
ABSTRACT
Previous studies have reported that cod caught using a newly developed dual sequential codend showed significant reduction in external catch-related damage compared to cod caught by a conventional codend. In this study, it was investigated whether this new codend affects residual blood in cod fillets and/or mitigates the effect of postponed bleeding. Residual blood was assessed by measuring hemoglobin using VIS/NIR hyperspectral imaging. No significant differences in hemoglobin content between fish caught by the two different codends were proven, and this was true for whole fillets and fillet compartments (loin, belly, and tail). However, a significant effect of postponed bleeding on the hemoglobin concentration in cod was demonstrated, and this effect was similar whether the fish was caught using the conventional or the sequential codend. Fish bled 40 min after catch had significantly higher levels of hemoglobin compared to fish bled immediately after catch. When comparing the hemoglobin content in the different fillet sections, significantly higher levels of hemoglobin were found in the belly compared to the loin and tail.
Introduction
Atlantic cod (Gadus morhua) is the dominant species in the demersal fishery in the Barents Sea (Nedreaas et al. Citation2011; Yaragina et al. Citation2011). During the last 5 years, the average annual catch of Atlantic cod in the Northeast Arctic (ICES Subareas I and II) was 810,664 metric tonnes (ICES Citation2020). The two main nations targeting Atlantic cod are Norway and Russia (Shamray and Sunnanå Citation2011), and about 70% of the annual quota is caught using bottom trawls (ICES Citation2018; Yaragina et al. Citation2011).
Trawl-caught cod has often been associated with reduced quality (Digre et al. Citation2010; Olsen et al. Citation2013; Rotabakk et al. Citation2011), and the most common quality defects in trawl-caught white fish are gear marks, skin abrasion, pressure injuries, internal and external ecchymosis, and insufficient exsanguination (Digre et al. Citation2017; Olsen et al. Citation2013; Rotabakk et al. Citation2011). Such quality defects may lead to a downgrading of the fish and economic loss for the producer, and a negative correlation between the amounts of catch defects and proportions of high-value products has been found (Margeirsson et al. Citation2006; Sogn-Grundvåg et al. Citation2021).
Quality defects in trawl-caught fish can be related to the design of the gear itself and to trawling procedures. Several studies have investigated the effect of various trawling procedures and trawl configurations on catch quality (Brinkhof et al. Citation2018a, Citation2018b, Citation2021; Sistiaga et al. Citation2020; Tveit et al. Citation2019). The large mesh size (i.e., minimum 130 mm codend mesh size) that is required by law is believed to contribute to catch defects (Brinkhof et al. Citation2018b), and a new codend concept aimed at improving the quality of trawl-caught fish has been tested (Brinkhof et al. Citation2018b). The new concept consists of a dual sequential codend: the first codend segment has size-selective properties as required by law, and the sequential codend segment is designed to reduce the strain on the fish. The sequential codend segment, which opens at a predefined depth during haul-back, consists of knotless small-sized meshes (i.e., 6 mm mesh size) that retain water inside the codend during haul-back, and it should reduce gear marks, bruises, and stress due to crowding. Brinkhof et al. (Citation2018b) showed that compared to cod caught with a conventional codend, the fish caught using the sequential codend had significantly reduced external catch-related damage, as evaluated by the catch damage index (CDI; Esaiassen et al. Citation2013).
The amounts and types of external catch damage registered by the CDI directly impact the use of caught fish in various fish products. Residual blood present in fish fillets also influences fish quality (Botta et al. Citation1987; Digre et al. Citation2017; Esaiassen et al. Citation2004; Margeirsson et al. Citation2007; Olsen et al. Citation2013, Citation2014; Svalheim et al. Citation2019, Citation2020), but it is not included in the CDI. Therefore, in this study, it was investigated whether the dual sequential codend, compared to a conventional codend, could reduce the amount of residual blood in cod fillets as measured by the hemoglobin levels in the fillets.
