2021;84(13):536–552. doi:10.1080/15287394.2021.1896404
- Gao Z, Rennie DC, Senthilselvan A. Allergic rhinitis and genetic components: focus on Toll-like receptors (TLRs) gene polymorphism. Appl Clin Genet. 2010;16:109–120.
- El-Husseini ZW, Gosens R, Dekker F, Koppelman GH. The genetics of asthma and the promise of genomics-guided drug target discovery. Lancet Respir Med. 2020;8(10):1045–1056. doi:10.1016/S2213-2600(20)30363-5
- Odimba U, Senthilselvan A, Farrell J, Gao Z. Current knowledge of asthma-COPD overlap (ACO) genetic risk factors, characteristics, and prognosis. COPD. 2021;18(5):585–595. doi:10.1080/15412555.2021.1980870
- Espuela-Ortiz A, Herrera-Luis E, Lorenzo-Díaz F, et al. Role of sex on the genetic susceptibility to childhood asthma in latinos and African Americans. J Pers Med. 2021;11(11):1140. doi:10.3390/jpm11111140
- Mersha TB, Martin LJ, Biagini Myers JM, et al. Genomic architecture of asthma differs by sex. Genomics. 2015;106(1):15–22. doi:10.1016/j.ygeno.2015.03.003
- Gautam Y, Afanador Y, Abebe T, López JE, Mersha TB. Genome-wide analysis revealed sex-specific gene expression in asthmatics. Hum Mol Genet. 2019;28(15):2600–2614. doi:10.1093/hmg/ddz074
- Myers RA, Scott NM, Gauderman WJ, et al. Genome-wide interaction studies reveal sex-specific asthma risk alleles. Hum Mol Genet. 2014;23(19):5251–5259. doi:10.1093/hmg/ddu222
- Szczeklik W, Sanak M, Szczeklik A. Functional effects and gender association of COX-2 gene polymorphism G-765C in bronchial asthma. J Allergy Clin Immunol. 2004;114(2):248–253. doi:10.1016/j.jaci.2004.05.030
- Forgetta V, Li R, Darmond-Zwaig C, et al. Cohort profile: genomic data for 26 622 individuals from the Canadian Longitudinal Study on Aging (CLSA). BMJ Open. 2022;12(3):e059021. doi:10.1136/bmjopen-2021-059021
- Raina PS, Wolfson C, Kirkland SA, et al. The Canadian Longitudinal Study on Aging (CLSA). Can J Aging. 2009;28(3):221–229. doi:10.1017/S0714980809990055
- Raina P, Wolfson C, Kirkland S, et al. Cohort profile: the Canadian Longitudinal Study on Aging (CLSA). Int J Epidemiol. 2019;48(6):1752–1753j. doi:10.1093/ije/dyz173
- Purcell S, Chang C. General usage - PLINK 1.9 [Internet]; 2022. Available from: https://www.cog-genomics.org/plink/1.9/general_usage. Accessed May 5, 2023.
- Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, Lee JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience. 2015;4:S13742. doi:10.1186/s13742-015-0047-8
- Pruim RJ, Welch RP, Sanna S, et al. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 2010;26(18):2336–2337. doi:10.1093/bioinformatics/btq419
- Ivanov AI, Romanovsky AA. Putative dual role of ephrin-Eph receptor interactions in inflammation. IUBMB Life. 2006;58(7):389–394. doi:10.1080/15216540600756004
- Wu J, Luo H. Recent advances on T-cell regulation by receptor tyrosine kinases. Curr Opin Hematol. 2005;12(4):292–297. doi:10.1097/01.moh.0000166497.26397.9f
- Darling TK, Lamb TJ. Emerging roles for Eph receptors and ephrin ligands in immunity. Front Immunol. 2019;10. doi:10.3389/fimmu.2019.01473
- Wagener AH, Zwinderman AH, Luiten S, et al. The impact of allergic rhinitis and asthma on human nasal and bronchial epithelial gene expression. PLoS One. 2013;8(11):e80257. doi:10.1371/journal.pone.0080257
- Wang Y, Thorin E, Luo H, et al. EPHB4 protein expression in vascular smooth muscle cells regulates their contractility, and EPHB4 deletion leads to hypotension in mice. J Biol Chem. 2015;290(22):14235–14244. doi:10.1074/jbc.M114.621615
- Wang Y, Wu Z, Thorin E, et al. Estrogen and testosterone in concert with EFNB3 regulate vascular smooth muscle cell contractility and blood pressure. Am J Physiol Heart Circ Physiol. 2016;310(7):H861–H872. doi:10.1152/ajpheart.00873.2015
- Luo H, Wu Z, Tremblay J, et al. Receptor tyrosine kinase Ephb6 regulates vascular smooth muscle contractility and modulates blood pressure in concert with sex hormones. J Biol Chem. 2012;287(9):6819–6829. doi:10.1074/jbc.M111.293365
- Oertle T, Schwab ME. Nogo and its paRTNers. Trends Cell Biol. 2003;13(4):187–194. doi:10.1016/S0962-8924(03)00035-7
