
ABSTRACT
Research carried out on austral plesiosaurs from the Weddellian Biogeographic Province in the decade since the last attempt to characterise the New Zealand elasmosaurid, Mauisaurus haasti Hector, 1874 (On the Fossil Reptilia of New Zealand. Transactions and Proceedings of the New Zealand Institute 6: 333–358), has prompted a reappraisal of this taxon and a new consideration of its relationships. The hypodigms used in previous descriptions of the species are shown to include specimens from a number of different taxa, and the defining apomorphy of Mauisaurus, a hemispherical capitulum on the femur, has now been observed in specimens known to belong to different clades. Mauisaurus is now regarded as nomen dubium with possible affinities with the Subfamily Aristonectinae. A re-assessment of the most complete specimen previously assigned to Mauisaurus suggests it is a typical long-necked elasmosaurid closely comparable to Tuarangisaurus keyesi Wiffen and Moisley, 1986 (Late Cretaceous reptiles (Families Elasmosauridae and Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand. New Zealand Journal of Geology and Geophysics 29: 205–252).
Introduction
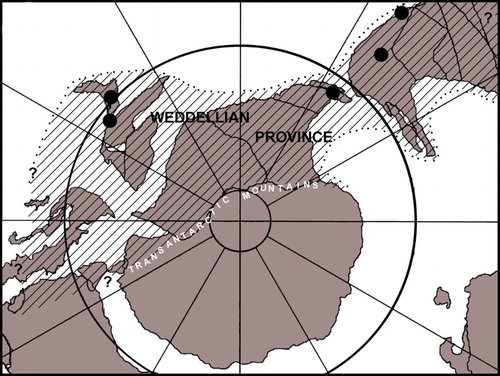
The genus Mauisaurus Hector, Citation1874 was established on material from the Upper Cretaceous of New Zealand and was one of the earliest plesiosaurian taxa to be recorded from the Southern Hemisphere. Although by today’s standards Hector’s (Citation1874) diagnosis would be regarded as woefully inadequate, from the time of its erection, the genus has been accepted as valid and numerous specimens from New Zealand and elsewhere have been assigned to it. It has even penetrated the awareness of the general public through postage stamps and coins dedicated to it () and through popular books on New Zealand fossils (e.g. Cox Citation1991; Long Citation1998). Throughout its long history, the composition of the hypodigm on which it was based has undergone a number of revisions (Welles Citation1962; Welles & Gregg Citation1971; Hiller et al. Citation2005). Now, as a result of discoveries made in South America and the Antarctic Peninsula over the last decade and the research these have supported (Otero et al. Citation2010, Citation2012; O’Gorman Citation2013; O’Gorman et al. Citation2014a, Citation2015; Otero et al. Citation2014, Citation2015a, Citation2015b), we believe that this is an appropriate time to re-examine Mauisaurus and all the specimens that have been assigned to it. Here, we examine Hector’s original concept of the taxon and the material on which it was based. We go on to look at how the concept evolved as more material was discovered in New Zealand and subsequently how more recent discoveries made in other parts of the Weddellian Biogeographic Province (; Zinsmeister Citation1979) have impacted on the concept.
Figure 1. A, Commemorative coin and B, C, postage stamps bearing an image of Mauisaurus.

Figure 2. The Weddellian Biogeographic Province in Late Cretaceous times, based on Zinsmeister (Citation1979: figure 3). Black dots indicate localities from which specimens mentioned in the text were derived.
The following institutional abbreviations are used in this article: AMNH, American Museum of Natural History, New York, USA; BMNH, Natural History Museum, London, UK; CM, Canterbury Museum, Christchurch, New Zealand; DM, Museum of New Zealand Te Papa Tongarewa, Wellington, New Zealand; GNS CD, GNS Science, Lower Hutt, New Zealand; KHM, Kaikoura Historical Museum, Kaikoura, New Zealand; MLP, Museo de La Plata, Argentina; MML, Museo Municipal de Lamarque, Río Negro, Argentina; SDSM, South Dakota School of Mines and Technology, Rapid City, South Dakota, USA; SGO.PV, Museo Nacional de Historia Natural, Santiago, Chile; TTU P, Museum of Texas Tech University, Lubbock, Texas, USA; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Original concept of Mauisaurus
When Hector (Citation1874) erected the genus Mauisaurus he diagnosed it as ‘centrum of dorsal vertebrae equal in length to the diameter, with smooth, concave sides, and an inferior mesial ridge; articular facets circular, flat, with a deep pit in the centre. Humerus with a large tuberosity.’ He assigned to it two species. The first of these, Mauisaurus haasti, was based on eight specimens (), Hector’s numbers 8a–h, comprising vertebrae, paddle bones, several pelvic elements and a few rib fragments, from three different localities (). Of these specimens, 8d and 8e are now missing and 8h turns out to be mosasaur teeth. Indeed, one tooth was figured by Knight (Citation1874) and identified as mosasaur in the same volume in which Hector’s paper appeared. The most significant of Hector’s specimens is 8a, comprising fragmentary pubes, a partial ilium and most of the right hind paddle (). Hector (Citation1874) mistakenly identified the bones as coming from the pectoral region of the skeleton rather than the pelvic.
Figure 3. Map showing the localities in New Zealand from which Mauisaurus has been recorded in the past.
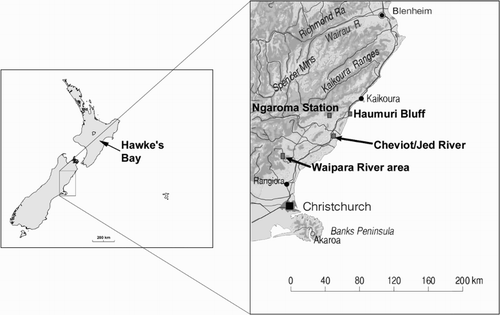
Figure 4. A copy of Hector’s (Citation1874) illustration showing the lectotype of Mauisaurus haasti.
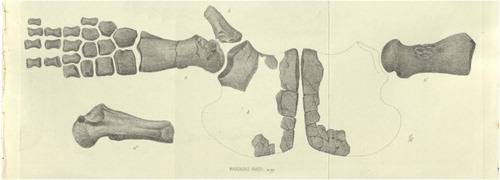
Table 1. Specimens used by Hector (Citation1874) in the original description of Mauisaurus haasti.
The second species assigned by Hector (Citation1874) to Mauisaurus is M. brachiolatus, based on two specimens; 9a the proximal end of a very large humerus and 9b a left humerus plus radius and radiale. The name of this species has been the source of some confusion. In the description of the bones, latibrachialis was the specific epithet applied (Hector Citation1874, p. 350) but in the list of specimens it is given as brachiolatus (Hector Citation1874, p. 336). Apparently, the former was a mistake that was corrected by Hector in an erratum slip. Hector gave no indication as to why he believed the two species, haasti and brachiolatus, should be placed in the same genus.
Welles (Citation1962), in a review of Cretaceous plesiosaurs, designated Hector’s specimen 8a as the lectotype of M. haasti. He regarded the pubes as being long and narrow and stated they could be pliosaurian (sensu lato). This view was apparently reinforced by the femur, which Welles (Citation1962) described as being very narrow distally and quite different from other plesiosaurs in having a hemispherical capitulum with a large trochanter dipping steeply posteriorly and separated from the capitulum by a broad groove (). Additionally, Welles (Citation1962) identified the very small fibular facet as a very unusual feature.
It appears that Welles (Citation1962) did not examine the specimen itself but based his opinion on his interpretation of the illustration provided by Hector (Citation1874, pl. 29). It seems that he failed to appreciate that this shows an oblique latero-ventral view of the femur, even though Hector (Citation1874, p. 363), in his explanation of the plate, pointed out that the paddle had nearly one-third greater width than shown.
Welles (Citation1962) placed M. brachiolatus as nomen vanum (nomen dubium in modern ICZN usage) as the material on which it is based is indeterminate.
Later modifications of the concept
Welles and Gregg (Citation1971), in their review of the Late Cretaceous marine reptiles of New Zealand, produced a revised diagnosis and the first detailed description of M. haasti based on the lectotype (DM R1529), one of the paralectotypes (Hector’s specimen 8g comprising seven posterior dorsal vertebrae; CM Zfr 92) and nine newly referred specimens (). They rejected from the hypodigm Hector’s specimens 8b, 8c and 8f, which they regarded as non-diagnostic, placing them in the Elasmosauridae indet. (DM R1530 and DM R1531) and Plesiosauria indet. (DM R829), respectively. Specimens 8d and 8e had already been lost by this time.
Table 2. Specimens referred by Welles and Gregg (Citation1971) to Mauisaurus haasti.
The specimens referred to M. haasti by Welles and Gregg (Citation1971) include Hector’s specimen 9a M. brachiolatus (DM R878), Hector’s specimen 9b M. brachiolatus (CM Zfr 88–90), Hector’s specimen 1c Plesiosaurus australis (trunk of a juvenile CM Zfr 103), BM R830 a broken distal humerus and ulna, CM Zfr 30 the left humerus of a juvenile, CM Zfr 95 an adult left femur, CM Zfr 102 anterior part of a pair of large coracoids, CM Zfr 104 anterior part of a right coracoid and associated vertebrae and paddle bones, and CM Zfr 109 the right femur of a juvenile. So, Welles and Gregg’s (Citation1971) hypodigm included juvenile as well as adult material from seven different localities.