Time from catch to bleeding is of utmost importance in achieving adequate exsanguination and minimizing residual blood (Borderías and Sánchez-Alonso Citation2011; Botta et al. Citation1986; Digre et al. Citation2011; Margeirsson et al. Citation2007; Olsen et al. Citation2014). According to Olsen et al. (Citation2013), it is not unusual for large hauls of fish caught by bottom trawls to be kept in storage bins for hours before bleeding and gutting. Poor exsanguination decreases fillet whiteness and thus its applicability to high-quality products that require a white fillet, such as fillet loins and clipfish. Hence, improving the quality of trawl-caught fish (e.g., minimizing residual blood by avoiding postponed bleeding) could increase its value and possibly expand the bottom trawl fishery market.
The aim of this study was to investigate the effect of codend type and postponed bleeding on the residual blood levels in cod fillets measured as hemoglobin concentration. In addition, hemoglobin content in different fillet sections (loin, belly, and tail) of trawl-caught cod was compared.
Materials and methods
Study area and trawl configuration
Atlantic cod was caught by the research trawler R/V Helmer Hanssen (63.8 m, 4080 HP) between 27 February and 5 March 2018. The fishing area was located along the coast of northern Norway in the southern Barents Sea (N 71°21′ E 23°43′–N 71°21′ E 24°24′). The towing speed (over ground) varied between 3.0 and 3.5 knots (average 3.3 knots). The cod were caught using two identical and commercially rigged trawls equipped with two different codend concepts: a conventional codend and a new dual sequential codend ().
Figure 1. Setup of the trawls with the conventional codend and the dual sequential codend.
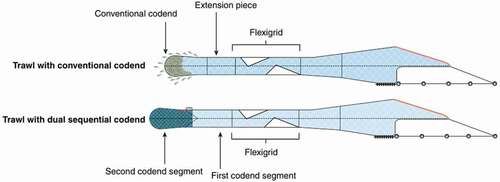
The trawls contained a set of Injector Scorpion otter boards for bottom trawling (3100 kg, 8 m2) with 3-m-long backstraps followed by a 7-m-long chain, which was linked to 60-m-long sweeps that were equipped with an Ø53 cm steel bobbin in the center to avoid excessive abrasion of the sweeps. The 46.9-m-long ground gear consisted of a 14-m-long chain (Ø 19 mm) with three equally spaced bobbins (Ø53 cm) on each side and an 18.9-m-long rockhopper gear in the center composed of Ø53 cm rubber discs. The ground gear was attached to the 19.2 m long fishing line of the trawl. The trawls used were a two-panel Alfredo 3 fish trawl built from polyethylene with a 155 mm nominal mesh size with a circumference of 420 meshes. The headline of the trawls was 35.6 m long and equipped with 170 floats (Ø20 cm). Both trawls were equipped with a flexigrid sorting system (55 mm bar spacing), which is one of the compulsory sorting grids for the trawl fishery in the Northeast Atlantic (Sistiaga et al. Citation2016).
In the conventionally configured trawl, the section with the flexigrid was followed by a 9-m-long extension piece (150 mm mesh size), which was preceded by an 11-m-long two-panel codend (). The codend was 11 m long and built from single-braided Ø8 mm Euroline Premium (Polar Gold) netting in the lower panel and double-braided Ø4 mm polyethylene in the upper panel, with a mean (± standard deviation, SD) mesh size of 133 ± 5.1 mm.
Figure 2. The conventional codend during a) towing and b) haul-back. The dual sequential codend consists of two codend segments: the first codend segment with size-selective properties as legislated, in which the fish are retained during c) trawling, and a second codend segment with expected quality-improvement properties, into which the fish falls during d) haul-back.
