
Abstract
We examined the effects on offspring of ingestion of the 1975 Japanese diet during pregnancy and lactation and after weaning in mice. Pregnant dams were divided into groups that were fed the Japanese diet or a control diet and raised until offspring were weaned. The offspring after weaning were further divided into groups that were raised on the Japanese diet or the control diet. Ingestion of the Japanese diet after weaning suppressed accumulation of visceral fat in offspring, and reduced the amount of lipids in serum and liver. This effect was weakened if the Japanese diet was only ingested during pregnancy and lactation. Therefore, it was suggested that ingestion of the Japanese diet of mothers during pregnancy and lactation weakens the lipid accumulation inhibitory effect of the Japanese diet in children.
Ingestion of the Japanese diet of mothers during pregnancy and lactation weakens the lipid accumulation inhibitory effect of the Japanese diet in children.
Prevention of obesity and metabolic syndrome has become a global problem, and there is an increased understanding of the importance of prevention of causes of obesity, such as excessive energy intake and lack of exercise. In recent years, the DOHaD (Developmental Origins of Health and Disease) theory that a mother’s malnutrition or overnutrition in pregnancy and lactation may lead to onset of metabolic syndrome in offspring after growth has been proposed, and several studies have shown that maternal dietary habits influence the risk of obesity in offspring [Citation1–3]. For example, obesity and fatty liver occur at a higher rate in offspring from obese dams after the offspring ingest a high-fat diet [Citation4,5], and a mother’s high fat diet changes sugar metabolism in offspring and increases the risk of development of insulin resistance [Citation6]. In rats, application of caloric restriction to mothers during lactation causes malnutrition in offspring and an increased risk of obesity and insulin resistance after growth [Citation7,8]. Thus, it has become clear that the diet ingested by a mother can have a major influence on the future health of offspring.
In our previous study, which was based on the marked changes in the Japanese diet over the past 50 years, we created one-week menus of the 1960, 1975, 1990 and 2005 Japanese diets using National Health and Nutrition Survey (NHNS) data. The meals for each period were prepared, freeze-dried, homogenized by grinding and stirring, and fed to mice. The results showed that the 1975 diet reduced obesity and had high health benefits [Citation9] However, in a study examining the effects of dietary intake during lactation on offspring, in which weekly menus from the 1960, 1975, 1990 and 2005 diets were prepared as powdered foods and given to dams during lactation, the 1975 diet promoted fat accumulation in the liver of offspring that were fed a high fat diet after weaning [Citation10]. This may have occurred because the ingredients of the 1975 diet given to the dam adversely affected offspring or because the large energy difference between the high fat diet after weaning and the 1975 diet had a bad effect on offspring.
In this study, we investigated whether the 1975 Japanese diet fed to dams had a bad effect on offspring by feeding the offspring after weaning with a control diet, rather than from a high-fat diet, to eliminate the possible effect of an energy difference. In addition, children are greatly affected by the nutritional status in pregnancy, as well as during lactation. Therefore, in this study, we focused on the pregnancy and lactation period and after weaning, and fed dams with the 1975 Japanese diet or control diet in these two stages to examine effects on offspring. Our results show that ingestion of the 1975 diet after weaning suppresses fat accumulation in offspring, but that this effect is weakened when the same diet is ingested during pregnancy and lactation only.
Materials and methods
Preparation of test diets
To understand the effect on offspring of intake of the 1975 Japanese diet during pregnancy and lactation, the diet was prepared as previously reported [Citation10,11]. We created weekly menus (for 21 meals) for the 1975 year based on NHNS, under the guidance of a registered dietitian (Table ) [Citation12,13]. The 1975 Japanese diet based on the menu were prepared, freeze-dried in a vacuum freeze dryer (FD-550 R; Tokyo Rikakikai, Japan), and homogenized by grinding and stirring. The nutritional composition (protein, fat, carbohydrate, moisture, ash, and energy) of the meals was determined as follows: protein, by a modified Dumas method; fat, by acid degradation; moisture, by vacuum oven drying; ash, by direct ashing; and carbohydrate, by subtracting the fat, protein, moisture, and ash contents from the total. The energy content was calculated by applying modified Atwater factors (4, 9, and 4 kcal/g for protein, fat, and carbohydrate, respectively) [Citation11,14]. To avoid any influences of differences in the mouse diet properties (such as stickiness), the Japanese diet (the test diet) comprised the control diet (98121701; Research Diets Inc., New Brunswick, NJ, USA) (Supplemental Table 1 and 2) supplemented with 30% of the 1975 Japanese diet (Table , Supplemental Table 2) by weight. Since the control diet is a meal designed for breeding the mouse normally, the control diet in this study used as a control to evaluate the phenomenon appearing in the body of the mouse by the Japanese diet.