- Wright PL, Yu J, Di YPP, et al. Epithelial reticulon 4B (Nogo-B) is an endogenous regulator of Th2-driven lung inflammation. J Exp Med. 2010;207(12):2595–2607. doi:10.1084/jem.20100786
- Scatena R, Chupp GL, Perez MF, et al. The role of neurite outgrowth inhibitory protein-B in poorly controlled asthma. Am Thoracic Society Int Conf Abst. 2011:A1349–A1349. doi:10.1164/ajrccm-conference.2011.183.1_MeetingAbstracts.A1349
- Kimura T, Endo S, Inui M, Saitoh SI, Miyake K, Takai T. Endoplasmic protein Nogo-B (RTN4-B) interacts with GRAMD4 and regulates TLR9-mediated innate immune responses. J Immunol. 2015;194(11):5426–5436. doi:10.4049/jimmunol.1402006
- Xu W, Hong W, Shao Y, Ning Y, Cai Z, Li Q. Nogo-B regulates migration and contraction of airway smooth muscle cells by decreasing ARPC 2/3 and increasing MYL-9 expression. Respir Res. 2011;12(1):14. doi:10.1186/1465-9921-12-14
- Ullah HMA, Elfadl AK, Park S, et al. Nogo-A is critical for pro-inflammatory gene regulation in myocytes and macrophages. Cells. 2021;10(2):282. doi:10.3390/cells10020282
- Torcia MG, Nencioni L, Clemente AM, et al. Sex differences in the response to viral infections: TLR8 and TLR9 ligand stimulation induce higher IL10 production in males. PLoS One. 2012;7(6):e39853. doi:10.1371/journal.pone.0039853
- Klein SL, Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. 2016;16(10):626–638. doi:10.1038/nri.2016.90
- Sakornsakolpat P, Prokopenko D, Lamontagne M, et al. Genetic landscape of chronic obstructive pulmonary disease identifies heterogeneous cell-type and phenotype associations. Nat Genet. 2019;51(3):494–505. doi:10.1038/s41588-018-0342-2
- Zhang L, Young JI, Gomez L, et al. Sex-specific DNA methylation differences in Alzheimer’s disease pathology. Acta Neuropathol Commun. 2021;9(1):77. doi:10.1186/s40478-021-01177-8
- Fric J, Zelante T, Wong AYW, Mertes A, Yu HB, Ricciardi-Castagnoli P. NFAT control of innate immunity. Blood. 2012;120(7):1380–1389. doi:10.1182/blood-2012-02-404475
- Han Y, Jia Q, Jahani PS, et al. Genome-wide analysis highlights contribution of immune system pathways to the genetic architecture of asthma. Nat Commun. 2020;11(1):1776. doi:10.1038/s41467-020-15649-3
- Johansson Å, Rask-Andersen M, Karlsson T, Ek WE. Genome-wide association analysis of 350 000 Caucasians from the UK Biobank identifies novel loci for asthma, hay fever and eczema. Hum Mol Genet. 2019;28(23):4022–4041. doi:10.1093/hmg/ddz175
- Waage J, Standl M, Curtin JA, et al. Genome-wide association and HLA fine-mapping studies identify risk loci and genetic pathways underlying allergic rhinitis. Nat Genet. 2018;50(8):1072–1080. doi:10.1038/s41588-018-0157-1
- Ferreira MA, Vonk JM, Baurecht H, et al. Shared genetic origin of asthma, hay fever and eczema elucidates allergic disease biology. Nat Genet. 2017;49(12):1752–1757. doi:10.1038/ng.3985
- Hinds DA, McMahon G, Kiefer AK, et al. A genome-wide association meta-analysis of self-reported allergy identifies shared and allergy-specific susceptibility loci. Nat Genet. 2013;45(8):907–911. doi:10.1038/ng.2686
- Jakobi M, Kiefer A, Mirzakhani H, et al. Role of nuclear factor of activated T cells 2 (NFATc2) in allergic asthma. Immun Inflamm Dis. 2020;8(4):704–712. doi:10.1002/iid3.360
- Tam A, Churg A, Wright JL, et al. Sex differences in airway remodeling in a mouse model of chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2016;193(8):825–834. doi:10.1164/rccm.201503-0487OC
- Tam A, Wadsworth S, Dorscheid D, Man SFP, Sin DD. Estradiol increases mucus synthesis in bronchial epithelial cells. PLoS One. 2014;9(6):e100633. doi:10.1371/journal.pone.0100633
- Jensen RT, Battey JF, Spindel ER, Benya RV. International union of pharmacology. LXVIII. Mammalian bombesin receptors: nomenclature, distribution, pharmacology, signaling, and functions in normal and disease states. Pharmacol Rev. 2008;60(1):1–42. doi:10.1124/pr.107.07108
- Kaczyńska K, Zając D, Wojciechowski P, Jampolska M. Regulatory peptides in asthma. Int J Mol Sci. 2021;22(24):13656. doi:10.3390/ijms222413656