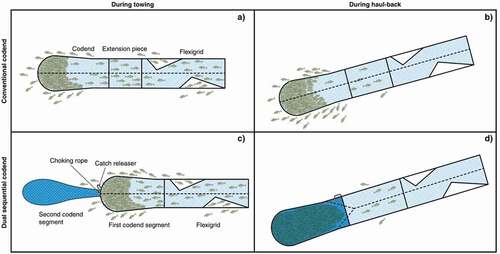
The second trawl was equipped with a dual sequential codend mounted directly to the flexigrid section (Brinkhof et al. Citation2018b), and it consisted of two codend segments: the first codend segment had the legislated selective properties, and the second (sequential) codend segment had quality-improving properties (). The first codend segment was built in the same way as the conventional codend and had a mean (±SD) mesh size of 139 ± 2.5 mm. The second codend segment was 10 m long and consisted of four panels. It had a nominal mesh size of 6 mm and a circumference of 1440 meshes (360 meshes in each panel). The codend segment was strengthened with an outer codend consisting of Ultracross knotless netting with a nominal mesh size of 112 mm (90 meshes in circumference) and four lastridge ropes, which were 5% shorter than the netting in the codend segment (). The two codend segments were connected to create a two-panel codend. The entrance of the second codend segment was closed during fishing because this codend does not have the size-selective properties required by law due to its small mesh size. During fishing at the seabed, the fish were retained in the first codend segment, which fulfilled the minimum mesh size requirement (). The entrance of the second codend segment was opened at a preset depth of 120 m during haul-back, thereby enabling free passage of fish from the selective codend segment into the codend with quality improving attributes (Brinkhof et al. Citation2018b) ().
Fish sampling
A total of 10 hauls were conducted, alternating between hauls with the conventional codend (5 hauls) and those with the dual sequential codend (5 hauls) (). Immediately after each haul, 28 cod were randomly sampled (weight: 3325 ± 779 g, length: 72 ± 6 cm, mean ± SD). The fish were split into four groups (n = 7), which were bled 0, 20, 40, and 60 min after being brought on board, respectively. Prior to bleeding by cutting the isthmus, the fish were stunned by a blow to the head, individually tagged, and kept in dry baskets. After exsanguination in running seawater for 30 min, individual length and weight before and after gutting were recorded. The fish were then beheaded and stored in dry baskets until freezing. The fish were frozen in blocks in commercial vertical plate freezers down to −18°C, packed in approved laminated paper bags and then stored at −30°C until landing. After the sea trail, the fish were transported to the laboratory in Nofima, Tromsø, and stored for 5 months at −20°C. Before analyses, the fish were thawed in tanks containing 600 L of chilled water (1°C) for 24 h and then on ice in 70 L fish crates with the belly cavity facing downward for another 24 h. Immediately after thawing, the fish were manually filleted with the skin retained. To enable evaluation of the belly flap, the black peritoneum was removed. The fillets were then cleaned in running freshwater. Both fillets (right and left side) from each fish were subjected to analyses. However, one fish from haul 6 was lost, resulting in 558 fillets for evaluation of residual blood.
Table 1. Overview of the hauls showing codend type, towing time, depth, and estimated catch size.
Hemoglobin measurement by hyperspectral imaging
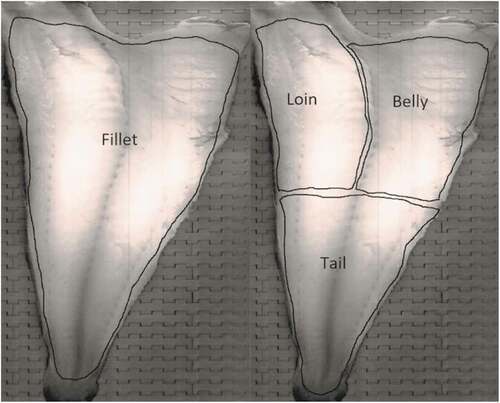
The hemoglobin content was estimated as a measure of the amount of blood in the muscle. Hyperspectral imaging of the fillets was conducted using an interactance setup (Sivertsen et al. Citation2012) in the wavelength range 400 to 1000 nm. This ensures that the light travels some distance inside the fillet before it is recorded. The muscle hemoglobin concentration in milligrams per gram of muscle (mg Hb/g muscle) was calculated following the procedure outlined in Skjelvareid et al. (Citation2017). To estimate the amount of blood in each of the fillet sections (loin, belly, and tail), the respective sections were marked manually in each image of the fillets (), and the amount of hemoglobin was calculated for each section separately. Two fillets were retained per fish (left and right), and the hemoglobin concentrations are reported as the average value of the left and right side.