Table 1. Menu card of 1975 Japanese diet.
Animals
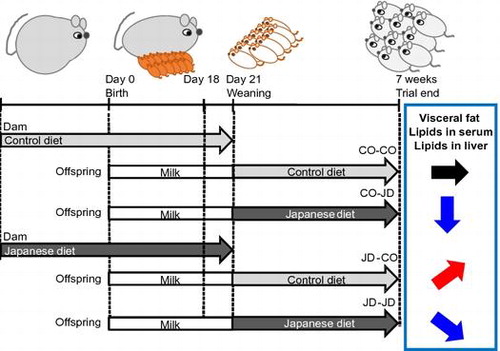
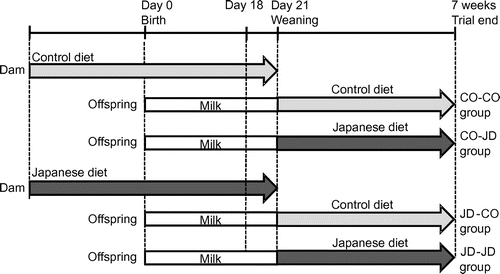
All procedures were performed in accordance with the Animal Experiment Guidelines of Tohoku University, and the animal protocol was approved by the Animal Use Committee at Tohoku University. Female ICR mice purchased on the fourth day of pregnancy (Japan CLEA, Tokyo) were maintained in a room at constant temperature (24 ± 1 °C) and humidity with light cycles of 12 h on and 12 h off in individual cages. Dams were randomly divided into two groups with free access to the control diet (CO group) or the prepared Japanese diet (JD group) and to water. In Experiment 1, offspring of each group were weaned at 3 weeks, and only male offspring were divided into four groups and raised for 4 weeks as follows: dams and offspring both fed with the control diet (CO-CO group); dams fed the control diet and offspring fed the Japanese diet (CO-JD group); dams fed the Japanese diet and offspring fed the control diet (JD-CO group); and dams and offspring both fed the Japanese diet (JD-JD group) (Figure ). After weaning, the food intake and body weight of offspring were measured every 3 days and 1 week, respectively. As in our previous study, offspring were weighed, anesthetized by aspirating isoflurane, and sacrificed at 7 weeks after 12 h fasting, and blood samples, brain, liver, kidneys, pancreas and white adipose tissue were collected [Citation15]. Blood was allowed to stand at room temperature for 20 minutes, then stored at 4 °C for 3 hours and centrifuged (900 × g, 5 °C, 15 min) to prepare serum. Serum and organs were stored at −80 °C until assays were performed. In Experiment 2, to investigate the influence of the mother’s 1975 Japanese diet intake during pregnancy and lactation on infancy offspring, sacrifice and dissection were performed after fasting for 12 h at 18 days of age before weaning, and blood samples, liver and white adipose tissue were collected. To reduce differences between dams, one offspring of average weight per dam was used. In addition, to match sex with experiment 1, only male offspring samples were prepared and stored in the same manner until use.
Figure 1. Study protocol. Dams were given the Control diet or the Japanese diet during pregnancy and lactation and offspring from each group was divided into two groups given the Control diet or 1975 Japanese diet after weaning at 3 weeks.
Biochemical analyses in serum and liver
Lipid levels in serum and liver were measured as described previously [Citation16]. Triacylglycerol (TG) and total cholesterol (TC) in serum and liver, and phospholipid (PL) and glucose in serum were measured using commercial enzyme kits (Wako Pure Chemical, Osaka, Japan). Insulin in serum was determined using an ELISA kit (Morinaga Institute of Biological Science, Kanagawa, Japan). Albumin in serum was determined using an ELISA kit (Shibayagi, Gunma, Japan). PL in liver was determined using the method described by Rouser [Citation17]. Since the amount of serum was small in 18-day-old mice, serum glucose and insulin were measured by mixing all samples from each group in equal amounts.
Histological analysis
Histological analysis of liver and white adipose tissue were observed as described previously [Citation16,18]. Liver and epididymal adipose tissue were fixed in 10% formalin and embedded in paraffin. Vertical sections (5 μm) were cut, mounted on a glass slide, stained with hematoxylin and eosin, and observed using a microscope (BZ-9000; Keyence, Osaka, Japan). The mean area of adipocytes in each group was calculated [Citation17,19].
mRNA expression analysis
For real-time quantitative reverse transcriptase PCR (qRT-PCR), total RNA was isolated from liver using an RNeasy Mini Kit (Qiagen, Valencia, CA, USA) [Citation16,18] and stored at −80 °C until assays were performed. cDNA was made using a PrimeScript® RT Master Mix (Takara Bio, Otsu, Japan) and mRNA levels were determined using a Thermal Cycler Dice Real Time System (Takara Bio, Otsu, Japan) to quantify expression levels of genes [Citation16,18]. Primers with sequences based on past reports were prepared by Sigma-Aldrich (Tokyo, Japan) (Table ) [Citation19–21]. The ratio between the β-actin content in standard and test samples was defined as the normalization factor.
