Abstract
Bacillus thuringiensis Berliner toxins are highly specific in terms of insect pest targeted. For example, Cry3Bb is active against Coleoptera. As a consequence, in the framework of using genetically modified plants expressing different Bt toxins in sustainable agriculture, there is a growing interest about the impact of these crops on the environment and on non-target organisms. The effects of genetically modified Bt-tomato plants expressing the toxin Cry3Bb on the aphid Macrosiphum euphorbiae (Thomas) and its natural enemies, the generalist predator Macrolophus caliginosus Wagner and the endoparasitoid wasp Aphidius ervi Haliday, were assessed under laboratory conditions. No significant differences between performance of M. euphorbiae were observed on genetically modified tomato plants (line UC82Bt) with respect to their near-isogenic control line (line UC82). Immunoassays did not detect Cry3Bb protein in M. euphorbiae developing on Bt-expressing tomato plants. Similarly, no significant differences were found on the longevity and prey consumption of M. caliginosus when fed aphids reared on UC82Bt or on UC82. Moreover, the genetic modification did not affect the attractiveness of uninfested tomato plants toward A. ervi. It is therefore concluded that Cry3Bb-expressing tomato plants did not show any acute adverse effects on the biological parameters of the non-target herbivore M. euphorbiae or its natural enemies, M. caliginosus and A. ervi, under laboratory conditions.
Introduction
Genetically modified plants expressing specific toxins from the entomopathogenic bacterium Bacillus thuringiensis Berliner have long since been entered in the agricultural practice in several countries. However, only in recent years, there is a growing interest about the impact of these crops on the entire ecosystem. Such interest is also prompted by the debate open about their sustainability in comparison to traditionally managed crops, where broad-spectrum insecticides are periodically applied. For these reasons, the potential effects on non-target organisms (NTO) need to be assessed as part of the environmental risk assessment (ERA) that takes place prior to the commercialization of any genetically modified (GM) crop (Nap et al. Citation2003; EFSA Citation2006). Priorities and criteria for selecting the most appropriate non-target species to be included in the ERA of GM plants may be quite different (e.g. Birch et al. Citation2004; Romeis et al., Citation2008). The list of potential non-target species and ecological functions that could be evaluated is vast, but the range of species and functions evaluated to date has been more restricted (Lövei and Arpaia Citation2005). Agriculture depends on several ecological functions that are essential to soil fertility and agricultural productivity (e.g. biological control of primary and secondary pests, crop pollination by animals, microbial decomposition and nutrient cycling, etc.). To ensure the sustainability of agricultural practices, the ecological services in agro-ecosystems need to be reinforced or, at least, preserved.
Herbaceous agro-ecosystems are typically simplified ecosystems compared with natural habitats due to the major impact of human activities and to the short time-span plants remain in the field. Even so, complex multi-trophic relationships are established between primary producers and consumers, commonly up to the fourth trophic level (Schmidt Citation2006). Any perturbations to the system, by changes in agricultural practices or environmental conditions, could affect the dynamics of trophic interactions in unpredictable ways. Most of the knowledge acquired so far in the area of GM crops and NTOs was obtained by studying commodity crops, which represent almost 100% of the current commercialized GM plants worldwide (James Citation2009). More insights on the environmental safety of new GM crop plants that could potentially enter agricultural markets would be therefore important, before such crops become available for commercial use.
In the present study we assessed the possible hazards for non-target arthropods involved in a trophic chain linked to a GM tomato (Solanum lycopersicum L.), a high value horticultural crop.
As a model, a non-commercial insect resistant GM tomato line expressing the B. thuringiensis Cry3Bb protein was used. This trait has been used to induce insect resistance in several crop plants, and commercial applications are currently available for GM maize plants resistant to the Western corn rootworm (James Citation2009); therefore more information about its interactions with non-target organisms are important.
Tomato plants are normally attacked by a large group of insect pests, characterized by different feeding habits (leaf chewers, sap and cell feeders, fruit borers, etc.) (Lange and Bronson Citation1981). Cry3Bb expressed in tomato targets coleopteran pests, such as the well-known Leptinotarsa decemlineata (Say), a pest of this crop, both in the open field and greenhouse. In this paper, we report the results of laboratory studies on the effects of GM tomato on survivorship, reproduction and development of the aphid Macrosiphum euphorbiae (Thomas) (Hemiptera: Aphididae). It is of interest to examine the impact of such toxins on aphids because of their pest status (Walgenbrach Citation1997), but also because of recent claims of Bt endotoxins activity against the pea aphid, Acyrthosiphon pisum (Harris) (Hemiptera: Aphididae) (Porcar et al. Citation2009).