Figure 3. Illustration of the fillet sections used to estimate hemoglobin content by hyperspectral analysis.
Data analysis
Statistical analysis was performed using IBM SPSS Statistics for Windows, version 28.0.0.0. The distribution was tested for normality using the Kolmogorov–Smirnov test and density plots and further tested for homogeneity of variance using Levene’s test. A two-way analysis of covariance was conducted to examine the effects of codend and bleeding time on hemoglobin concentration after controlling for catch size, catch depth, and towing time. Catch size and towing time were included as covariates because previous studies have reported reduction in fish quality with increasing catch size and towing time (Digre et al. Citation2017; Margeirsson et al. Citation2007; Olsen et al. Citation2013; Rotabakk et al. Citation2011; Veldhuizen et al. Citation2018). Catch depth was included because capture depth may affect the condition of the fish (Digre et al. Citation2017) and catch quality (Brinkhof et al. Citation2018a). A Bonferroni post hoc test and pairwise comparison were used to determine significant differences between the groups. A one-way analysis of variance with the Game–Howell post hoc test (the groups had heterogeneity of variance) was conducted to determine whether there were any statistically significant differences in mean hemoglobin concentration between loin, belly, and tail. p-values <0.05 were considered to be statistically significant.
Results and discussion
No statistically significant two-way interaction between codend and bleeding time on hemoglobin concentration in cod fillets was detected when controlling for catch size, catch depth, and towing time (F(3, 268) = 0.754, p = .521, partial η2 = 0.008). Because there was no statistically significant interaction between codend and bleeding time on hemoglobin levels, the data were combined, and the analysis was followed up using the main effects of codend and bleeding time.
Effect of codend on hemoglobin concentration in cod fillets
shows the concentration of hemoglobin in fillets and fillet compartments from cod caught using the conventional and sequential codends. The adjusted marginal mean hemoglobin concentration was similar in fillets from fish caught using both the conventional and the sequential codend, 0.144 g Hb/g muscle. Thus, no significant effect of codend was demonstrated (F(1, 268) = 0.026, p = 0,871, partial η2 = 0.00).
Table 2. Adjusted means with standard error for hemoglobin concentration (mg Hb/g muscle) for fillets and for the different fillet sections (loin, belly, and tail) in fish caught by the conventional and sequential codend.
This result does not align with the findings by Brinkhof et al. (Citation2018b), who reported that the sequential codend improved quality of trawl-caught cod. However, in the study by Brinkhof et al. (Citation2018b), external damages were assessed, whereas residual blood in the muscle was assessed in the present study. Svalheim et al. (Citation2020) showed that exhaustive swimming and crowding cause fillet redness and increased muscle hemoglobin. Thus, one possible explanation for the contradictory results could be that the dual sequential codend reduces external detectable damage caused during haul-back but not residual blood in muscle caused by exhaustive swimming and crowding during trawling.
On the other hand, it is possible that an effect of the sequential codend on the amount of residual blood in the fish in the present study is concealed by the fact that the catches from the hauls using the sequential codend were generally larger than the catches from the conventional codend and that the influence of codend type could not be separated from the influence of catch size. Previous studies have reported a reduction in fish quality with increasing catch size and towing time (Digre et al. Citation2017; Margeirsson et al. Citation2007; Olsen et al. Citation2013; Rotabakk et al. Citation2011; Veldhuizen et al. Citation2018). Compared to commercial fisheries, where catches easily exceed 10 tonnes, the catch size in this study was very low, and the towing time for each haul in this study was relatively short (45–90 min). It is thus also possible that the small catch size and short towing time may have concealed effects of codend type on the amount of residual blood in the fish. Hence, drawing a definite conclusion about the effects of the different codends on residual blood in cod fillets is not possible from this experiment.