Table 2. Primer pairs used for the quantitative RT-PCR analysis.
Statistical analysis
Results are expressed as the mean ± standard error (SE). For Experiment 1, because there were two factors of maternal diet and post-weaning diet, data were analyzed using two-way analysis of variance (ANOVA). For items with significant differences, one-way ANOVA was performed, followed by a Tukey-Kramer test for multiple comparisons among 4 groups. For Experiment 2, data were analyzed by Student t test for multiple comparisons among 2 groups. A difference was considered significant at p < 0.05.
Results
Composition of the test diets
Nutritional compositions of test diets are given in Table . Protein and fat were higher and carbohydrate was lower in the Japanese diet than in the control diet. The energy per 100 g of diet was 11 kcal/100 g higher in the Japanese diet.
Table 3. Nutritional compositions of test diets.
The number and the ratio of male to female of offspring per dam
The number of offspring per dam was 13.1 ± 1.1 (n = 6) in dam fed the control diet and 13.9 ± 1.2 mice (n = 8) in dam fed the Japanese diet. There were no significant effects of the maternal diet on the number of offspring per dam. The ratio of male to female of offspring per dam (male / female) was 1.16 ± 0.27 (n = 6) in dam fed the control diet and 1.07 ± 0.22 mice (n = 8) in dam fed the Japanese diet. There were no significant effects of the maternal diet on the ratio of male to female of offspring per dam.
Growth parameters
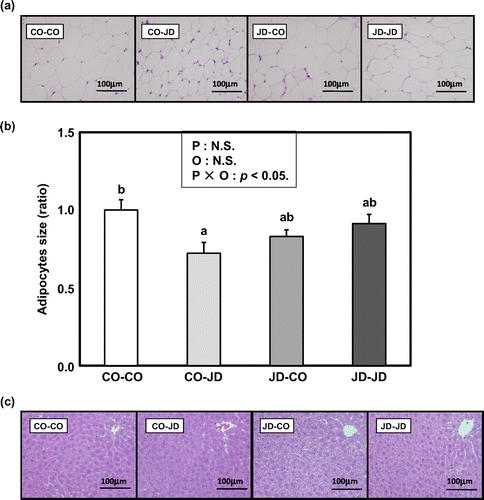
There were no significant effects of the maternal diet or post-weaning diet on initial and final body weight, body weight gain, food intake, and energy intake among the CO-CO, CO-JD, JD-CO and JD-JD groups (Table ). There were also no significant effects on the weights of brain, liver and pancreas, but the maternal diet and post-weaning diet did significantly affect the weight of the kidney. There was no significant difference in kidney weight between the CO-JD and CO-CO groups, but a tendency for this weight to be higher in the CO-JD group compared to the CO-CO group and to be lower in the JD-JD group compared to the CO-JD group. Significant effects of the post-weaning diet were seen for the mesenteric, perinephric and total white adipose tissue weight. The white adipose tissue weight was significantly lower in the CO-JD group than in the CO-CO group, and higher in the JD-JD group than in the CO-JD group (not significant, NS). Effects of the maternal diet and post-weaning diet were also seen for the epididymal adipose tissue weight, which was significantly lower in the CO-JD group than in the CO-CO group and showed a tendency to be higher in the JD-JD group compared to the CO-JD group. Since changes in adipose tissue weight were observed, epididymal adipose tissues were stained with hematoxylin & eosin (H&E) and subjected to histological analysis. Adipocytes were smaller in the CO-JD group than in the CO-CO group, and this phenomenon was weakened in the JD-JD group (Figure a). The area of adipocytes in each group indicated a significant effect of the interaction between maternal diet and post-weaning diet. The area of adipocytes was significantly reduced in the CO-JD group compared to the CO-CO group, and showed a tendency to be larger in the JD-JD group than in the CO-JD group (Figure b). These results show that ingestion of the Japanese diet after weaning suppressed accumulation of visceral fat in offspring, and that this suppressive effect was weakened by ingestion of the Japanese diet during pregnancy and lactation.
Table 4. Growth parameters.