We also investigated possible effects along food chains including M. euphorbiae, and tritrophic experiments were set up including two of its most efficient and widespread natural enemies, the generalist predator Macrolophus caliginosus Wagner (Hemiptera: Miridae) and the endoparasitoid Aphidius ervi (Haliday) (Hymenoptera: Braconidae), which are commercially available for biological control (Kennedy Citation2003). The parasitoid A. ervi has a role of specific and effective antagonist of M. euphorbiae in tomato crops and it was therefore chosen as a model organism to conduct behavioral observation in confined conditions.
The measurement endpoints considered at the third trophic level were longevity and prey consumption for the mirid predator M. caliginosus. Furthermore, the possible alteration of the attractiveness of tomato plants following the genetic transformation was assessed by a behavioral bioassay with the aphid parasitoid A. ervi.
Materials and methods
Plants
GM tomato plants were obtained from Metapontum Agrobios s.r.l. (Metaponto, MT, Italy). Tomato plants (line UC82) were transformed via Rhizobium radiobacter (Beijerinck and van Delden 1902) Young et al. (2001)=Agrobacterium tumefaciens (Smith and Townsend 1907) Conn 1942] according to Iannacone et al. (Citation2008) using a modified version of a Cry3Bb gene (Iannacone et al. Citation1997) derived from B. thuringiensis. Plants were successively selfed to achieve the homozygosis of the Cry3Bb gene. The expression of the gene in the plants selected for all laboratory bioassays was assessed using a Quickstix kit (RAISIO Diagnostics, Ardea, RM, Italy). All the tomato plants used in the different bioassays resulted positive as for the presence of the gene Cry3Bb.
Insects
Macrosiphum euphorbiae is permanently reared at Institute for Plant Protection, and the culture was started from field-collected colonies from tomato plants in Scafati (SA) in 1998. The colony is maintained in a growth chamber at 20±1°C, 65±5% relative humidity, 18 hours light:6 hours dark photoperiod on tomato cv UC82.
The initial strain of M. caliginosus was obtained from Koppert Italia s.r.l. (Bussolengo, VR, Italy). Mirids were reared in a climatic chamber at 25±1°C, 65±5% relative humidity, 18 hours light:6 hours dark photoperiod. They were fed daily with Myzus persicae Sulzer (Hemiptera: Aphididae) from a colony reproducing on tomato cv UC82 and with eggs of Ephestia kühniella Zeller (Lepidoptera: Pyralidae). Several discs of wet filter paper (Whatman n.1) were added in the rearing cages to enhance the longevity of the mirids. Adults were used in bioassays within 48 hours from emergence.
Aphidius ervi is permanently reared at Department of Agricultural Entomology and Zoology ‘Filippo Silvestri’ and has been since 1988 on its natural host, the pea aphid A. pisum maintained on potted broad bean plants (Vicia faba, cv Aquadulce) as described in Guerrieri et al. (Citation2002). The initial strain of the parasitoid was obtained from field collections in Campania (Italy) from A. pisum on alfalfa. The parasitoid culture is kept in a growth chamber at 20±1°C, 60±5% relative humidity and 18 hours light:6 hours dark photoperiod. The parasitoid colony is periodically refreshed by adding adult parasitoids from mummies collected on field-grown alfalfa plants. Parasitoids for the bioassays were reared as synchronized cohorts as previously reported (Guerrieri et al. Citation2002).
Biological parameters of M. euphorbiae
The effect of tomato UC82 expressing Cry3Bb toxin on the performance of M. euphorbiae in comparison to the near-isogenic cultivar/line UC82 was assessed by calculating aphid developmental parameters. In a glasshouse, we set up an isolator, made of a wood frame and fine mesh net (280 cm×80 cm×150 cm), that contained all the experimental plants. This cage was divided into 2 parts by a net, to prevent aphids shifting from a group of plants to the other. For each treatment (UC82-Bt and non-transformed UC82), 54 tomato plants, each constituting a replicate, were reared for 4 weeks after emergence. Each plant was infested with a single newly-born aphid, transferred with a soft brush. All plants were checked daily to assess presence of aphids, presence of exuviae (evidence of molting), presence and number of newly-laid nymphs and death. When reproduction started, the offspring were removed daily.