Effect of bleeding time on hemoglobin concentration in cod fillets
The results show that the time before bleeding is important for the quality of trawl-caught cod regardless of the codend used, and a statistically significant main effect of bleeding time on adjusted marginal mean hemoglobin concentration in cod fillets was demonstrated (F(3, 268) = 13.682, p < .001, partial η2 = 0.133). As the time from catch until bleeding increased, the amount of residual blood in the fillets and fillet compartments increased ().
Table 3. Adjusted means with standard error for hemoglobin concentration (mg Hb/g muscle) for fillets and for the different fillet sections (loin, belly, and tail) for fish bled 0, 20, 40, and 60 min after catch.
Marginal mean hemoglobin concentration was significantly higher in fillets from fish bled 40 min after catch (0.147 mg Hb/g muscle) than in fillets from fish bled immediately (0 min) after catch (0.137 mg Hb/g muscle) (p < .001). These findings are in accordance with previous studies as reviewed by Erikson et al. (Citation2021), which showed that postponed bleeding negatively affected the quality of the fish. The effects of other factors on the blood drainage of fish, such as slaughter and cutting method (Digre et al. Citation2011; Olsen et al. Citation2006; Roth et al. Citation2005), temperature (Olsen et al. Citation2006), and water flow (Eliasson et al. Citation2020), have also been investigated. However, it has been shown that the time before bleeding is crucial. Botta et al. (Citation1986) showed that time was more important than the cutting procedure in trawl-caught cod that were bled 0, 1, 2, and 3 h after catch. Olsen et al. (Citation2014) studied cod bled after 0, 30, 60, and 180 min and recommended that cod should be bled within 30 min after catch. In the latter study, the fish were kept alive after catch; they were carefully netted and stunned before bleeding. According to Svalheim et al. (Citation2019), this recommendation based on results from unstressed fish is likely to be less relevant in commercial fisheries, where fish are exposed to a number of stressors. Svalheim et al. (Citation2019, Citation2020) showed that exhaustive swimming, crowding, and especially air exposure, are associated with increased metabolic stress, resulting in increased amount of residual blood in muscles after bleeding. In the present experiment, due to fish welfare considerations, the fish were stunned with a blow to the head prior to air exposure and postponed bleeding. This has probably influenced the results, and the effect of postponed bleeding may have been higher if the fish were kept conscious in air prior to bleeding.
Hemoglobin concentration in fillet sections
As with whole fillets, no statistically significant two-way interaction between codend and bleeding time was demonstrated for loin, belly, or tail on hemoglobin concentration when controlling for catch size, catch depth, and towing time. No significant effect of codend type on hemoglobin concentration was uncovered either. However, a statistically significant main effect of bleeding time on the adjusted marginal mean hemoglobin concentration was demonstrated for loin, belly, and tail parts of the fillets (, ).
Figure 4. Mean hemoglobin concentration with 95% confidence intervals in the different fillet sections (loin, tail, and belly). Different letters indicate significant differences between the fillet section for each bleeding time (p < .001).