Figure 2. Effects of the Japanese diet intake from pregnancy and lactation on adipocytes and liver of offspring at 7 weeks. (a) Effects of the Japanese diet intake from pregnancy and lactation on adipocytes of offspring at 7 weeks. Photographs were hematoxylin-eosin staining of epididymal adipose tissue sections from representative mice of each group (scale bar = 100 μm). (b) Bar graph of the adipocytes size ratio of the CO-CO, CO-JD, JD-CO and JD-JD groups. Statistical analysis was performed using one-way ANOVA and two-way ANOVA followed by Tukey-Kramer’s test. Different superscript letters indicate significantly different means at p < 0.05. Maternal diet effect, Post-weaning diet effect and interaction were described at the top of the graph: P, Maternal diet effect; O, post-weaning diet effect; P × O, interaction. (c) Effects of the Japanese diet intake from pregnancy and lactation on liver of offspring at 7 weeks. Photographs were hematoxylin-eosin staining of liver sections from representative mice of each group (scale bar = 100 μm).
Biochemical parameters in serum
The maternal diet and post-weaning diet had significant effects on the serum TG level (Table ). TG was significantly lower in the CO-JD group than in the CO-CO group, and higher in the JD-JD group than in the CO-JD group (NS). A significant effect of the maternal diet and the interaction was seen for serum TC, which was lower in the CO-JD group than in the CO-CO group (NS) and significantly higher in the JD-JD group than in the CO-JD group. A significant effect of the interaction was seen for serum PL, which was lower in the CO-JD group than in the CO-CO group (NS) and significantly higher in the JD-JD group compared to the CO-JD group. There was no significant effect on serum glucose. The post-weaning diet had a significant effect on serum insulin, but there were no significant differences among the CO-CO, CO-JD, JD-CO and JD-JD groups. There was no significant effect on serum albumin. These results show that ingestion of the Japanese diet after weaning reduced the TG level in serum of offspring, and that this suppressive effect was weakened by ingestion of this diet during pregnancy and lactation.
Table 5. Biochemical parameters.
Biochemical parameters in liver
The post-weaning diet influenced TG and TC levels in liver (Table ). Liver TG was significantly lower in the CO-JD group than in the CO-CO group and higher in the JD-JD group than in the CO-JD group (NS). Liver TC was higher in the CO-JD group than in the CO-CO group (NS) and lower in the JD-JD group than in the CO-JD group (NS). There were no significant effects on PL in liver. To confirm the change in liver TG, histological analysis was performed by staining liver tissue with H&E (Figure c). There was little lipid accumulation in hepatocytes in the CO-JD and JD-JD groups. These results show that ingestion of the Japanese diet after weaning suppresses lipid accumulation in the liver of offspring, and that this effect is weakened by ingestion of this diet during pregnancy and lactation.
mRNA expression analysis in the liver
To examine changes in lipid content in white adipose tissue and liver in detail, the expression levels of lipid metabolism-related genes in liver, which plays a central role in lipid metabolism, were determined by qRT-PCR. The post-weaning diet had a significant effect on the mRNA level of Acc, which promotes fatty acid synthesis (Table ). The Acc level was significantly lower in the CO-JD group than in the CO-CO group, and there was no significant difference between the JD-JD and CO-JD groups. The maternal diet had significant effects on the mRNA levels of Fasn, G6pdx, Me, and the post-weaning diet had a significant effect on the mRNA level of Srebp1c. All of these proteins promote fatty acid synthesis. However, there were no significant differences in the levels of these mRNAs among the CO-CO, CO-JD, JD-CO and JD-JD groups. The post-weaning diet had significant effects on the mRNA levels of Acox1, Cpt1, Cpt2 and Ppara, which are involved in fatty acid β oxidation. The mRNA levels of Acox1, Cpt1 and Cpt2 were lower in the CO-JD group than in the CO-CO group (NS) and higher in the JD-JD group than in the CO-JD group (NS). In addition, the mRNA level of Ppara was significantly lower in the CO-JD group than in the CO-CO group, and higher in the JD-JD group than in the CO-JD group (NS). The maternal diet had a significant effect on the mRNA level of Hmgcoar, which is involved in cholesterol synthesis: this level was higher in the CO-JD group than in the CO-CO group (NS) and in the JD-JD group compared in the CO-JD group (NS). The maternal diet and post-weaning diet had significant effects on the mRNA level of Sqle, which is also involved in cholesterol synthesis: this level was lower in the CO-JD group than in the CO-CO group (NS) and higher in the JD-JD group compared to the CO-JD group (NS). The post-weaning diet had a significant effect on the mRNA level of Abcg5, which is related to cholesterol exhaust: this level was significantly higher in the CO-JD group than in the CO-CO group, and lower in the JD-JD group compared to the CO-JD group (NS). There was no significant effect on the mRNA levels of Acat3 and Cyp7a1. The post-weaning diet and interaction had a significant effect on the mRNA level of Lxra: this level was significantly lower in the CO-JD group than in the CO-CO group, and higher in the JD-JD group than in the CO-JD group (NS). These results suggest that ingestion of the Japanese diet by offspring after weaning suppresses fatty acid synthesis and oxidation in liver, reduces synthesis of cholesterol, and tends to promote catabolism. These changes were weakened by ingestion of the Japanese diet during pregnancy and lactation.