The data obtained were used to calculate the intrinsic rates of increase (r m ), which was computed by an iterative solution of the approximation of the Euler equation (Birch Citation1948)
where
with l x and m x representing the age-specific adult survival and the reproduction rates of female offspring at the interval age x (d), assigned by taking the mean development time in days + 0.5 as the starting point.
The accurate value of r m was then calculated by solving the equation
The tests were run in a glasshouse at 21±2°C, 75±10% relative humidity and 18 hours light:6 hours dark photoperiod.
Tritrophic bioassays with M. caliginosus
To assess the effect of Bt-tomato plants on the quality of the aphid as expressed by longevity and prey consumption of M. caliginosus, small arenas were set up in Petri dishes (diameter 55 mm). Apical leaves freshly cut from control and transformed plants were inserted into 3.5 ml of solidified agar confined to one side of the arena. On each leaf, five larvae of M. euphorbiae (first and second instar) were added using a soft brush along with an adult M. caliginosus. Prey consumption and predator mortality were checked daily.
Consumed prey were replaced daily by an equal number of aphids to those consumed to keep a constant 1:5 ratio between predator and prey in each arena. The experiments were stopped when 70% of the predators were dead. Two replicate experiments, each including 13 replicates, were run in a growth chamber at 25±1°C, 65±5% relative humidity and 16 hours light:8 hours dark photoperiod.
Flight behavior bioassay
The effect of GM tomatoes on the flight behavior of adult females of the parasitoid A. ervi was tested in single choice wind tunnel test. Experimental conditions during the bioassay were set as follows: temperature 20±1°C, relative humidity: 65±5%; wind speed: 25±5 cm/sec; distance between releasing vial and target: 50 cm; light intensity at releasing point: 3600 lux.
All experiments were conducted between the 3rd and the 7th hour from the onset of photophase and carried out over several days, with the targets presented in a random order to reduce the effect of temporal variability on the results. Ninety parasitoid females, 24–48 hours old, mated, fed with honey solution (10%) and naive, that is without any previous experience with either tomato and/or M. euphorbiae, were tested for each target. The percentage of response (oriented flights, landings on the target) was recorded and analyzed using an event-recording software (the Observer, Noldus Information Technology, Wageningen, the Netherlands).
For each treatment, 6 uninfested plants between 4 and 5 weeks after sowing were tested daily over two consecutive weeks. Bt and control plants were offered in a random order to avoid any influence of the time of test and parasitoid response.
Cry3Bb extraction and detection in M. euphorbiae
Assays to determine the presence of the Cry3Bb toxin in aphid whole bodies were performed using kits designed for the detection of the Bt toxin in plants (Burgio et al. Citation2007). Aphids were reared on UC82-Bt plants for several generations, and M. euphorbiae samples were regularly collected, weighted and frozen. Before extraction, the samples were pooled, to reach weights that are considered above the detectability level of the assays (Burgio et al. Citation2007 and references therein). The pooled samples were homogenized with a pestle with 100 µl of extraction buffer (as indicated in the kit's instructions) and centrifuged, and the surnatant was assayed either with Envirologix QuickStix (Catalogue No. AS 015 LS) or with Agdia Bt-Cry3Bb1 ELISA kit (Catalogue No. PSP 06100).
Aphids reared on non-Bt UC82 tomatoes were used as a control. The pooled samples consisted of 111 mg of aphids from Bt plants and 99 mg of aphids from control plants in the case of EnviroLogix QuickStix strip, and 87 and 91 mg of aphids from Bt plants and 67 and 67 mg of aphids from control plants for the Agdia ELISA kit.
Statistical analyses
The effect of Bt tomato on the aphid's developmental periods, demographic parameters, longevities, and mean number of offspring per aphid were analyzed using Student's t-test. Survivor curves were analyzed using Student's t-test (two samples, paired). Data from tritrophic bioassays were analyzed with the analysis of variance. For M. caliginosus longevity, data were analyzed considering a fixed maximum duration (right censored data). Differences between means were assessed using Student's t-test. The percentages of oriented flights and landings were compared by a G-test for independence with William's correction (Sokal and Rohlf Citation1981). The resulting values of G were compared with the critical values of χ 2 (Rohlf and Sokal Citation1995). All statistical analyses were performed using the SAS package (SAS Citation1989).