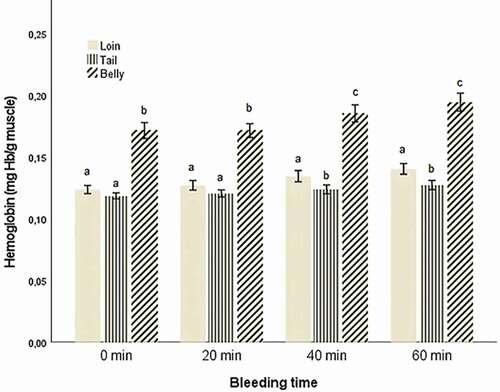
Comparing the hemoglobin content in the different fillet sections revealed that the belly contained a significantly higher level of hemoglobin than the loin and tail (, ). Olsen et al. (Citation2008) studied the distribution of heme-pigments among fillet sections in bled, unbled, stressed, and unstressed Atlantic cod and found that the largest differences in heme-pigments in white muscle from bled and unbled fish were found in the belly flaps, and there was generally more blood in the belly compartment compared to other fillet sections. However, their analyses focused on the effects of stress prior to slaughter and whether the fish was bled or not rather than on differences in blood content between sections within the same fillet.
To the best of our knowledge, reports on how postponed bleeding affects different fillet sections are scarce. There are, however, a few Norwegian reports addressing the topic (Akse et al. Citation2012; Tobiassen et al. Citation2019), and the results in the present study are in accordance with these reports. Akse et al. (Citation2012) reported that residual blood in both the belly and loin of Atlantic cod was significantly affected by postponed bleeding and that there was more residual blood in the belly of the cod bled after 30 min than in the loin of the cod bled after 180 min. Additionally, using hyperspectral imaging, Tobiassen et al. (Citation2019) showed that the belly of haddock (Melanogrammus aeglefinus) was more vulnerable to postponed bleeding compared with the other fillet parts.
Conclusion
No significant effect of the sequential codend on the amount of residual blood in cod fillets compared to cod caught using the conventional codend was proven. There was a statistically significant effect of postponed bleeding on the amount of residual blood in cod fillets for both the dual sequential codend and the conventional codend. Postponed bleeding led to a higher amount of residual blood in the fish regardless of the codend used. Furthermore, it is shown that the belly of cod contained significantly higher amounts of residual blood than the loin and tail.
Acknowledgements
We thank the crew of R/V Helmer Hanssen, technicians Ivan Tatone and Kunuk Lennert, and Dr Jesse Brinkhof for the help provided during the sea trial. We also thank researchers at Nofima, Dr Tatiana N. Ageeva and Dr Stein Harris Olsen, for valuable assistance during the analyses.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Akse L, Joensen S, Heia K, Tobiassen T, Sivertsen AH, Wang PA. 2012. Bleeding of cod – bleeding methods and time before bleeding or direct gutting. Tromsø (Norway): Nofima. Report No: 19/2012. ISBN 978-82-7251-988-8 (in Norwegian).
- Borderías AJ, Sánchez-Alonso I. 2011. First processing steps and quality of wild and farmed fish. J Food Sci. 76(1):R1–R5.
- Botta JR, Squires BE, Johnson J. 1986. Effect of bleeding/gutting procedures on the sensory quality of fresh raw Atlantic cod (Gadus morhua). Can Inst Food Sci Technol J. 19(4):186–90.
- Botta JR, Bonnell G, Squires BE. 1987. Effect of method of catching and time of season on sensory quality of fresh raw Atlantic cod (Gadus morhua). J Food Sci. 52(4):928–31.
- Brinkhof J, Larsen RB, Herrmann B, Olsen SH. 2018a. Assessing the impact of buffer towing on the quality of Northeast Atlantic cod (Gadus morhua) caught with a bottom trawl. Fish Res. 206:209–19.
- Brinkhof J, Olsen SH, Ingólfsson ÓA, Herrmann B, Larsen RB. 2018b. Sequential codend improves quality of trawl-caught cod. PLoS One. 13(10):e0204328.
- Brinkhof J, Herrmann B, Sistiaga M, Larsen RB, Jacques N, Gjøsund SH. 2021. Effect of gear design on catch damage on cod (Gadus morhua) in the Barents sea demersal trawl fishery. Food Control. 120:e107562.