Table 6. mRNA expression levels for lipid metabolism-related genes in liver.
Growth parameters at 18 days of age
Because Japanese diet intake during pregnancy and lactation weakened the effect of suppression of lipid accumulation in offspring after growth, we examined the body composition of offspring at age 18 days to investigate whether this phenomenon had already appeared in offspring before weaning (Experiment 2). The results showed no significant difference between the CO and JD groups for body weight, liver weight and white adipose tissue weight at 18 days (Table ). Therefore, consumption of the Japanese diet during pregnancy and lactation did not significantly affect the body composition of offspring before weaning.
Table 7. Growth parameters of 18 days offspring.
Biochemical parameters in serum and liver at 18 days of age
TG, TC and PL levels in serum at age 18 days did not differ significantly between the CO and JD groups (Table ). Since the amount of serum was small, glucose and insulin levels were determined in mixed samples with equal amounts from each mouse in each group. The glucose level in serum was lower in the JD group (6.79 mmol/L) than in the CO group (7.27 mmol/L), but the insulin level was 0.06 nmol/L in both groups. TG in liver was significantly lower in the JD group than in the CO group, but TC and PL in liver did not differ significantly between the two groups. These results show that ingestion of the Japanese diet during pregnancy and lactation suppresses lipid accumulation in the liver of offspring before weaning.
Table 8. Biochemical parameters of 18 days offspring.
mRNA expression analysis in the liver at 18 days of age
To investigate the mechanism of changes in lipid contents in liver, expression of lipid metabolism-related genes in liver at 18 days of age was measured by quantitative RT-PCR. The mRNA levels of Acc and Fasn, which are involved in fatty acid synthesis, were significantly lower in the JD group than in the CO group (Table ). There were no significant differences in the mRNA levels of G6pdx, Me and Srebp1c. The mRNA levels of Aco, Cpt1, Cpt2 and Ppara, which are involved in fatty acid β oxidation, also did not differ significantly between the two groups. The mRNA levels of Hmgcoar and Sqle, which are involved in cholesterol synthesis, were significantly lower in the JD group than in the CO group. The mRNA levels of Abcg5, Acat3and Cyp7a1, which are related to cholesterol catabolism, did not differ significantly between the groups, but the mRNA level of Lxra was significantly lower in the JD group. These results show that ingestion of the Japanese diet during pregnancy and lactation suppresses lipid accumulation in the liver by suppressing the synthesis of fatty acids and cholesterol in the liver of offspring before weaning.
Table 9. mRNA expression levels for lipid metabolism-related genes in liver of 18 days offspring.
Discussion
In this study, the 1975 Japanese diet or a control diet were ingested by the mother during pregnancy and lactation, and by offspring after weaning. The results were used to examine the effects of the 1975 Japanese diet intake during these periods on lipid metabolism in offspring. Ingestion of the Japanese diet after weaning suppressed fat accumulation in offspring, and this effect was weakened by intake of the Japanese diet during pregnancy and lactation. Ingestion of the Japanese diet during pregnancy and lactation suppressed lipid accumulation in the liver of offspring before weaning. The Japanese diet used in this study was prepared by freeze-drying and powdering of dishes that reproduced a 21-meal menu for one week based on NHNS data for 1975, as in our previous studies [Citation10,11]. The characteristics of the 1975 Japanese diet include high levels of soy products such as tofu and fried tofu, fruits and seaweed. Our recent study clearly showed that the beneficial effect of the 1975 Japanese diet on health of adult mice does not depend on the PFC balance [Citation11]. Therefore, the results in the current study are due to differences in ingredients included in the 1975 Japanese diet.
In 7-week-old offspring, white adipose tissue weight and TG levels in serum and liver were significantly reduced by intake of the Japanese diet after weaning. This confirms that the Japanese diet has a suppressive effect on lipid accumulation, as found previously [Citation11]. However, these phenomena were all weakened by ingestion of the Japanese diet during pregnancy and lactation. This tendency was also observed in H&E staining of white adipose tissue. In the previous study, the adverse effect of lipid accumulation in the liver of offspring after ingestion of the 1975 Japanese diet during lactation could have been due to the large energy difference between the high fat diet given to the offspring and the 1975 Japanese diet [Citation10]. Therefore, in the current study, a control diet was fed to offspring to eliminate the possibility of an adverse affect due to the energy difference. Therefore, the current results suggest that ingestion of the Japanese diet during pregnancy and lactation had an influence on the fetus or infant and weakened suppression of lipid accumulation in offspring after growth. Interestingly, analysis of 18-day-old offspring before weaning showed that TG in liver was significantly lower if mothers ingested the Japanese diet. This may be due to the caloric restriction-like effect of the 1975 Japanese diet affecting growth of the fetus and infant. Several studies have shown that caloric restriction during pregnancy and lactation leads to malnutrition of offspring and restricted growth in the uterus, and affects processes such as lipid metabolism in offspring after birth [Citation7,22–25]. Therefore, the Japanese diet strongly inhibited lipid accumulation in the liver of fetuses and infant, and caused the offspring to be in a state of lipid deficiency that affected their lipid metabolism system after growth.