More recently, in the last attempt to understand M. haasti, Hiller et al. (Citation2005) revised the hypodigm yet again. They rejected CM Zfr 92, DM R878, BM R830, CM Zfr 30 and CM Zfr 109, regarding each as non-diagnosable, but they added the new specimens CM Zfr 115, comprising some skull bones, an almost complete vertebral series and bones from all four limbs (see below) and KHM N 99–1079, pelvic bones and gastralia from a very large individual (). Once more, juvenile as well as adult material was included from seven localities. This new revision gave the first clear idea about the affinities of Mauisaurus, mostly based on CM Zfr 115, which, using current concepts, is a long-necked (at least 65 cervical vertebrae), non-aristonectine elasmosaurid (sensu Otero et al. Citation2012).
Table 3. Specimens included in Mauisaurus haasti by Hiller et al. (Citation2005).
Recent research and its implications
Plesiosaur specimens from outside New Zealand and referred to Mauisaurus have been recorded from Antarctica and South America. Fostowicz-Frelik and Gaździcki (Citation2001) tentatively referred to Mauisaurus sp. a postcranial skeleton from middle-to-upper Maastrichtian beds of Seymour Island (Marambio Island), Antarctica, based on the presence of the hemispherical head of the femur. Later, Gasparini et al. (Citation2003) referred two specimens from upper Maastrichtian beds of northern Argentinean Patagonia to cf. Mauisaurus sp. based on the similarities with the humeri referred by Welles and Gregg (Citation1971) to Mauisaurus; Martin et al. (Citation2007) also referred to Mauisaurus sp. a juvenile specimen from Maastrichtian levels of Vega Island, Antarctica, based on the presence of a rounded end of the femur, a conical ventral process of the coracoids, and fork-like gastralia. Subsequently, Otero et al. (Citation2010) referred to this genus two propodials from upper Maastrictian beds of central Chile, based on the presence of a distinctive large hemispherical femoral head, which then was still regarded as an autapomorphy of M. haasti, as pointed out previously (Welles Citation1962; Welles & Gregg Citation1971). Otero et al. (Citation2010) avoided a specific determination due to differences in size between the studied specimens and the lectotype of M. haasti.
In the latest revision of Patagonian and Antarctic elasmosaurids, made during research for a PhD thesis, one of the current authors (JPO’G) discussed whether Mauisaurus was present among the records from Argentinean Patagonia and the Antarctic Peninsula. One of the main conclusions was that a hemispherical femoral capitulum is present among members of the Aristonectinae. However, New Zealand specimen CM Zfr 115, placed in Mauisaurus by Hiller et al. (Citation2005), is a long-necked form with more than 60 cervical vertebrae and clearly not an aristonectine, thus raising the question about the validity of the concept of Mauisaurus. This result, with additional information from Kaiwhekea katiki Cruickshank & Fordyce, Citation2002 and Aristonectes quiriquinensis Otero et al. Citation2014, was discussed in detail by Otero et al. (Citation2015a) and it has become clear that femora with strongly hemispherical capitula are present in more than one aristonectine and also in the non-aristonectine elasmosaurid CM Zfr 115.
In recent publications, the current authors have questioned the validity of the M. haasti hypodigm. O’Gorman et al. (Citation2014b) showed that CM Zfr 104 belongs to the clade Aristonectinae, whereas CM Zfr 115 is a typical long-necked elasmosaurid. Also, Otero et al. (Citation2015a) have shown that the presence of hemispherical capitula on the femora, a character once believed to be unique to Mauisaurus and thus its defining apomorphy, is also seen in specimens that can be referred to other taxa, particularly A. quiriquinensis and K. katiki, both aristonectines. Also, the same authors noted that the hemispherical articular head can occur in both humeri and femora, as is the case in A. quiriquinensis.
The presence of a conical ventral process on the ventral side of the coracoids also appears to be present in more than one lineage so cannot be used as a diagnostic character of Mauisaurus as it was by Hiller et al. (Citation2005). Indeed, this feature is present in aristonectines (Otero et al. Citation2012; O’Gorman et al. Citation2014a) and in non-aristonectines (Welles Citation1952: figure 4a; Welles Citation1962: figure 5b). Therefore, it cannot be considered as a reliable character for genus-level determinations.
Another piece of significant research in recent years that can be brought to bear on our understanding of the marine reptiles attributed to M. haasti is the development of a refined biostratigraphy, based on dinoflagellate cysts, of the Upper Cretaceous deposits of New Zealand (Roncaglia et al. Citation1999). This research showed that the Conway Formation, the stratigraphic unit from which many marine reptile remains were recovered, is markedly diachronous, being older (middle–late Campanian) at Haumuri Bluff in the north than further south at Waipara River (late Campanian–early Maastrichtian) (). Extracting dinoflagellates from matrix adhering to reptile bones allowed Wilson et al. (Citation2005) to determine fairly precise ages for some individual marine reptile specimens. Such stratigraphic control contrasts with the provenance of most of the specimens from overseas that have been referred to Mauisaurus.
The New Zealand specimens attributed to M. haasti by Hector (Citation1874) and those who followed him (Welles & Gregg Citation1971; Wiffen & Moisley Citation1986; Hiller et al. Citation2005), and for which ages could be determined, are spread over an age range extending from middle Campanian to early Maastrichtian (), a time span of almost 10 million years (approximately 80–70 Ma). When Argentinean, Chilean and Antarctic specimens attributed to Mauisaurus are taken into account, the age range for the genus could be extended to the late Maastrichtian (see below). This seems an unusually long stratigraphic range for a single species, apparently without undergoing any evolution, given the plasticity shown by plesiosaurs throughout their fossil record.
Figure 5. Stratigraphic distribution of Mauisaurus specimens in New Zealand. See and for locality details.

Other specimens assigned to Mauisaurus
New Zealand
In addition to the specimens mentioned above, Wiffen and Moisley (Citation1986) assigned a specimen (GNS CD 430) comprising eight vertebrae, six dorsal and two sacral, the head of a propodial, two isolated phalanges, and rib and ischia fragments to M. haasti. They regarded this as the first specimen of Mauisaurus to have been recorded from the North Island and identified it based on the presence of a central swelling with a medial pit on the articular faces of the vertebra. They regarded this feature as a diagnostic character of Mauisaurus. However, as pointed out by Hiller et al. (Citation2005, p. 599), this feature is known from elasmosaurid taxa other than Mauisaurus (e.g. CM Zfr 145, a rather unusual specimen from New Zealand (Hiller & Mannering Citation2005) and A. quiriquinensis, among others) and, moreover, might depend on the ontogenetic stage of the individual. So Wiffen and Moisley’s specimen must be regarded as an indeterminate elasmosaurid. The central perforation on the articular facets likely represents the notochordal pit, and a paedomorphic condition could explain its presence in adult individuals such that described by Wiffen and Moisley (Citation1986).
South America
Elasmosaurid material from upper Maastrichtian beds of northern Patagonia was described by Gasparini et al. (Citation2003) and two specimens (MML‐PV3 and MML‐PV4) were identified as cf. Mauisaurus sp. The humerus of MML‐PV4 was likened to that of M. haasti, in particular to CM Zfr 90. MML‐PV3 preserves the proximal and distal ends of a femur, and although the capitulum is convex it does not appear to be as hemispherical as that of Mauisaurus. Also, the hemispherical condition was, at that time, exclusively regarded as diagnostic of the femur (but later verified to occur also in the humerus). MML‐PV3 also includes the intermedium of the hind limb, which Gasparini et al. (Citation2003) described as having a pentagonal shape similar to that of M. haasti (Hector Citation1874, pl. 29, figure a). Subsequently, Gasparini et al. (Citation2007) re-assessed this material and found that it was not identifiable to generic level, although it could be placed in the Elasmosauridae.
Otero et al. (Citation2010) reported on two specimens, a complete left femur (SGO.PV.135) and the proximal portion of another putative left femur (SGO.PV.169), from upper Maastrichtian beds of central Chile, which they regarded as being identifiable as Mauisaurus. A more recent re-assessment of this material by Otero et al. (Citation2015a) questioned this assignment and concluded that the specimens are referable to A. quiriquinensis. SGO.PV.169, previously considered as a large ‘femur’ was identified as the proximal part of a left humerus of this species. An additional conclusion by Otero et al. (Citation2015a) is that hemispherical heads are indeed present both in femora and humeri, and also, that such features depend on the ontogenetic stages of the specimens.
Otero et al. (Citation2015b) described several elasmosaurid specimens from Maastrichtian beds in the Magallanes Basin, southernmost Chile. Among them, a dwarf adult postcranial skeleton, recovered from lower Maastrichtian beds and identified as an aristonectine, was found to possess a hemispherical femoral capitulum. Otero et al. (Citation2015b) considered this specimen to be related to specimens from Antarctica (ZPAL R.8 and MLP 82-I-28-1) that at one time were regarded as closely related to Mauisaurus (see below).