Results
Biological parameters of M. euphorbiae
Survival curves of the aphids on conventional and Bt tomato () were significantly different (t=5.86; p<0.001; df 54). In detail, by examining the daily percentage of survivors, it appears that after reaching the adult stage, aphids performed better on control plants but, at the end of the test, the reverse occurred, with a few individuals living for one more week on Bt plants (). Regardless of these differences, the values of longevity resulted similar for Bt and control plants (). No significant differences were found in the other biological parameters recorded for M. euphorbiae on control and Bt plants, leading to similar values of r m ().
Figure 1. Survival curves for Macrosiphum euphorbiae on conventional (UC82, dots) and Bt tomato plants (UC82Bt, triangles).
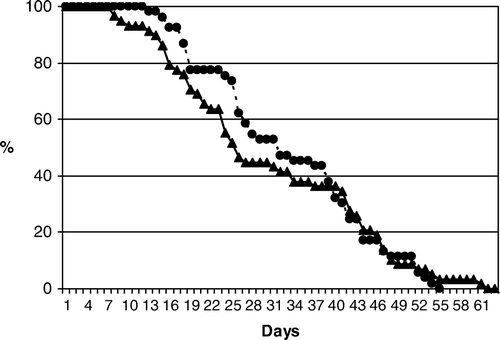
Table 1. Life history traits of the aphid Macrosiphum euphorbiae (Thomas) reared on conventional (UC82, n=54) and Bt tomato (UC82Bt, n=54).
None of the two immuno-assays used (ELISA and Quickstix) detected Cry toxin in the aphid samples from UC82-Bt plants.
Tritrophic bioassays with M. caliginosus
No statistical differences were found between the longevity of M. caliginosus adults on control and Bt tomato plants. In the first experiment, adults of the predator lived 6.0 days (±2.1) on control plants and 5.67 days (±1.87) on GM plants (p=0.41). In the second replicate we recorded a longevity of 5.01 days (±3.96) and 6.29 days (±3.79) on control and Bt plants, respectively (p=0.50).
Similarly, there was no significant difference in predator daily consumption between treatments.
Mean values were 0.76±0.41 aphid consumed on control plants and 1.11±0.27 aphids consumed on UC82Bt tomato (p=0.074). In the second experiment we recorded higher values of prey consumption on both control and Bt plants but, again, not significantly different between each other (control: 1.70±0.94 aphids; Bt: 2.08±1.39 aphids. p=0.67).
Flight behavior bioassay
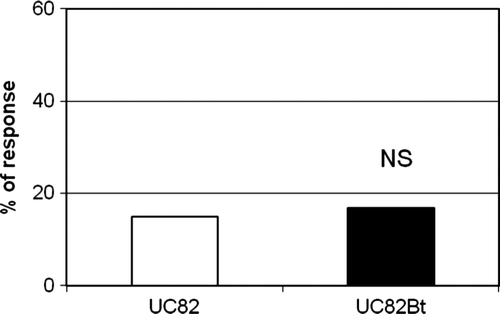
There was no significant difference between oriented flights and landings on target treatments by the parasitoid A. ervi toward the plant when control or Bt tomato plants were assayed (). Values recorded in our wind tunnel bioassays are similar to literature reports for uninfested tomato plants (Guerrieri et al. Citation2004; Corrado et al. Citation2007; Sasso et al. Citation2007).
Figure 2. Percentage of landings on the target by A. ervi toward conventional (UC82) and Bt (UC82 Bt) tomato plants not infested by aphids.
Discussion
There is a growing interest about the possible impact of genetically modified plants on non-target organisms in the environment (e.g. Craig et al. Citation2008; EFSA, Citation2008). Both direct and indirect effects on non-target herbivore species are deemed important because phytophagous insects can represent a way to transfer the toxin to higher trophic levels (e.g. Obrist et al. Citation2006).