Effects of the Japanese diet during pregnancy and lactation and/or post-weaning on the lipid metabolism system of offspring were also observed at the genetic level. In 7-week-old offspring, mRNA levels of Acc [Citation26] (fatty acid synthesis) and Acox1 [Citation27], Cpt1 [Citation28], Cpt2 [Citation29] and Ppara [Citation30] (fatty acid β oxidation) in the liver were low in offspring that ingested the Japanese diet after weaning. Therefore, intake of the Japanese diet after weaning suppresses lipid accumulation in the liver by suppressing the liver fatty acid synthesis system, and the fatty acid β-oxidation system in the liver is then suppressed because of the low lipid level. The mRNA level of Hmgcoar [Citation31] (cholesterol synthesis) tended to be higher in offspring that ingested the Japanese diet after weaning, but that of Sqle [Citation32] located downstream tended to be lower. The mRNA level of Abcg5 [Citation33] (cholesterol emission) was also significantly higher in offspring that ingested the Japanese diet after weaning. Therefore, the Japanese diet after weaning suppresses cholesterol synthesis in liver and promotes catabolism, thereby suppressing cholesterol accumulation.
Ingesting the Japanese diet during pregnancy and lactation seemed to weaken changes in gene expression induced by intake of the diet after weaning. To clarify whether such changes had already occurred before weaning, pre-weaning (18-day-old) offspring were examined. In these offspring, the mRNA levels of Acc and Fasn (fatty acid synthesis) were significantly lower in the JD group than in the CO group. Therefore, ingestion of the Japanese diet during pregnancy and lactation suppressed lipid accumulation in the liver by inhibiting fatty acid synthesis in infants. In addition, although there was no significant change in the liver TC level at 18 days of age, the mRNA levels of Hmgcoar and Sqle (cholesterol synthesis) and Lxra [Citation34] (cholesterol metabolism) in liver were significantly lower in the JD group compared to the CO group. This suggests that ingestion of the Japanese diet during pregnancy and lactation suppressed metabolism of cholesterol in the liver of infants. These results suggest that intake of the Japanese diet during pregnancy and lactation restricts lipid accumulation of offspring by affecting expression of lipid metabolism-related genes in the liver before weaning, which causes offspring to be in a state of lipid deficiency. Thus, when these offspring can take food themselves after weaning, their constitution has changed so that more lipid can be absorbed than usual, and the suppressive effect on lipid accumulation of the Japanese diet is weakened. Catch-up growth after weaning of low birth weight infants reported in several studies may occur through a similar phenomenon [Citation35,36].
This study showed that ingestion of the Japanese diet during pregnancy and lactation weakened the suppressive effect of the Japanese diet on lipid accumulation after weaning in offspring. This suggests that ingesting the 1975 Japanese diet during weaning without intake of the same diet during pregnancy and lactation gives a higher health benefit. In addition, ingestion of the Japanese diet during pregnancy and lactation did not adversely affect growth of offspring, except for weakening of the effect of the diet after weaning. Actually, the Japanese diet intake of dams did not affect the number and the ratio of male to female of offspring per dam, and serum albumin level in its offspring was not significantly affected. Food intake by offspring after weaning determines their future metabolic state to a greater extent than food eaten by the mother [Citation37], which suggests that the choice of food ingested after weaning is important for the health conditions of the offspring.
Ingestion of the Japanese diet after weaning suppressed accumulation of visceral fat in offspring and reduced the lipid levels in serum and liver. This effect was weakened when the diet was ingested during pregnancy and lactation. Interestingly, ingestion of the Japanese diet during pregnancy and lactation suppressed lipid accumulation in the liver of offspring before weaning.
Author’s contribution
Study concept and design: Tsuyoshi TSUDUKI. Acquisition of data: Haruna ISHIKAWA, Xiaoxu GUO, Saeko SUGAWARA, Yui IWAGAKI, and Kazushi YAMAMOTO. Analysis and interpretation of data: Haruna ISHIKAWA, Xiaoxu GUO, Saeko SUGAWARA, Yui IWAGAKI, Kazushi YAMAMOTO, and Tsuyoshi TSUDUKI. Drafting of the manuscript: Haruna ISHIKAWA, Xiaoxu GUO, and Tsuyoshi TSUDUKI. All authors reviewed and approved the final manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was supported by Project of the NARO Bio-oriented Technology Research Advancement Institution (Advanced integration research for agriculture and interdisciplinary fields); The Canon Foundation.