Antarctica
In discussing the possible affinities of M. haasti, Hiller et al. (Citation2005) mentioned several specimens from the upper Maastrichtian of the Antarctic Peninsula region that they believed might possibly be related to that species. An indeterminate elasmosaurid specimen, TTU P 9221, was described by Chatterjee and Small (Citation1989) who observed central swellings with medial pits on the articular faces of dorsal centra. They noted that similar features were known in Mauisaurus but did not assign their specimen to the genus. As mentioned above, these features are not diagnostic.
Specimen (MLP 82-I-28-1), a partial hind limb with associated vertebral column, was described by Gasparini et al. (Citation1984) from Seymour Island (Marambio Island). This femur had a morphology, including a hemispherical head, that Hiller et al. (Citation2005) regarded as being very close to Mauisaurus. Gasparini et al. (Citation1984) remarked that they had not seen anything with a comparable morphology, an opinion reflecting that of Welles (Citation1962) when he nominated DM R1529 as the lectotype of M. haasti. The specimen was reviewed by one of the present authors (JPO’G) during research for a PhD thesis and placed as an indeterminate elasmosaurid.
Fostowicz-Frelik and Gaździcki (Citation2001) described a subadult elasmosaurid (ZPAL R.8) from lower Maastrichtian beds (Klb2) of Seymour Island (Marambio Island) stating that some features of the vertebral centra and the shapes of the tibia and proximal end of the femur suggested that the bones belonged to Mauisaurus or some closely related genus. However, they concluded that the remains could not be identified.
Gasparini et al. (Citation2003) referred to cf. Mauisaurus sp. another specimen (TTU P 9217), originally described by Chatterjee and Small (Citation1989), on the grounds that it was similar to one (MML‐PV4) they described from Patagonia. As shown above, the Patagonian specimen is regarded as an indeterminate elasmosaurid, so the Antarctic specimen must be similarly assigned.
Martin et al. (Citation2007) described the articulated skeleton of a juvenile plesiosaurian (SDSM 78156) from the Maastrichtian of Vega Island. Features of the femora and coracoids, and the presence of forked gastralia led them to identify the specimen as belonging in Mauisaurus but they were unable to provide a specific assignment. They regarded the specimen as being very similar to CM Zfr 103 from the Waipara River area of New Zealand. We show below that Mauisaurus cannot be adequately diagnosed so the Vega Island juvenile must be placed as Elasmosauridae indet., as was indicated by O’Gorman (Citation2013). In addition, the specimen (SDSM 78156) possesses anterior caudal centra with octagonal articular facets (RA Otero, pers. obs. 2013), which have been considered as diagnostic of the genus Aristonectes (Otero et al. Citation2012, Citation2014, Citation2015b).
Europe
Seeley (Citation1877) described a very large elasmosaurid (BMNH 47295) from the Gault of Folkestone, England, and identified it as Mauisaurus gardneri. The specimen, comprising cervical vertebrae and humerus, was later referred to the genus Plesiosaurus by Lydekker (Citation1889). Welles (Citation1962, p. 48) viewed the material as indeterminate although it showed elasmosaurid affinities; he considered the name to be a nomen vanum (dubium).
Discussion
It now seems apparent that the hypodigm of Mauisaurus, as modified by Welles and Gregg (Citation1971) and, most recently, by Hiller et al. (Citation2005), includes more than one taxon. The single significant autapomorphy of Mauisaurus identified by Welles (Citation1962) is the large hemispherical capitulum on the femur. However, this feature is now shown to be present in other taxa, rendering Mauisaurus non-diagnosable. In addition, the conical ventral midline process of the coracoids is known in specimens that do not belong to Mauisaurus. Indeed, a conical ventral process is also present in the juvenile specimen SGO.PV.260 from the upper Maastrichtian of Chile, referred to A. quiriquinensis (Otero et al. Citation2012, Citation2014); It is likely present on the juvenile former holotype of ‘Tuarangisaurus? cabazai’ Gasparini et al. Citation2003 from the upper Maastrichtian of Argentinean Patagonia, currently considered as an indeterminate aristonectine (O’Gorman et al. Citation2014a). Finally, the conical ventral process is present in Vegasaurus molyi, from the lower Maastrichtian of Antarctica (O’Gorman et al. Citation2015). Thus, the conical ventral process of the coracoids is found to occur in aristonectines as well as in non-aristonectine elasmosaurids, indicating that this feature cannot be considered as reliable for genus-level determinations
On the specimens used by Hiller et al. (Citation2005) in their description of M. haasti we can make the following observations, including the locality from which each was recovered and its age where determined.
DM R1529 remains the lectotype of M. haasti, but it must be regarded as nomen dubium as the specimen is not diagnosable. It is placed here as Elasmosauridae indet. The femur, with a generally stocky morphology and a hemispherical capitulum, bears a close resemblance to those of K. katiki and A. quiriquinensis, suggesting a possible assignment to the Aristonectinae (Otero et al. Citation2015a). However, the shapes of the epipodials, being wider than long, indicate, in our opinion, probable affinities to less derived elasmosaurids. The specimen has been dated as Middle Campanian (Wilson et al. Citation2005) Cheviot/Jed River ().
CM Zfr 90 is an isolated left humerus, with associated radius and radiale (CM Zfr 88 and 89, respectively). Heathstock, Waipara River area (). It could not be dated directly but is probably early Maastrichtian. This material shares with Kaiwhekea, Morenosaurus, Kawanectes and Vegasaurus the posterodistal extremity ending in an accessory facet (Welles Citation1943; O’Gorman et al. Citation2015; O’Gorman Citation2016). These four genera were recovered as monophyletic by O’Gorman (Citation2016) and therefore the presence of this humeral morphology in New Zealand is consistent with it.
CM Zfr 95 is a robust left femur that closely resembles that of DM R1529 and may belong to the same taxon but, coming from Birch Hollow, Waipara River area (), is probably younger (early Maastrichtian) although it could not be dated directly. It should be placed as Elasmosauridae indet.
CM Zfr 102 comprises the anterior portion of a pair of very large coracoids, regarded as the largest in a growth series by Hiller et al. (Citation2005). The features of the specimen include a ventral conical process commonly observed in several austral elasmosaurids (Sachs Citation2004; Hiller & Mannering Citation2005; Otero et al. Citation2012, Citation2014; O’Gorman et al. Citation2013, Citation2014b), allowing it to be identified as Elasmosauridae indet. and the large size might indicate an aristonectine affinity. Weka Creek, Waipara River area (). It could not be dated directly.
CM Zfr 103 is the trunk and tail of a juvenile specimen. This is Hector’s (Citation1874) specimen 1a, which Hutton (Citation1894) made the type of Cimoliasaurus caudalis (nomen dubium). It was placed in M. haasti by Welles and Gregg (Citation1971). The cordiform intercoracoid vacuity clearly indicates it belongs to the Elasmosauridae. This has an additional historical value because it was the first time that the cordiform vacuity, a highly diagnostic feature of the Elasmosauridae (O’Keefe Citation2001; Benson & Druckenmiller Citation2014), was described. Hiller et al. (Citation2005) used the coracoid as the smallest member of their growth series. One interesting feature of this specimen is the almost completely laterally directed diapophyses, differing from the dorsolateral direction observed in other elasmosaurids, such as Morenosaurus stocki (Welles, Citation1943), Elasmosauridae indet. (AMNH 6796; Colbert Citation1949), Styxosaurus sp. (AMNH 1495; Welles Citation1952; Otero Citation2016), among others. A similar feature was recorded in Kawanectes (O’Gorman Citation2016). Boby’s Stream, Waipara River area (); early Maastrichtian.
CM Zfr 104, recently described by O’Gorman et al. (Citation2014b), is an immature specimen comprising the anterior part of the right coracoid, several vertebrae (nine cervicals, one pectoral, eight posterior caudals) and a number of bones from one limb. Based on the proportions of the cervical vertebrae, O’Gorman et al. (Citation2014b) placed the specimen in the Aristonectinae. Weka Creek, Waipara River area (); early Maastrichtian.
CM Zfr 115 was used by Hiller et al. (Citation2005) as the basis for a new description of M. haasti but, given the results of recent research, it should be removed from this taxon and placed elsewhere (see below). It is a typical long-necked elasmosaurid with more than 65 cervical vertebrae. Ngaroma Station (); late Campanian.
KHM N 99–1079 is the part pelvis comprising ilium, ischium and pubis of a very large individual. It can be identified only as Plesiosauria indet. Haumuri Bluff (); middle Campanian.