- Digre H, Hansen UJ, Erikson U. 2010. Effect of trawling with traditional and ‘T90’ trawl codends on fish size and on different quality parameters of cod Gadus morhua and haddock Melanogrammus aeglefinus. Fish Sci. 76(4):549–59.
- Digre H, Erikson U, Misimi E, Standal IB, Gallart-Jornet L, Riebroy S, Rustad T. 2011. Bleeding of farmed Atlantic cod: Residual blood, color, and quality attributes of pre-and postrigor fillets as affected by perimortem stress and different bleeding methods. J Aquat Food Prod Technol. 20(4):391–411.
- Digre H, Rosten C, Erikson U, Mathiassen JR, Aursand IG. 2017. The on-board live storage of Atlantic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) caught by trawl: Fish behaviour, stress and fillet quality. Fish Res. 189:42–54.
- Eliasson S, Arason S, Margeirsson B, Palsson OP. 2020. Onboard evaluation of variable water flow and recirculation effects on bleeding of Atlantic cod (Gadus morhua). Foods. 9(11):1519.
- Erikson U, Uglem S, Greiff K. 2021. Freeze-Chilling of whitefish: effects of capture, on-board processing, freezing, frozen storage, thawing, and subsequent chilled storage—a review. Foods. 10(11):2661.
- Esaiassen M, Nilsen H, Joensen S, Skjerdal T, Carlehög M, Eilertsen G, Elvevoll EO. 2004. Effects of catching methods on quality changes during storage of cod (Gadus morhua). LWT - Food Sci Technol. 37(6):643–48.
- Esaiassen M, Akse L, Joensen S. 2013. Development of a catch-damage-index to assess the quality of cod at landing. Food Control. 29(1):231–35.
- ICES. 2018. ICES advice on fishing opportunities, catch, and effort Arctic Ocean, Barents Sea, Faroes, Greenland Sea, Icelandic Waters, and Norwegian Sea ecoregions – cod (Gadus morhua) in subareas 1 and 2 (Northeast Arctic). ICES [accessed 2020 Aug 20]. doi:10.17895/ices.pub.4412.
- ICES. 2020. Arctic fisheries working group (AFWG). ICES Sci Rep. 2(52):577. [accessed 2020 Aug 20]. doi:10.17895/ices.pub.6050.
- Margeirsson S, Nielsen AA, Jonsson GR, Arason S. 2006. Effect of catch location, season and quality defects on value of Icelandic cod (Gadus morhua) products. In: Luten JB, Jacobsen C, Bekaert K, Sæbø A, Oehlenschläger J, editors. Seafood research from fish to dish – quality, safety & processing of wild and farmed fish. The Netherlands: Wageningen Academic Publishers. p. 265–74.
- Margeirsson S, Jonsson GR, Arason S, Thorkelsson G. 2007. Influencing factors on yield, gaping, bruises and nematodes in cod (Gadus morhua) fillets. J Food Eng. 80(2):503–08.
- Nedreaas KH, Drevetnyak KV, Shamray EA. 2011. Commercial data. In: Jakobsen T, Ozhigin VK, editors. The Barents Sea: Ecosystem, resources, management: Half a century of Russian-Norwegian cooperation. Trondheim (Norway): Tapir Academic Press. p. 609–20.
- Olsen SH, Joensen S, Tobiassen T, Heia K, Akse L, Nilsen H. 2014. Quality consequences of bleeding fish after capture. Fish Res. 153:103–07.
- Olsen SH, Sørensen NK, Larsen R, Elvevoll EO, Nilsen H. 2008. Impact of pre-slaughter stress on residual blood in fillet portions of farmed Atlantic cod (Gadus morhua)—measured chemically and by Visible and Near-infrared spectroscopy. Aquaculture. 284(1–4):90–97.
- Olsen SH, Sorensen NK, Stormo SK, Elvevoll EO. 2006. Effect of slaughter methods on blood spotting and residual blood in fillets of Atlantic salmon (Salmo salar). Aquaculture. 258(1–4):462–69.