In this paper we addressed the effects of exposing the aphid M. euphorbiae, a worldwide pest of several Solanaceae crops, for example potato, tomato, and eggplant (Walgenbrach Citation1997) to a Cry3Bb toxin expressed in a GM tomato line. The subsequent effects on two of its natural enemies, a parasitoid and a predator, were also studied.
The possible effects of Bt toxins on organisms at higher trophic levels delivered via GM plants and aphids have been described in several plant–Cry toxin systems, but the results are sometimes contrasting (Head et al. Citation2001; Raps et al. Citation2001; Dutton et al. Citation2002; Lumbierres et al. Citation2004; Liu et al. Citation2005; Burgio et al. Citation2007; Ramirez-Romero et al. Citation2008; Zhang et al. Citation2008).
However, the actual exposure of aphids to Cry toxins expressed in GM plants is still a contentious issue, since there are contrasting evidences about the presence of Bt-derived toxins in aphids, sometimes even for the same species reared on the same host plant. Concentrations of Cry toxins in aphids were reported to be below the limit of detection in several studies and similarly we could not detect Cry3Bb toxin in aphids reared on Bt tomato. For instance, Raps et al. (Citation2001) did not detect Cry1Ab toxin in Rhopalosiphum padi (L.) (Hemiptera: Aphididae) after feeding on Bt-maize (event Bt11) in greenhouse. Similar results were obtained using MON810 maize by Head et al. (Citation2001) and Ramirez-Romero et al. (Citation2008), to cite a few. However, some researchers did find measurable amounts of Bt protein in aphids. For example, Dutton et al. (Citation2002) found trace amounts (0.02 µg g−1) in R. padi, and in cotton, in two different events, Zhang et al. (Citation2006) detected the toxin both in the aphid Aphis gossypii Glover (Hemiptera: Aphididae) (6 ng g−1) and in its predator ladybird, Propylaea japonica (Thunberg) (Coleoptera: Coccinellidae). Burgio et al. (Citation2007) obtained mixed results in the detection of Bt toxin in different strains of M. persicae (Sulzer) (Hemiptera: Aphididae) on oilseed rape reared under different conditions. More precisely, the aphids reared in the greenhouse were heavier than those reared in a growth chamber and were shown to contain the Cry1Ac toxin, while only in one sample out of eight aphids reared in the growth chamber was the toxin detectable. Romeis and Meissle (Citation2010) argued that false positive results might have been caused by contamination and/or extraction methods. However, Burgio et al. (Citation2011) repeated experiments with the same Cry1Ab-expressing oilseed rape in growth chamber with plants in a pre-flowering stage and again obtained several positive results.
The actual presence of Cry toxin in the vascular tissues of transformed plants clearly depends on the promoter used, but whether the toxin is translocated into phloem sap is controversial: it appears that the toxin is not detected when methods based on microcapillary techniques (Raps et al. Citation2001) or stilectomy are used (Faria et al. Citation2007), while, when leaf extraction (Raps et al. Citation2001; Dutton et al. Citation2002) or petiole exudation (Burgio et al. Citation2007) are performed, the presence of the toxin can be revealed. Since in one case, when Cry toxin was administered via artificial diet, activity against aphids was demonstrated (Porcar et al. Citation2009), it seems reasonable to expect a similar activity, should the same concentration of toxin be presented via the phloem sap.
The lack of effect of the Cry3Bb toxin on the biological parameters of M. euphorbiae detected in our experiments agrees with previous results recorded on different aphid/Bt-plant combinations (Head et al. Citation2001; Raps et al. Citation2001; Dutton et al. Citation2002; Lumbierres et al. Citation2004; Ramirez-Romero et al. Citation2008). However, for a finer analysis, we chose to also include in our investigations measurement endpoints other than mortality, as effects found in laboratory studies conducted with GM crops indicate that mortality is not the most sensitive parameter to detect such impacts (Lövei et al. Citation2009).
In other papers an increased performance of aphids on Cry-expressing plants was indicated. In a multigeneration study, A. gossypii performed better on Cry1A-expressing cotton than on untransformed controls with a visible impact on morphological traits (Liu et al. Citation2005). Similarly, R. padi performed better on Bt-maize in terms of fecundity with respect to control plants, though no final effect on r m was recorded (Lumbierres et al. Citation2004). An involvement of the different amino acid concentration and composition in the sap on aphid development was hypothesized by Faria et al. (Citation2007), who detected an enhanced aphid fitness on GM maize compared to the control.