The affinities of the Mauisaurus lectotype
The affinities of DM R1529 are difficult to assess because of the scarcity of useful features in the specimen. The ilium is ventrally stocky, but no other feature is observed (). The pubis () is fragmentary and the reconstruction provided by Hector (Citation1874) is probably quite speculative. The other preserved elements belong to a hind limb () that shows some interesting features:
Anterior and posterior knees. The development of anterior and posterior knees (distal expansions), giving a morphology very like that of a humerus, is very surprising; femoral knees are usually less developed than those of the humerus (Sato et al. Citation2006; O’Gorman et al. Citation2015). However, a similar shape is observed in A. quiriquinensis (Otero et al. Citation2014). The development of a convex distal posterior expansion (knee) in Mauisaurus is unusual as the posterior margins of femora are usually straight, as in Hydrotherosaurus alexandrae (Welles Citation1943, pl. 20d) and Callawayasaurus colombiensis (Welles Citation1962, pl. 4) or concave, as in Styxosaurus browni (Welles Citation1952, figure 7) and Thalassomedon haningtoni (Welles Citation1952, figure 17).
Concave epipodial facets. The presence of concave epipodial facets on the distal margin of the femur has been recorded in other elasmosaurids such as Futabasaurus suzukii (Sato et al. Citation2006, figure 8D). They are distinctly developed on the humeri of A. quiriquinensis, but the femora have epipodial facets comparatively less excavated. A very similar condition also occurs in K. katiki.
Dorsally extended and posteriodistally directed trochanter. The trochanter is almost completely visible in dorsal view and diagonally disposed, as has been recorded in the femora of K. katiki (Otero et al. Citation2015a, figure 2E), A. quiriquinensis (Otero et al. Citation2015a, figure 2A,C) and V. molyi (O’Gorman et al. Citation2015).
Hemispherical capitulum. The hemispherical capitulum of the femur was considered the diagnostic feature of M. haasti until the recognition of its presence among the Aristonectinae (Otero et al. Citation2015a).
Articular facets for a pisiform between fibula and fibulare. One of the most interesting features of the M. haasti lectotype is the presence of a postaxial notch between the fibula and fibulare. This feature was also recorded in A. quiriquinensis (Otero et al. Citation2014, figure 16A) but has not been properly described in other elasmosaurid hind limbs.
Epipodials comparatively short. DM R1529 has epipodials broader than long. Such proportions are commonly observed among Late Cretaceous elasmosaurids from the northern Pacific (e.g. H. alexandrae Welles, Citation1943) and from the Western Interior Seaway (e.g. S. browni Welles, Citation1952; H. alexandrae (Welles, Citation1943)). However, among known aristonectines preserving enough complete limbs, the epipodials are longer than broad, as occurs in K. katiki and A. quiriquinensis. In early Late Cretaceous elasmosaurids such C. colombiensis Welles, Citation1962 and Thalassomedon haningtoni Welles, Citation1943 the epipodials are also longer than broad.
DM R1529 shows some features that indicate aristonectine affinities – hemispherical femoral head, fibula–fibulare postaxial notch and diagonally disposed trochanter – but other features indicate affinities closer to the non-aristonectine elasmosaurids – short epipodials. Therefore, it is possible that this specimen belongs to a form that shows features intermediate between the two groups, although a more accurate determination is currently impossible.
Systematic palaeontology
Sauropterygia Owen, Citation1860
Plesiosauria de Blainville, Citation1835
Plesiosauroidea (Gray, Citation1825) Welles, Citation1943
Xenopsaria Benson & Druckenmiller, Citation2014
Elasmosauridae Cope, Citation1869
Tuarangisaurus sp.
(Figs. 6–12)
Mauisaurus haasti Hector, Citation1874: Hiller et al. Citation2005, 592–597, figs 6–17
Material
CM Zfr 115, a substantially complete skeleton comprising a partial skull, most of the spinal column, bones from all four limbs, and fragmentary remains of both pectoral and pelvic girdles along with fragmentary ribs and gastralia. Numerous gastroliths accompany the bones.
Formation and locality
Conway Formation; Ngaroma Station, on the Conway River, North Canterbury (). The site is high on a hillside, scarred by numerous landslips (New Zealand Fossil Record File number 032/f8862).
Age
Dinoflagellate cysts extracted from the rock adhering to the plesiosaur bones indicate a late Campanian age for the specimen (Wilson et al. Citation2005).
Collectors
As reported by Welles and Gregg (Citation1971, p. 18), the specimen was retrieved between November 1969 and March 1970, by Messrs G. Warren (New Zealand Geological Survey), N. M. Hyde (Ngaroma Station), R. W. Tyree (Christchurch) and S. P. Welles (University of California), with the assistance of various members of the Warren, Hyde and Tyree families and lodged in Canterbury Museum, Christchurch.
Taphonomy
CM Zfr 115 was preserved in a flat calcareous slab dissected by many calcite veins. Removal of most of the veins by weathering reduced the slab to a number of small blocks that were subsequently separated by movements of a landslip. Skull fragments and some anterior cervical vertebrae were completely weathered out of the matrix, but the rest of the skeleton was recovered in the matrix, albeit in fragments. Final preparation of the skeleton was achieved using mechanical techniques because there was good separation of matrix from bone.
Although no attempt was made to reconstruct the original relationships of the blocks after collection, once preparation was complete, the bones were laid out in what was believed to be their most likely resting positions prior to burial. These indicated that the right fore-paddle and left rear-paddle lay ventral side up and the disarticulated left coracoid had come to lie behind the right, with both displaced posteriorly. The other limbs and caudal vertebrae were disassociated, with no clear relation to the rest of the skeleton. The carcass was interpreted as having come to rest on its right side anteriorly, but from the sacral series posteriorly the skeleton seems to have twisted so as to lie ventral side up. Evidence of some scavenging was provided by disruption of the skeleton and the presence of three hexanchid shark teeth adjacent to the bones of the right front paddle.
Description
The specimen CM Zfr 115 formed the basis of the last attempt to understand Mauisaurus, presented by Hiller et al. (Citation2005) and it was described in detail in that publication. Only parts of that description are repeated here, with some modification. In the vertebrae of the cervical series and in the preserved caudals, the neural arches are not firmly fused to their centra, whereas in the dorsal vertebrae, neural arches and centra are firmly fused. This situation leads us to interpret the specimen as the skeleton of an individual that had not reached full maturity at the time of death.
Skull remains (Figure 6)
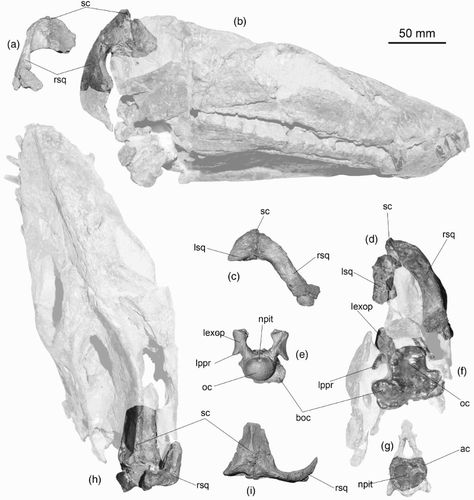
CM Zfr 115 preserves a partial braincase comprising the incomplete basioccipital, most of the basisphenoid and both exoccipital–opisthotics (D–G). It also preserves the right dorsal ramus of the squamosal, which is joined to the posterior part of the parietals (A–C), and part of the left lower jaw ramus, including the glenoid (H,I). The posterior width of the skull is estimated at 140 mm based on the distance between the midline and the exterior margin of the squamosal.
Figure 6. CM Zfr 115 skull bones. A–C, preserved portions of parietal and right squamosal in A, right lateral; B, posterior view with bilateral symmetry restored; C, dorsal view with bilateral symmetry restored. D–G, brain case elements. D, Basioccipital and exoccipital–opisthotics in posterior view. E, Basioccipital in ventral view. F, Basisphenoid in ventral view (anterior towards top of page). G, Basisphenoid in anterior view. H, I, Posterior of incomplete right lower jaw ramus in H, internal view, and I external view. J, Right exoccipital–opisthotic in anterior view. K, Right exoccipital–opisthotic in medial view. a, angular; acc, ascending canal; agp, anterior glenoid process; exop, exoccipital–opisthotic; f, foramina; fbo, facet for basioccipital; fenov, position of fenestra ovalis; feo, facet for exoccipital; fso, facet for supraoccipital; gl, glenoid; gr, condylar groove; jfo, jugular foramen; npit, notochordal pit; oc, occipital condyle; p, parietal; pgp, posterior glenoid process; ppr, paraoccipital process; ptf, pterygoid facet; ra, retroarticular process; sc, sagittal crest; sq, squamosal; ut, depression for utriculus; vr, ventral ridge; (IX + X), posterior cranial nerves (glossopharyngeal and vagus) emerge through jugular foramen; XI, foramen for cranial nerve XI (accessorius); XII, foramen for cranial nerve XII (hypoglossal). Cross-hatch shading indicates broken bone surface; open-circle shading indicates adhering sediment. Modified after Hiller et al. Citation2005.