- Olsen SH, Tobiassen T, Akse L, Evensen TH, Midling KØ. 2013. Capture induced stress and live storage of Atlantic cod (Gadus morhua) caught by trawl: Consequences for the flesh quality. Fish Res. 147:446–53.
- Rotabakk BT, Skipnes D, Akse L, Birkeland S. 2011. Quality assessment of Atlantic cod (Gadus morhua) caught by longlining and trawling at the same time and location. Fish Res. 112(1–2):44–51.
- Roth B, Torrisen OJ, Slinde E. 2005. The effect of slaughtering procedures on blood spotting in rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Aquaculture. 250(3–4):796–803.
- Shamray EA, Sunnanå K. 2011. Development of management strategies. In: Jakobsen T, Ozhigin VK, editors. The Barents Sea: Ecosystem, resources, management: Half a century of Russian-Norwegian cooperation. Trondheim (Norway): Tapir Academic Press. p. 532–40.
- Sistiaga M, Brinkhof J, Herrmann B, Grimaldo E, Langård L, Lilleng D. 2016. Size selective performance of two flexible sorting grid designs in the Northeast Arctic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) fishery. Fish Res. 183:340–51.
- Sistiaga M, Herrmann B, Brinkhof J, Larsen RB, Jacques N, Santos J, Gjøsund SH. 2020. Quantification of gear inflicted damages on trawl-caught haddock in the Northeast Atlantic fishery. Mar Pollut Bull. 157:e111366.
- Sivertsen AH, Heia K, Hindberg K, Godtliebsen F. 2012. Automatic nematode detection in cod fillets (Gadus morhua L.) by hyperspectral imaging. J Food Eng. 111(4):675–81.
- Skjelvareid MH, Heia K, Olsen SH, Stormo SK. 2017. Detection of blood in fish muscle by constrained spectral unmixing of hyperspectral images. J Food Eng. 212:252–61.
- Sogn-Grundvåg G, Zhang D, Henriksen E, Bendiksen BI, Hermansen Ø. 2021. Fish quality and market performance: The case of the coastal fishery for Atlantic cod in Norway. Mar Policy. 127:e104449.
- Svalheim RA, Burgerhout E, Heia K, Joensen S, Olsen SH, Nilsen H, Tobiassen T. 2019. Differential response to air exposure in crowded and uncrowded Atlantic cod (Gadus morhua): Consequences for fillet quality. Food Biosci. 28:15–19.
- Svalheim RA, Aas-Hansen Ø, Heia K, Karlsson-Drangsholt A, Olsen SH, Johnsen HK. 2020. Simulated trawling: Exhaustive swimming followed by extreme crowding as contributing reasons to variable fillet quality in trawl-caught Atlantic cod (Gadus morhua). PLoS One. 15(6):e0234059.
- Tobiassen T, Martinsen G, Kristoffersen S, Hustad A, Olsen SH, Heia K, Joensen S, Ingolfsson O, Nordtvedt TS. 2019. Controlled slaughter of live stored haddock provides great advantages during processing and cold storage. Tromsø (Norway): Nofima. Report No: 10/2019. ISBN 978-82-8296-587-3 (in Norwegian).
- Tveit GM, Sistiaga M, Herrmann B, Brinkhof J. 2019. External damage to trawl-caught northeast arctic cod (Gadus morhua): Effect of codend design. Fish Res. 214:136–47.
- Veldhuizen LJL, Berentsen PBM, De Boer IJM, Van De Vis JW, Bokkers EAM. 2018. Fish welfare in capture fisheries: A review of injuries and mortality. Fish Res. 204:41–48.
- Yaragina NA, Aglen A, Sokolov KM. 2011. Cod. In: Jakobsen T, Ozhigin VK, editors. The Barents Sea: Ecosystem, resources, management: Half a century of Russian-Norwegian cooperation. Trondheim (Norway): Tapir Academic Press. p. 225–70.