Opposing effects were reported in another study, in which A. gossypii showed slightly better performances on conventional cotton (Zhang et al. Citation2008). The same study indicated higher mortality for the predator Orius sauteri (Poppius) (Hemiptera: Anthocoridae) when cotton plants expressing a fused Cry1Ac/Ab gene were observed in tritrophic experiments.
The feeding performances of M. caliginosus in our conditions were similar when their prey aphids were fed GM or control tomato plants. Considering that adult longevity was not impaired by the different diets, it is expected that feeding activity of this predator in GM agro-ecosystems should be maintained. Predators are considered generally less sensitive than parasitoids to toxins expressed in GM plants resistant to insects (Lövei et al. Citation2009; Naranjo Citation2009).
It must be considered that lab tests are extremely conservative as for the environmental risk on non-target organisms. As recently reported by Duan et al. (Citation2010), field studies with Coleopteran-active Cry proteins have either confirmed lab results or overestimated the possible negative effects. In this view, our results confirm that, at least for the species selected, the use of GM tomato in the field has no appreciable effects on non-target pests and antagonists. However, it remains to assess the effect of a long-term exposure to Cry proteins on each component of the trophic chain, at least for a period (months) compatible with the current cultivation procedures.
Among the possible effects of a genetic transformation, they must be considered either the pleiotropic effects or insertional mutagenesis (Maessen Citation1997), leading to an alteration of the volatile profile that affect the foraging behavior of insect antagonists. For example, the overexpression of a protein involved in the plant response to wounding resulted in a higher attractiveness of transformed tomato plants toward the parasitoid A. ervi (Corrado et al. Citation2007). In this view, we tried to assess the attractiveness for parasitoids of GM tomato plants with respect to control plants. No differences in the flight behavior of this parasitoid were found, and since the response of A. ervi remained unchanged, we therefore hypothesize that the volatile profile of the Cry3Bb expressing tomato plants was not different from conventional plants in relation to plant–parasitoid interactions, at least for those compounds involved in the foraging behavior of A. ervi (Sasso et al. Citation2009). This hypothesis was reinforced by the fact that the values of attractiveness recorded for our Bt tomato plants were very similar to those reported for different (not infested) cultivars of tomato, namely M82 (30%, Guerrieri et al. Citation2004), Better Boy (20%, Corrado et al. Citation2007) and UC82 (20%, Sasso et al. Citation2007). All these cultivars, except Better Boy, are significantly more attractive toward A. ervi when infested by M. euphorbiae following a quantitative alteration of the volatile compounds released (Sasso et al. Citation2007; Guerrieri et al. unpublished results).
In conclusion, the results of the present paper do not show any significant impact of tomato plants expressing the B. thuringiensis toxin Cry 3Bb on the biological performances of the non-target aphid M. euphorbiae and its natural enemies, M. caliginosus and A. ervi. In a scenario of a field deployment of genetically modified horticultural crops, where sap feeders are an important component of the agro-ecosystem (e.g. Arpaia et al. Citation2007), multi-trophic studies along their food chains may be particularly important to address. Other taxa are active in tomato fields as primary consumers feeding on aphids (e.g. Coleoptera Coccinellidae, Neuroptera Chrysopidae, and Hymenoptera Aphelinidae) and since the sensitivity to the toxins expressed in plants is very different among taxa (Lövei et al. Citation2009) and their exposure not completely clarified, the possible effects on population dynamics are not easy to be determined.
Acknowledgements
This project has been supported by MIUR (Project n. 32, Regioni Obiettivo 1) and by Italian CNR.
Additional information
Notes on contributors
Raffaele Sasso
†Current address: ENEA Centro Ricerche Casaccia UTAGRI – ECO via Anguillarese 301, 00123, Rome, ItalyReferences
- Arpaia , S , Di Leo , GM , Fiore , MC , Schmidt , JEU and Scardi , M . 2007 . Composition of arthropod species assemblages in Bt-expressing and near isogenic eggplants in experimental fields . Environ Entomol. , 36 : 213 – 227 .
- Beijerinck , MW and Van Delden , A . 1902 . Über die Assimilation des freien Stickstoffs durch Bakerien . Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Abteilung II , 9 : 3 – 43 .