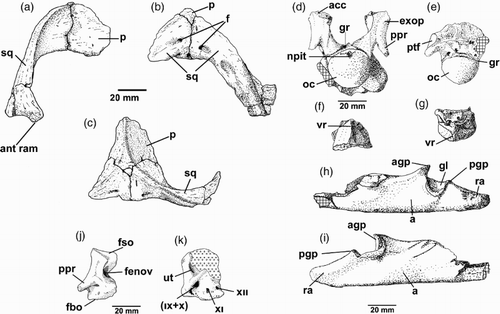
The basioccipital and basisphenoid are not fused. The paroccipital processes of the exoccipital–opisthotics are incomplete, but were apparently short, strap-like and directed downwards. The exoccipital–opisthotics have foramina for the posterior cranial nerves IX, X, XI exiting from the jugular foramen, with XII emerging from a separate foramen behind these (Hiller et al. Citation2005). Internally, there are distinct foramina for the jugular (with IX, X) another for XI and a third for XII (J,K). There is a substantial excavation for the fenestra ovalis (J). The vertical semi-circular canals are almost entirely in the supraoccipital, with only small openings in the upper surface of the exoccipital–opisthotics. The occipital condyle is subcircular in outline and, in posterior view, shows a slight depression along the dorsal margin where a shallow circular notochordal pit is situated (D). The condyle arises solely from the body of the basioccipital and a shallow groove separates it from the rest of the basioccipital. Posteriolaterally, the basioccipital bears subcircular facets for articulation with the pterygoids. In posterior view (D), the basioccipital appears asymmetrical below the condyle with the right side extending further from the condyle than the left. The left side may have suffered some damage. The anterior surface of the basioccipital bears a deeply pitted transversely ovoid facet for contact with the basisphenoid. The basisphenoid, which possesses a ventral keel, is broken on its anterior face. It shows the course of the carotid arteries that pierced the bone.
The right squamosal is represented by the dorsal ramus, extending from the parietal suture to the point where it sweeps forwards into the anterior ramus. The left squamosal is represented only by its dorsal-most portion immediately adjacent to the parietal suture (A–C). The squamosals are laterally broad and axially compressed.
The parietals are represented by the posterior portions only; the median suture between the two bones is not clear and they seem to be firmly fused. The ventromedial surfaces are heavily roughened for attachment of the supraoccipital, and an irregular ridge extends along the ventral midline where a suture might be expected. The dorsolateral surfaces of the parietals are gently concave and meet at an acute angle to form a sharp sagittal crest (C). The crest bifurcates posteriorly and extends down each squamosal as a sharp ridge. About one-third of the way down the dorsal ramus, the ridge divides to define the anterior margin of the dorsal ramus and thence the dorsal margin of the anterior ramus, and to separate the posterior surface of the dorsal ramus from its lateral surface. The posterior surface of the squamosal is marked by a series of grooves and foramina.
The left lower jaw fragment (H,I) is c. 120 mm long and has very high lips to the glenoid fossa, with the anterior lip being higher than the posterior. The maximum vertical thickness of the jaw, measured at the posterior margin of the glenoid is only three-quarters that measured at the anterior margin of the glenoid. The elements comprising this part of the mandible are firmly fused and the sutures between them are obscure.
Axial skeleton
In the previous description of CM Zfr 115, Hiller et al. (Citation2005) followed Carpenter (Citation1999) in not designating any vertebrae as pectorals. In this account, we have chosen to recognise pectoral vertebrae in the manner of Welles (Citation1943), leading to an apparent discrepancy in the cervical counts between to two papers.
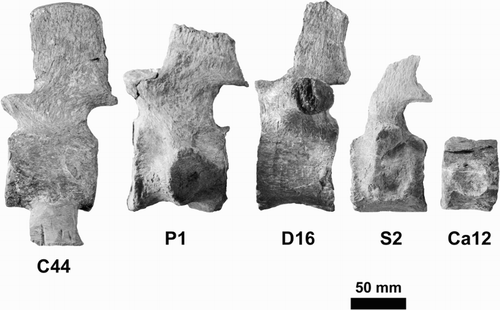
There are 63 vertebrae preserved in the neck region, but no atlas or axis. As explained in Hiller et al. (Citation2005, p. 594), and judging by the relative size of the occipital condyle and the first preserved cervical vertebra, we believe that at least one more vertebra is missing from the sequence; thus, a count of a minimum of 66 cervical vertebrae is likely. There are two pectoral vertebrae, 18 dorsal vertebrae, three sacral vertebrae and at least 20 caudals. Typical examples of each are shown in . The vertebrae were described in detail and figured by Hiller et al. (Citation2005) when CM Zfr 115 was regarded as belonging to Mauisaurus; that description is not repeated here. A table of the dimensions of the vertebrae and an analysis of their shapes were presented by O’Keefe and Hiller (Citation2006).
Figure 7. Typical examples of vertebrae from different regions of the spinal series. C44, mid-cervical; P1, first pectoral; D16, posterior dorsal; S2, second sacral; Ca12, mid-caudal.
Limbs
Fragments of all four limbs are present in CM Zfr 115. The right humerus has most of the shaft and part of the head preserved, but the shaft has been split lengthwise, and part of the distal end is missing (A). The capitulum is gently convex with an elliptical outline. It also shows the tuberosity to be slightly prominent. The right femur is preserved in two main parts, with a small portion of the diaphysis missing. This bone has a strongly convex capitulum with an almost circular outline (B). The capitulum is well separated and angled with respect to the rest of the diaphysis. The trochanter is dorsally flattened. The distal ends of both femora and humeri are distinctly angled, with clear non-concave facets for the epipodials.
Figure 8. CM Zfr 115 propodials. A, Right humerus. B, Right femur.

A reasonably well preserved right fore paddle () shows an elongately ovoid epipodial foramen between the radius and ulna. Both epipodials are significantly broader than long. Typical elasmosaurid features in this paddle include the proximal row of carpals comprising the radiale, centrale and ulnare; the distal row made up of distal carpal I, fused distal carpals II and III, and distal carpal IV. Metacarpal V has been displaced to rest alongside both the distal carpal row and the row containing the other metacarpals (O’Keefe Citation2001).
Figure 9. CM Zfr 115 right fore paddle in A, dorsal and B, ventral views. ce, centrale; dc, distal carpal; ef, epipodial foramen; mc, metacarpal; r, radius; re, radiale; ph, phalanx; u, ulna; ue, ulnare. Arrows indicate the positions of hexanchid shark teeth.

Girdles
The pectoral girdle of CM Zfr 115 includes well-preserved scapulae, the right almost complete and undistorted. This shows a typical triradiate structure (), with the angle between the ventral and dorsal rami, measured at the ridge on the ventral surface where they meet, c. 130° (). The dorsal (ascending) ramus is high, with almost parallel sides and a flat (‘square’ rather than tapered) distal margin (). The remainder of the bone is thick, with pockmarked surfaces for cartilaginous extensions on the anteromedial surfaces.
Figure 10. CM Zfr 115 right scapula in medial view. cf, coracoid facet; dr, dorsal ramus; gl, glenoid; sym, symphyseal surface; vr, ventral ramus.

The coracoids are fragmentary with much of their mass missing and cannot be reconstructed accurately. The anterior portions form the midline junction between the paired coracoids; they are deep along the midline, with an inverted V-shape in profile, suggesting the presence of a transverse dorsal thickening and a ventrally directed, blunt, conical process. Along the midline each coracoid extends anteriorly into a blunt process with a V-shaped notch between them.
As pointed out by Hiller et al. (Citation2005), in no New Zealand elasmosaurid specimen are there bones that can be identified as clavicles or interclavicles. If these elements were present in life, they probably were never ossified.
The elements of the pelvic girdle are also fragmentary in CM Zfr 115 and the detailed morphologies of the component parts are not entirely clear. However, pieces of what we interpret to be the right pubis and right ischium () allow a partial description to be made. The pubis is preserved in three fragments that can be joined by aligning the few attached gastralia and, although the contact between fragments has been lost, the general outline of the bone can be observed. It has a dorsoventrally deep ischial facet, with a concave acetabular surface. It bears a rounded, anterolateral prominence. There is a poorly developed symphyseal contact; no posterior symphyseal extension is present. The ischium is also preserved in three parts, one preserving the pubic facet that precisely matches the respective pubis contact. The two other fragments are attached by matrix and show a rounded symphyseal margin but any anterior extension has been lost, as has the acetabular margin. The lack of broad symphyseal contact between the pubes and the ischia is consistent with a subadult ontogenetic stage (Brown Citation1981).
Figure 11. CM Zfr 115 right pubis and right ischium in dorsal view.

Ribs and gastralia are too fragmentary for meaningful description.