- Kane MA, Toi-Scott M, Johnson GL, Kelley KK, Boose D, Escobedo-Morse A. Bombesin-like peptide receptors in human bronchial epithelial cells. Peptides. 1996;17(1):111–118. doi:10.1016/0196-9781(95)02088-8
- Shan L, Emanuel RL, Dewald D, et al. Bombesin-like peptide receptor gene expression, regulation, and function in fetal murine lung. Am J Physiol Lung Cell Mol Physiol. 2004;286(1):L165–L173. doi:10.1152/ajplung.00436.2002
- Yang G, Huang H, Tang M, et al. Role of neuromedin B and its receptor in the innate immune responses against influenza A virus infection in vitro and in vivo. Vet Res. 2019;50(1):80. doi:10.1186/s13567-019-0695-2
- Pujantell M, Altfeld M. Consequences of sex differences in Type I IFN responses for the regulation of antiviral immunity. Front Immunol. 2022;13. doi:10.3389/fimmu.2022.986840
- Eni-Aganga I, Lanaghan ZM, Balasubramaniam M, Dash C, Pandhare J. PROLIDASE: a review from discovery to its role in health and disease. Front Mol Biosci. 2021;8. doi:10.3389/fmolb.2021.723003
- Araujo BB, Dolhnikoff M, Silva LFF, et al. Extracellular matrix components and regulators in the airway smooth muscle in asthma. Eur Respir J. 2008;32(1):61–69. doi:10.1183/09031936.00147807
- Parameswaran K, Willems-Widyastuti A, Alagappan VKT, Radford K, Kranenburg AR, Sharma HS. Role of extracellular matrix and its regulators in human airway smooth muscle biology. Cell Biochem Biophys. 2006;44(1):139–146. doi:10.1385/CBB:44:1:139
- Alrumayyan N, Slauenwhite D, McAlpine SM, et al. Prolidase deficiency, a rare inborn error of immunity, clinical phenotypes, immunological features, and proposed treatments in twins. Allergy Asthma Clin Immunol. 2022;18(1):17. doi:10.1186/s13223-022-00658-2
- Wang H, Kurien BT, Lundgren D, et al. A nonsense mutation of PEPD in four Amish children with prolidase deficiency. Am J Med Genet A. 2006;140(6):580–585. doi:10.1002/ajmg.a.31134
- Kaleli S, Akkaya A, Akdogan M, Gültekin F. The effects of different treatments on prolidase and antioxidant enzyme activities in patients with bronchial asthma. Environ Toxicol Pharmacol. 2006;22(1):35–39. doi:10.1016/j.etap.2005.11.001
- Cakmak A, Zeyrek D, Atas A, Celik H, Aksoy N, Erel O. Serum prolidase activity and oxidative status in patients with bronchial asthma. J Clin Lab Anal. 2009;23(2):132–138. doi:10.1002/jcla.20303
- Gecit I, Aslan M, Gunes M, et al. Serum prolidase activity, oxidative stress, and nitric oxide levels in patients with bladder cancer. J Cancer Res Clin Oncol. 2012;138(5):739–743. doi:10.1007/s00432-011-1136-4
- Pirinççi N, Kaba M, Geçit İ, et al. Serum prolidase activity, oxidative stress, and antioxidant enzyme levels in patients with renal cell carcinoma. Toxicol Ind Health. 2016;32(2):193–199. doi:10.1177/0748233713498924
- Sahiner UM, Birben E, Erzurum S, Sackesen C, Kalayci O. Oxidative stress in asthma. World Allergy Organ J. 2011;4(10):151–158. doi:10.1097/WOX.0b013e318232389e
- Kander MC, Cui Y, Liu Z. Gender difference in oxidative stress: a new look at the mechanisms for cardiovascular diseases. J Cell Mol Med. 2017;21(5):1024–1032. doi:10.1111/jcmm.13038
- White JH, Chiano M, Wigglesworth M, et al. Identification of a novel asthma susceptibility gene on chromosome 1qter and its functional evaluation. Hum Mol Genet. 2008;17(13):1890–1903. doi:10.1093/hmg/ddn087
- Susman MW, Karuna EP, Kunz RC, et al. Kinesin superfamily protein Kif26b links Wnt5a-Ror signaling to the control of cell and tissue behaviors in vertebrates. eLife. 2017;6:e26509. doi:10.7554/eLife.26509
- Li C, Smith SM, Peinado N, et al. WNT5a-ROR signaling is essential for alveologenesis. Cells. 2020;9(2):384. doi:10.3390/cells9020384
- Chawengsaksophak K, Svingen T, Ng ET, et al. Loss of Wnt5a disrupts primordial germ cell migration and male sexual development in mice1. Biol Reprod. 2012;86(1):1–12. doi:10.1095/biolreprod.111.095232
- Veerasingam E, Gao Z, Beach J, Senthilselvan A. Sex-specific characteristics for the coexistence of asthma and COPD in the Canadian population: a cross-sectional analysis of CLSA data. J Asthma. 2022;4:1–22.