Supplemental data
Supplemental data for this article can be accessed at https://doi.org/10.1080/09168451.2018.1428788.
S._tables-h.pdf
Download PDF (421.7 KB)Related Research Data
References
- Ashino NG, Saito KN, Souza FD, et al. Maternal high-fat feeding through pregnancy and lactation predisposes mouse offspring to molecular insulin resistance and fatty liver. J Nutr Biochem. 2012;23:341–348.10.1016/j.jnutbio.2010.12.011
- Osmond C, Barker DJ. Fetal, infant, and childhood growth are predictors of coronary heart disease, diabetes, and hypertension in adult men and women. Environ Health Perspect. 2000;108:545–553.10.1289/ehp.00108s3545
- Barouki R, Gluckman PD, Grandjean P, et al. Developmental origins of non-communicable disease: implications for research and public health. Environ Health. 2012;11:42.10.1186/1476-069X-11-42
- Shankar K, Harrell A, Liu X, et al. Maternal obesity at conception programs obesity in the offspring. Am J Physiol Regul Integr Comp Physiol. 2008;294:R528–R538.10.1152/ajpregu.00316.2007
- Elahi MM, Cagampang FR, Mukhtar D, et al. Long-term maternal high-fat feeding from weaning through pregnancy and lactation predisposes offspring to hypertension, raised plasma lipids and fatty liver in mice. Br J Nutr. 2009;102:514–519.10.1017/S000711450820749X
- Samuelsson AM, Matthews PA, Argenton M, et al. Diet-induced obesity in female mice leads to offspring hyperphagia, adiposity, hypertension, and insulin resistance: A novel murine model of developmental programming. Hypertension. 2008;51:383–392.10.1161/HYPERTENSIONAHA.107.101477
- García AP, Palou M, Sánchez J, et al. Moderate caloric restriction during gestation in rats alters adipose tissue sympathetic innervation and later adiposity in offspring. PLoS ONE. 2011;6:e17313.10.1371/journal.pone.0017313
- Matveyenko AV, Singh I, Shin BC, et al. Differential effects of prenatal and postnatal nutritional environment on β-cell mass development and turnover in male and female rats. Endocrinology. 2010;151:5647–5656.10.1210/en.2010-0978
- Honma T, Kitano Y, Kijima R, et al. Comparison of the health benefits of different eras of japanese foods: lipid and carbohydrate metabolism focused research. Nippon Shokuhin Kagaku Kogaku Kaishi. 2013;60:541–553.10.3136/nskkk.60.541
- Htakeyama Y, Kitano Y, Honma T. Effects of Japanese food typical of that consumed during different periods in the past on breast milk-mediated lipid metabolism in offspring. J Jpn Soc Nutr Food Sci. 2014;67:255–270.
- Kitano Y, Honma T, Hatakeyama Y, et al. Effects of historical differences in components of the Japanese diet on the risk of obesity in mice. J Jpn Soc Nutr Food Sci. 2014;67:73–85.10.4327/jsnfs.67.73
- Ministry of Health, Labour and Welfare. Standard of Japanese Meal Intake. 2010 (DAI-ICHI SHUPPAN) pp 62–117.
- The Ministry of Health, Labour, and Welfare. 2010. National Health and Nutrition Survey Report. pp 62–63.
- Ministry of Education Culture Sports Science and Technology. Standard Tablesof Food Composition in Japan ( Seventh Revised Edition). 2015. pp 6–12.