Comparisons
With other New Zealand elasmosaurids
Comparisons can be drawn between CM Zfr 115 and several elasmosaurids described from New Zealand. Of these, only one, Tuarangisaurus keyesi Wiffen & Moisley, Citation1986, has been named; the others are represented by incomplete specimens that cannot be fully diagnosed. CM Zfr 104 is a juvenile aristonectine (O’Gorman et al. Citation2014b) possessing typical short cervical vertebrae that immediately separate it from CM Zfr 115. CM Zfr 145 is an unusual form that Hiller and Mannering (Citation2005) believed could be separated from all previously described elasmosaurids. In particular, its scapulae and coracoids show marked differences from the same bones in CM Zfr 115. In the latter, the angle between the dorsal and ventral rami of the scapula is 130°, the dorsal ramus has parallel sides with a square termination. In CM Zfr 145 the angle between dorsal and ventral rami is 140° and the dorsal ramus tapers distally and has a rounded termination. The coracoids of CM Zfr 145 are firmly fused anterior of the intercoracoid vacuity unlike the situation in CM Zfr 115 where the coracoids are separate. The coracoids of CM Zfr 145 are flat and plate like with no sign of the dorsal transverse thickening seen in CM Zfr 115 and on the ventral surface they bear a rod-like projection quite unlike the rounded conical ventral process of CM Zfr 115.
CM Zfr 115 can be compared with the T. keyesi holotype from Hawke’s Bay () insofar as they have several anterior cervical vertebrae and several skull bones in common. In comparing the skull elements of CM Zfr 115 with those in T. keyesi (GNS CD 425 and 426) (), we can make the following observations:
Figure 12. Comparison between skull fragments of CM Zfr 115 and the skull of Tuarangisaurus keyesi Wiffen and Moisley Citation1986 (GNS CD 425, holotype). A, CM Zfr 115, right squamosal and part of the right parietal in right lateral view. B, GNS CD 425 in right lateral view. Equivalent elements are digitally enhanced. C, CM Zfr 115, squamosals in occipital view. D, GNS CD 425, occipital view of the skull, with the squamosals digitally enhanced. E, CM Zfr 115, occipital condyle and exoccipital–opistothics articulated, in occipital view. F, Equivalent elements digitally enhanced on GNS CD 425 skull. G, GNS CD 426, anterior view of the atlantal cup of the atlas–axis. The notochordal pit is visible, as it happens on the CM Zfr 115 occipital condyle, however, in the latter this has a comparatively more dorsal position. H, GNS CD 425, dorsal view of the skull with posterior part of the sagittal crest and the squamosals digitally enhanced for comparison. I, CM Zfr 115, sagittal crest and squamosals in dorsal view. ac, atlantal cup; boc, basioccipital; lexop, left exoccipital-opisthotic; lppr, left paraoccipital process; lsq, left squamosal; npit, notochordal pit; oc, occipital condyle; rsq, right squamosal; sc, sagittal crest.
Squamosals: the squamosal of CM Zfr 115 is similar to that of the T. keyesi holotype. Both specimens preserve a right squamosal in which the dorsal ramus is axially compressed. Also, in both specimens the squamosals join dorsally in the midline forming a bulb. In posterior view, both squamosals are remarkably similar, having a dorsal ramus with a flat posterior surface and a cross-section with a conserved breadth that only expands near the dorsal midline.
Parietals: even though the posterior part of the skull of T. keyesi is crushed laterally it is possible to assess that the parietals in each specimen are remarkably similar. In both, the parietals form a low sagittal crest with a flat dorsal surface having an axially extended triangular outline.
Basioccipital: the basioccipital of T. keyesi is broken. The portion anterior to the occipital condyle is still attached in its anatomical position within the skull. Even though this is laterally crushed, the basioccipital contour does not reflect evident deformation. In posterior view, the broken section of the T. keyesi basioccipital shows a ventrally massive bone, twice as broad as the dorsal portion where the occipital condyle is expected to be attached. By comparison, the lateral extension of the ventral basioccipital in CM Zfr 115 appears to be less expanded, which could reflect an earlier ontogenetic stage. Because the T. keyesi basioccipital is posteriorly broken, it is difficult to evaluate the shape of the pterygoidal articulation, only visible on the right side of the CM Zfr 115 basioccipital.
The occipital condyle of T. keyesi is broken away, making it impossible to compare it with that of CM Zfr 115. The outline of the atlantal cup of T. keyesi is very similar to the occipital condyle outline of CM Zfr 115, but in the latter, the notochordal pit lies in the dorsal part of the condyle, whereas in the T. keyesi atlantal cup, the notochordal pit appears to be in a central position.
Basisphenoid: the basisphenoid of CM Zfr 115 is separated from the rest of the braincase. Its preserved length is near the same as the basioccipital excluding the condyle. Ventrally, it possesses a sharp median keel (F). In T. keyesi, the basisphenoid is obscured by matrix, however, O’Gorman et al. (Citation2017) have assessed its basisphenoid via computed tomography scan. Interestingly, this also has a sharp ventral keel, but the extension of the bone seems to be axially larger. This could be the result of the combined length of the basisphenoid and the anterior part of the parasphenoid.
Exoccipital–opisthotics: these elements in CM Zfr 115 and T. keyesi are remarkably similar. In posterior view, the T. keyesi skull allows us to observe a good deal of the external surface of the left exoccipital–opisthotic. Above the paraoccipital process, both specimens possess a slightly concave dorsolateral surface. Both also have a visible suture that extends dorsoventrally over their posterior surface, likely belonging to the contact between opisthotic and the exoccipital. The paraoccipital process of both specimens is very similar, being ventrally recurved at c. 45°, having a short and dorsoventrally compressed shaft with oval cross section. In the T. keyesi holotype, the distal end of its left paraoccipital process is broken, suggesting the presence of a shaft slightly larger than that of CM Zfr 115. Such a difference could be expected due to the different ontogenetic stages of the specimens.
Mandibular rami: CM Zfr 115 only preserves a posterior part of the left ramus. This includes the retroarticular process, the glenoid, part of the coronoid and a posterior fragment of the dentary. Regrettably, both rami of T. keyesi are broken just after the end of the dentition, being absent a good deal of the coronoid (part of it is obscured by matrix), while the glenoid and retroarticular processes are lost. The rami of both specimens share the presence of bowed mandibles in ventral view, however, this is a trait typical of most elasmosaurids (O’Keefe Citation2001).
Wiffen and Moisley (Citation1986) attributed a small section of a left angular, quadrate and squamosal (GNS CD 426) to the holotype of T. keyesi and these can be compared with CM Zfr 115. These show that the lower jaw glenoids of both CM Zfr 115 and Tuarangisaurus have enlarged glenoid processes, with the anterior one being higher than the posterior, although this feature is more marked in CM Zfr 115.
In addition to the skull elements, both CM Zfr 115 and the T. keyesi holotype show comparable characters in the neck, such as the anterior and posterior zygapophyses becoming conjoined from about the ninth or tenth cervical vertebra posteriorly, but their vertebral length indices (Brown Citation1981) differ (). Although we cannot be certain about the serial position of the cervical vertebrae in CM Zfr 115, it appears that it has consistently lower values (c. 101 vs. 113) for the first seven preserved centra other than atlas and axis. The significance of this difference is uncertain and it may be accounted for by ontogenetic differences between the holotype of T. keyesi, a somewhat more mature animal (judging by the comparatively larger skull elements that the two specimens have in common), and the subadult CM Zfr 115. Brown (Citation1981, p. 269 and figure 13) has shown that vertebral indices change during ontogeny.
Table 4. Vertebral length index (VLI) for the anterior cervical vertebrae (excluding atlas and axis) for CM Zfr 115 (N) and Tuarangisaurus (GNS CD 426; T). VLI is calculated as L/D × 100, where L = mid ventral centrum length and D = posterior average centrum diameter (Brown Citation1981). Lack of data is indicated by —. The vertebra number for CM Zfr 115 is the number of the preserved centrum as explained in Hiller et al. (Citation2005); for Tuarangisaurus the vertebra number represents the actual place occupied in the series by the centrum, where atlas and axis are 1 and 2, respectively.
In addition to the vertebral length indices (Brown Citation1981) analysed above, the cervical vertebrae of the T. keyesi holotype and those of CM Zfr 115 share several morphological traits. In articular view, both specimens have neural pedicels that are thin and high, leaving a dorsoventrally extended, oval neural canal. Both also share the presence of prezygapophyses slightly broader than the neural arch. In lateral view, the cervical centra of both specimens have lateral keels. The few preserved neural spines in CM Zfr 115 allow us to assess that the prezygapophyses of both specimens similarly extend beyond the articular faces of their respective centra. Although these traits are not diagnostic at genus level, and are typical of austral, non-aristonectine elasmosaurids (Hiller et al. Citation2005), the cervical features of both specimens suggest close affinities between them.
Comparison with other Late Cretaceous Weddellian elasmosaurids
The features of the axial skeleton and limbs of CM Zfr 115 differ from those of the aristonectines. In particular, the vertebral proportions of CM Zfr 115 indicate relatively longer and narrower centra than found among the aristonectines (Cruickshank & Fordyce Citation2002; Gasparini et al. Citation2003; Otero et al. Citation2014). In addition, the ulnae and radii of aristonectines are longer than wide (Otero et al. Citation2014), contrary to the situation in CM Zfr 115.
The features of CM Zfr 115 also differ from the two non-aristonectines known from the Weddellian Province, V. molyi O’Gorman et al. Citation2015 and Kawanectes lafquenianum (Gasparini and Goñi) O’Gorman, Citation2016. CM Zfr 115 has a cervical vertebral count of at least 65 elements, while V. molyi has only 54 cervicals (and 3 versus 2 pectorals). Vegasaurus molyi also has a scapular ridge (O’Gorman et al. Citation2015, figure 8B,C), a feature not seen in the New Zealand specimen. Kawanectes lafquenianum differs from CM Zfr 115 in the greater lateral extension of its caudal parapopyses (O’Gorman Citation2016).
Comparison with North American Late Cretaceous elasmosaurids
The axial formula of CM Zfr 115 indicates c. 66 cervical vertebrae present, with the possibility that there may have been as many as 68 originally. This allows us to reject close relationships with taxa such as Elasmosaurus platyurus Cope, Citation1869 and Albertonectes vanderveldei Kubo et al. Citation2012, which are characterised by possessing more than 70 cervical vertebrae. All other described forms have fewer cervical vertebrae than CM Zfr 115.
In addition, CM Zfr 115 does not possess elongated centra in the mid-cervical region (the so-called can-shaped centra of Otero (Citation2016)) like those present in E. platyurus, Albertonectes vanderveldei, Terminonatator pointeixensis Sato, Citation2003, Styxosaurus browni Welles, Citation1943 (AMNH 5385) and Styxosaurus sp. (AMNH 1495). Therefore, all these taxa can be distinguished from CM Zfr 115.
CM Zfr 115, and the holotype of T. keyesi, have a posteriorly flat squamosal arch, while the squamosal shaft is dorsoventrally thicker than axially long. This contrasts with the squamosal arches of Thalassomedon haningtoni Welles, Citation1943 and Styxosaurus spp., which have an anteriorly directed embayment in dorsal view (Welles Citation1943, Citation1952; Carpenter Citation1999). Interestingly, a straight squamosal arch (in dorsal view) is also present in C. colombiensis Welles, Citation1962 and in H. alexandrae Welles, Citation1943. However, the anterior cervical vertebrae of C. colombiensis are very different from those of CM Zfr 115 in having a near circular articular facet without a ventral notch (Welles Citation1962: plate 3c). Also, the radius and ulna are longer than wide in the Albian C. colombiensis (Welles Citation1962) whereas those of CM Zfr 115 are wider than long. H. alexandrae differs from CM Zfr 115 in having fewer cervical vertebrae and it shares with CM Zfr 115 and with the T. keyesi holotype the formation of a dorsal triangular prominence at the squamosal midline contact (Welles Citation1962: figure 4; this study: Figure 12). Although these taxa could be closely related, they are different.
The discussion above shows that the closest affinities of CM Zfr 115 are to be found with the holotype of T. keyesi. Overlapping of the skull bones of T. keyesi and CM Zfr 115 () is restricted to only a few portions of the posterior braincase and the mandibles. Even though these do not preserve reliable diagnostic elements among elasmosaurids, there are obvious very close morphological similarities between the two specimens. The shape of the squamosals and the exoccipital-opisthotics are very alike, as are the anterior cervical vertebrae; the minor differences that do occur probably reflect the different ontogenetic stage of each specimen. All these facts strongly suggest that they could belong to closely related taxa, probably the same genus, if not the very same species.
The question arises as to how comparable in age Tuarangisaurus and CM Zfr 115 might be. We know that the latter can be dated to the upper Isabelidinium pellucidum zone of Wilson et al. (Citation2005) but, unfortunately, Tuarangisaurus has not been dated directly. However, Young and Hannah (Citation2010) studied the dinoflagellate biostratigraphy of the Maungataniwha Sandstone of northwest Hawke’s Bay, New Zealand, the lithological unit from which Tuarangisaurus was derived. They concluded that the plesiosaurian-bearing calcareous concretions from this unit exhibited a range of ages, from lower to upper Haumurian (early Campanian to early Maastrichtian), spanning the entire age range of the formation, i.e. from the Satyrodinium haumuriense (Vozzhennikovia spinulosa subzone)–I. pellucidum zones. It is therefore quite feasible that Tuarangisaurus and CM Zfr 115 are about the same age but, equally likely, Tuarangisaurus may be slightly older than CM Zfr 115.
Conclusions
For well over a century, the species Mauisaurus haasti has been considered a very strange austral elasmosaurid with obscure affinities due to the incompleteness of its lectotype (DM R1529). Additional specimens, later referred to the same species, are currently shown to belong to taxa different from DM R1529. As a consequence, the name Mauisaurus haasti Hector should be applied only to the lectotype of the species, DM R1529, and it should be regarded as a nomen dubium as the species cannot be adequately diagnosed, although it does show possible affinities with the Subfamily Aristonectinae. Thus, it is recommended here that further usage of the genus and species M. haasti be abandoned.
Specimen CM Zfr 115, a typical long-necked elasmosaurid at one time believed to belong in Mauisaurus (Hiller et al. Citation2005), can now be regarded as closely comparable to T. keyesi. CM Zfr 115 and the holotype of T. keyesi are shown to be very similar in many respects and both specimens share a common biogeographic and stratigraphic occurrence. This leads us to conclude that CM Zfr 115 can be placed in Tuarangisaurus. However, the former lacks some important elements of the latter, so we prefer to leave it in open nomenclature.
Acknowledgements
The authors are grateful to the directors and curatorial staff of the following New Zealand institutions for allowing access to Mauisaurus specimens in their care: Museum of New Zealand Te Papa Tongarewa, Wellington; Canterbury Museum, Christchurch; GNS Science, Lower Hutt. We are also grateful to anonymous reviewers whose comments prompted us to substantially improve the manuscript. Associate editor: Professor Kathy Campbell.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Benson RBJ, Druckenmiller PS. 2014. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biological Reviews. 89:1–23. doi: 10.1111/brv.12038
- de Blainville HD. 1835. Description de quelques espèces de reptiles de la Californie, précédée de l’analyse d’un système géneral d’erpétologie et d’amphibologie. Nouvelles Annales du Muséum d’Histoire Naturelle, Paris. (série 3). 4:233–269.
- Brown DS. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of the British Museum (Natural History), Geology Series. 35:253–347.
- Carpenter K. 1999. Revision of North American elasmosaurs from the Cretaceous of the Western Interior. Paludicola. 2:148–173.
- Chatterjee S, Small BJ. 1989. New plesiosaurs from the Upper Cretaceous of Antarctica. Geological Society, London, Special Publications. 47:197–215. doi: 10.1144/GSL.SP.1989.047.01.15
- Colbert E. 1949. A new Cretaceous plesiosaur from Venezuela. American Museum Novitates. 1420:1–22.
- Cope ED. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society. 14:1–252. doi: 10.2307/1005355
- Cox GJ. 1991. Prehistoric animals of New Zealand. Auckland: Collins.
- Cruickshank ARI, Fordyce RE. 2002. A new marine reptile (Sauropterygia) from New Zealand: Further evidence for a Late Cretaceous Austral radiation of cryptoclidid plesiosaurs. Palaeontology. 45:557–575. doi: 10.1111/1475-4983.00249
- Fostowicz-Frelik Ł, Gaździcki A. 2001. Anatomy and histology of plesiosaur bones from the Late Cretaceous of Seymour Island, Antarctic Peninsula. In: Gaździcki A, editor. Palaeontological Results of the Polish Antarctic Expeditions. Part III. Palaeontologia Polonica. 60:7–32.
- Gasparini Z, del Valle R, Goñi R. 1984. Un elasmosáurido (Reptilia, Plesiosauria) del Cretácico Superior de la Antárdida. Instituto Antárdico Argentino, Contribución. 305:1–24.
- Gasparini Z, Salgado L, Casadío S. 2003. Maastrichtian plesiosaurs from northern Patagonia. Cretaceous Research. 24:157–170. doi: 10.1016/S0195-6671(03)00036-3
- Gasparini Z, Salgado L, Parras A. 2007. Late Cretaceous plesiosaurs from northern Patagonia, Argentina. Geological Journal. 42:185–202. doi: 10.1002/gj.1079
- Gray JE. 1825. A synopsis of the genera of reptiles and Amphibia, with a description of some new species. Annals of Philosophy. 26:193–217.
- Hector J. 1874. On the Fossil Reptilia of New Zealand. Transactions and Proceedings of the New Zealand Institute. 6:333–358.
- Hiller N, Mannering AA. 2005. An unusual new elasmosaurid plesiosaur (Sauropterygia) from the Upper Haumurian (Maastrichtian) of the South Island, New Zealand. Memoirs of the Queensland Museum. 51:27–37.
- Hiller N, Mannering AA, Jones CM, Cruickshank ARI. 2005. The nature of Mauisaurus haasti Hector, 1874 (Reptilia:Plesiosauria). Journal of Vertebrate Paleontology. 25:588–601. doi: 10.1671/0272-4634(2005)025[0588:TNOMHH]2.0.CO;2
- Hutton FW. 1894. On a new plesiosaur from the Waipara River, New Zealand. Transactions and Proceedings of the New Zealand Institute. 26:354–358.