- Tsuduki T, Kitano Y, Honma T, et al. High dietary fat intake during lactation promotes development of diet-induced obesity in male offspring of mice. J Nutr Sci Vitaminol (Tokyo). 2013;59:384–392.10.3177/jnsv.59.384
- Tsuduki T, Kikuchi I, Kimura T, et al. Intake of mulberry 1-deoxynojirimycin prevents diet-induced obesity through increases in adiponectin in mice. Food Chem. 2013;139:16–23.10.1016/j.foodchem.2013.02.025
- Rouser G, Fleischer S, Yamamoto A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids. 1970;5:494–496.10.1007/BF02531316
- Honma T, Shinohara N, Ito J, et al. High-fat diet intake accelerates aging, increases expression of Hsd11b1, and promotes lipid accumulation in liver of SAMP10 mouse. Biogerontology. 2012;13:93–103.10.1007/s10522-011-9363-2
- Hoekstra M, Kruijt JK, Van Eck M, et al. Specific gene expression of ATP-binding cassette transporters and nuclear hormone receptors in rat liver parenchymal, endothelial, and kupffer cells. J Biol Chem. 2003;278:25448–25453.10.1074/jbc.M301189200
- Ota M, Mori K, Nakashima A, et al. Resistance to excessive bodyweight gain in risperidone-injected rats. Clin Exp Pharmacol Physiol. 2005;32:279–287.10.1111/cep.2005.32.issue-4
- Scott DK, Collier JJ, Doan TT, et al. A modest glucokinase overexpression in the liver promotes fed expression levels of glycolytic and lipogenic enzyme genes in the fasted state without altering SREBP-1c expression. Mol Cell Biochem. 2003;254:327–337.10.1023/A:1027306122336
- Kim J, Park J, Hong S, Kim MK. Effect of corn gluten and its hydrolysate consumptions on weight reduction in rats fed a high-fat diet. Nutr Res Pract. 2009;3:200–207.10.4162/nrp.2009.3.3.200
- Suzuki M, Shibanuma M, Kimura S. Effect of severe maternal dietary restriction on growth and intra-abdominal adipose tissue weights in offspring rats. J Nutr Sci Vitaminol (Tokyo). 2010;56:293–298.10.3177/jnsv.56.293
- Lukaszewski MA, Mayeur S, Fajardy I, et al. Maternal prenatal undernutrition programs adipose tissue gene expression in adult male rat offspring under high-fat diet. Am J Physiol Endocrinol Metab. 2011;301:E548–E559.10.1152/ajpendo.00011.2011
- Palou M, Konieczna J, Torrens JM, et al. Impaired insulin and leptin sensitivity in the offspring of moderate caloric-restricted dams during gestation is early programmed. J Nutr Biochem. 2012;23:1627–1639.10.1016/j.jnutbio.2011.11.005
- Yi X, Cao S, Chang B, et al. Effects of acute exercise and chronic exercise on the liver leptin-AMPK-ACC signaling pathway in rats with type 2 diabetes. J Diabetes Res. 2013;2013:946432.
- Mazzucco MB, Fornes D, Capobianco E, et al. Maternal saturated-fat-rich diet promotes leptin resistance in fetal liver lipid catabolism and programs lipid homeostasis impairments in the liver of rat offspring. J Nutr Biochem. 2016;27:61–69.10.1016/j.jnutbio.2015.08.019
- Serviddio G, Bellanti F, Vendemiale G. Free radical biology for medicine: learning from nonalcoholic fatty liver disease. Free Radic Biol Med. 2013;65:952–968.10.1016/j.freeradbiomed.2013.08.174
- Wahlang B, Falkner KC, Gregory B, et al. Polychlorinated biphenyl 153 is a diet-dependent obesogen that worsens nonalcoholic fatty liver disease in male C57BL6/J mice. J Nutr Biochem. 2013;24:1587–1595.10.1016/j.jnutbio.2013.01.009
- Lu Y, Liu X, Jiao Y, et al. Periostin promotes liver steatosis and hypertriglyceridemia through downregulation of PPARα. J Clin Invest. 2014;124:3501–3513.10.1172/JCI74438
- Tanaka S, Yamamoto H, Nakahashi O, et al. Dietary phosphate restriction induces hepatic lipid accumulation through dysregulation of cholesterol metabolism in mice. Nutr Res. 2013;33:586–593.10.1016/j.nutres.2013.05.004
- Stylianou IM, Clinton M, Keightley PD, et al. Microarray gene expression analysis of the Fob3b obesity QTL identifies positional candidate gene Sqle and perturbed cholesterol and glycolysis pathways. Physiol Genomics. 2005;20:224–232.10.1152/physiolgenomics.00183.2004
- Dikkers A, de Boer JF, Groen AK, et al. Hepatic ABCG5/G8 overexpression substantially increases biliary cholesterol secretion but does not impact in vivo macrophage-to-feces RCT. Atherosclerosis. 2015;243:402–406.10.1016/j.atherosclerosis.2015.10.010
- Jianhua L, Xueqin M, Jifen H. Expression and clinical significance of LXRα andSREBP-1c in placentas of preeclampsia. Open Med (Wars). 2016;11:292–296.
- Beukers F, Cranendonk A, de Vries JI, et al. Catch-up growth in children born growth restricted to mothers with hypertensive disorders of pregnancy. Arch Dis Child. 2013;98:30–35.10.1136/archdischild-2012-302510
- Hack M, Schluchter M, Margevicius S, et al. Trajectory and correlates of growth of extremely-low-birth-weight adolescents. Pediatr Res. 2014;75:358–366.10.1038/pr.2013.209
- King V, Norman JE, Seckl JR, et al. Post-weaning diet determines metabolic risk in mice exposed to overnutrition in early life. Reprod Biol Endocrinol. 2014;12:73.10.1186/1477-7827-12-73