- Knight C. 1874. On the teeth of the Leiodon. Transactions and Proceedings of the New Zealand Institute. 6:358–364.
- Kubo T, Mitchell MT, Henderson DM. 2012. Albertonectes vanderveldei, a new elasmosaur (Reptilia, Sauropterygia) from the Upper Cretaceous of Alberta. Journal of Vertebrate Paleontology. 32:557–572. doi: 10.1080/02724634.2012.658124
- Long JA. 1998. The Dinosaurs of Australia and New Zealand, and other animals of the Mesozoic Era. Sydney: UNSW Press.
- Lydekker R. 1889. Catalogue of the fossil Reptilia and Amphibia in the British Museum. Part 2. Orders Ichthyopterygia and Sauropterygia. London: British Museum. p. 307.
- Martin JE, Sawyer JF, Reguero M, Case JA. 2007. Occurrence of a young elasmosaurid plesiosaur skeleton from the Late Cretaceous (Maastrichtian) of Antarctica. In: Cooper AK, Raymond CR et al. editors. Antarctica: A Keystone in a Changing World—Online Proceedings of the 10th ISAES. USGS Open-File Report 2007–1047, Short Research Paper 066:1–4.
- O’Gorman JP. 2013. Plesiosauricos del Cretácico Superior de Patagonia y Penísula Antártica. La Plata (AR): National University of La Plata.
- O’Gorman JP. 2016. A small body sized non-aristonectine elasmosaurid (Sauropterygia, Plesiosauria) from the Late Cretaceous of Patagonia with comments on the relationships of the Patagonian and Antarctic elasmosaurids. Ameghiniana. 53:245–268. doi: 10.5710/AMGH.29.11.2015.2928
- O’Gorman JP, Gasparini Z, Salgado L. 2013. Postcranial morphology of Aristonectes (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of Patagonia and Antarctica. Antarctic Science. 25:71–82. doi: 10.1017/S0954102012000673
- O’Gorman JP, Gasparini Z, Salgado L. 2014a. Reappraisal of Tuarangisaurus? cabazai (Elasmosauridae, Plesiosauria) from the Upper Maastrichtian of northern Patagonia, Argentina. Cretaceous Research. 47:39–47. doi: 10.1016/j.cretres.2013.10.003
- O’Gorman JP, Otero RA, Hiller N. 2014b. A new record of an aristonectine elasmosaurid (Sauropterygia, Plesiosauria) from the Upper Cretaceous of New Zealand: Implications for the Mauisaurus haasti Hector, 1874 hypodigm. Alcheringa: An Australasian Journal of Palaeontology. 38:504–512. doi: 10.1080/03115518.2014.908267
- O’Gorman JP, Otero RA, Hiller N, Simes J, Terezow M. (2017). Redescription of Tuarangisaurus keyesi (Sauropterygia; Elasmosauridae), a key species from the uppermost Cretaceous of the Weddellian Province – internal skull anatomy and phylogenetic position. Cretaceous Research. 71:118–136. doi: 10.1016/j.cretres.2016.11.014
- O’Gorman JP, Salgado L, Olivero EB, Marenssi SA. 2015. Vegasaurus molyi, gen. et sp. nov. (Plesiosauria, Elasmosauridae), from the Cape Lamb Member (Lower Maastrichtian) of the Snow Hill Island Formation, Vega Island, Antarctica, and remarks on Weddellian Elasmosauridae. Journal of Vertebrate Paleontology. 35:e931285. doi:10.1080/02724634.2014.931285.
- O’Keefe FR. 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica. 213:1–63.
- O’Keefe FR, Hiller N. 2006. Morphologic and ontogenetic patterns in elasmosaur neck length, with comments on the taxonomic utility of neck length variables. Paludicola. 5:206–229.
- Otero RA. 2016. Taxonomic reassessment of Hydralmosaurus as Styxosaurus: New insights on the elasmosaurid neck evolution throughout the Cretaceous. PeerJ. 4:e1777. doi:10.7717/peerj.1777.
- Otero RA, O’Gorman JP, Hiller N. 2015a. Reassessment of the late Maastrichtian specimens from central Chile referred to the genus Mauisaurus Hector, 1874 (Plesiosauroidea, Elasmosauridae) with comments about the hemispherical femoral head among aristonectines. New Zealand Journal of Geology and Geophysics. 58:252–261. doi: 10.1080/00288306.2015.1037775
- Otero RA, Soto-Acuña S, O’Keefe FR, O’Gorman JP, Stinnesbeck W, Suárez ME, Rubilar-Rogers D, Salazar C, Quinzio-Sinn LA. 2014. Aristonectes quiriquinensis, sp. nov., a new highly derived elasmosaurid from the upper Maastrichtian of central Chile. Journal of Vertebrate Paleontology. 34:100–125. doi: 10.1080/02724634.2013.780953
- Otero RA, Soto-Acuña S, Rubilar-Rogers D. 2010. Presence of the elasmosaurid plesiosaur Mauisaurus in the Maastrichtian (Late Cretaceous) of central Chile. Acta Palaeontologia Polonica. 55:361–364.
- Otero RA, Soto-Acuña S, Rubilar-Rogers D. 2012. A postcranial skeleton of an elasmosaurid plesiosaur from the Maastrichtian of central Chile, with comments on the affinities of Late Cretaceous plesiosauroids from the Weddellian Biogeographic Province. Cretaceous Research. 37:89–99. doi: 10.1016/j.cretres.2012.03.010
- Otero RA, Soto-Acuña S, Salazar C, Oyarzún JL. 2015b. New elasmosaurids (Sauropterygia, Plesiosauria) from the Late Cretaceous of the Magallanes Basin, Chilean Patagonia: Evidence of a faunal turnover during the Maastrichtian along the Weddellian Biogeographic Province. Andean Geology. 42:237–267.
- Owen R. 1860. On the orders of fossil and Recent Reptilia, and their distribution in time. Report of the British Association for the Advancement of Science for 1859. 29:153–166.
- Roncaglia L, Field BD, Raine JI, Schiøler P, Wilson GJ. 1999. Dinoflagellate biostratigraphy of Piripauan–Haumurian (Upper Cretaceous) sections from northeast South Island, New Zealand. Cretaceous Research. 20:271–314. doi: 10.1006/cres.1999.0153
- Sachs S. 2004. Redescription of Woolungasaurus glendowerensis (Plesiosauria: Elasmosauridae) from the Lower Cretaceous of Northeast Queensland. Memoirs of the Queensland Museum. 49:713–731.
- Sato T. 2003. Terimonatator ponteixensis, a new elasmosaur (Reptilia; Sauropterygia) from the Upper Cretaceous of Saskatchewan. Journal of Vertebrate Paleontology. 23:89–103. doi: 10.1671/0272-4634(2003)23[89:TPANES]2.0.CO;2
- Sato T, Hasegawa Y, Manabe M. 2006. A new elasmosaurid plesiosaur from the Upper Cretaceous of Fukushima, Japan. Palaeontology. 49:467–484. doi: 10.1111/j.1475-4983.2006.00554.x
- Seeley HG. 1877. On Mauisaurus gardneri (Seeley), an elasmosaurian from the base of the Gault at Folkstone. Quarterly Journal of the Geological Society. 33:541–547. doi: 10.1144/GSL.JGS.1877.033.01-04.32
- Welles SP. 1943. Elasmosaurid plesiosaurs with a description of new material from California and Colorada. Memoirs of the University of California. 13:125–215.
- Welles SP. 1952. A review of the North American Cretaceous elasmosaurs. University of California Publications in Geological Sciences. 29:46–144.
- Welles SP. 1962. A new species of Elasmosaur from the Aptian of Colombia and a review of the Cretaceous plesiosaurs. University of California, Publications in Geological Sciences. 44:1–96.
- Welles S, Gregg DR. 1971. Late Cretaceous marine reptiles of New Zealand. Records of the Canterbury Museum. 9:1–111.
- Wiffen J, Moisley WL. 1986. Late Cretaceous reptiles (Families Elasmosauridae and Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand. New Zealand Journal of Geology and Geophysics. 29:205–252. doi: 10.1080/00288306.1986.10427535
- Wilson GJ, Schiøler P, Hiller N, Jones CM. 2005. Age and provenance of Cretaceous marine reptiles from the South Island and Chatham Islands, New Zealand. New Zealand Journal of Geology and Geophysics. 48:377–387. doi: 10.1080/00288306.2005.9515120
- Young MD, Hannah MJ. 2010. Dinoflagellate biostratigraphy of the vertebrate fossil-bearing Maungataniwha Sandstone, northwest Hawke’s Bay, New Zealand. New Zealand Journal of Geology and Geophysics. 53:81–87. doi: 10.1080/00288301003639742
- Zinsmeister WJ. 1979. Biogeographic significance of the Late Mesozoic and early Tertiary molluscan faunas of Seymour Island (Antarctic Peninsula) to the final break-up of Gondwanaland. In: Gray J, Boucot AJ. editors. Historical biogeography, plate tectonics and the changing environment. Corvallis: Oregon State University Press; p. 349–355.