- Birch , LC . 1948 . The intrinsic rate of natural increase of an insect population . J Anim Ecol. , 17 : 15 – 26 .
- Birch , ANE , Wheatley , R , Anyango , B , Arpaia , S , Capalbo , D , Getu Degaga , E , Fontes , E , Kalama , P , Lelmen , E Lövei , G . 2004 . “ Biodiversity and non-target impacts: a case study of Bt maize in Kenya ” . In Environmental risk assessment of transgenic organisms: a case study of Bt maize in Kenya , Edited by: Hilbeck , A and Andow , DA . 117 – 185 . Wallingford , , UK : CAB International Publishing .
- Burgio , G , Dinelli , G , Marotti , I , Zurla , M , Bosi , S and Lanzoni , A . 2011 . Bt-toxin uptake by the non-target herbivore, Myzus persicae (Hemiptera: Aphididae), feeding on transgenic oilseed rape in laboratory conditions . Bull Entomol Res. , 101 : 241 – 247 .
- Burgio , G , Lanzoni , A , Accinelli , G , Dinelli , G , Bonetti , A , Marotti , I and Ramilli , F . 2007 . Evaluation of Bt-toxin uptake by the non-target herbivore, Myzus persicae (Hemiptera: Aphididae), feeding on transgenic oilseed rape . Bull Entomol Res. , 97 : 211 – 215 .
- Conn , HJ . 1942 . Validity of the genus Alcaligenes . J. Bacteriol , 44 : 353 – 360 .
- Corrado , G , Sasso , R , Pasquariello , M , Iodice , L , Carretta , A , Cascone , P , Ariati , L , Digilio , MC , Guerrieri , E and Rao , R . 2007 . Systemin regulates both systemic and volatile signaling in tomato plants . J Chem Ecol. , 33 : 669 – 681 .
- Craig , M , Tepfer , M , Degrassi , G and Ripandelli , L . 2008 . An overview of general features of risk assessment of genetically modified crops . Euphytica , 164 : 853 – 880 .
- Duan , JJ , Lundgren , JG , Naranjo , S and Marvier , M . 2010 . Extrapolating non-target risk of Bt Crops from laboratory to field . Biol Lett. , 6 : 74 – 77 .
- Dutton , A , Klein , H , Romeis , J and Bigler , F . 2002 . Uptake of Bt-toxin by herbivores feeding on transgenic maize and consequences for predator Chrysoperla carnea . Ecol Entomol. , 27 : 441 – 447 .
- European Food Safety Authority (EFSA) . 2006 . Guidance document of the scientific panel on genetically modified organisms for the risk assessment of genetically modified plants and derived food and feed . EFSA J. 4:1–100 .
- European Food Safety Authority (EFSA) . 2008 . Environmental risk assessment of genetically modified plants – challenges and approaches . Parma , , Italy : EFSA Scientific Colloquium Summary Report , 20–21 June 2007 , 159 pp.
- Faria CA Wäckers FL Pritchard J Barrett DA Turlings TCJ. 2007 . High susceptibility of Bt maize to aphids enhances the performance of parasitoids of lepidopteran pests . PLoS ONE 2 7 , e600 . doi: 10.1371/journal.pone.0000600
- Guerrieri , E , Lingua , G , Digilio , MC , Massa , N and Berta , G . 2004 . Do interactions between plant roots and the rhizosphere affect parasitoid behaviour? New Perspectives . Ecol Entomol. , 29 : 753 – 756 .
- Guerrieri , E , Poppy , G , Powell , W , Rao , R and Pennacchio , F . 2002 . Plant to plant communication mediating in-flight orientation of Aphidius ervi . J Chem Ecol. , 28 : 1703 – 1715 .
- Head , G , Brown , CR , Groth , ME and Duan , JJ . 2001 . Cry1Ab protein levels in phytophagous insects feeding on transgenic corn: implications for secondary exposure risk assessment . Entomol Exp Appl. , 99 : 37 – 45 .
- Iannacone , R , Grieco , PD and Cellini , F . 1997 . Specific sequence modifications of a cry3B endotoxin gene result in high levels of expression and insect resistance . Plant Mol Biol. , 34 : 485 – 496 .
- Iannacone , R , Mittempergher , F , Morelli , G , Panio , G , Perito , A , Ruberti , I , Sessa , G and Cellini , F . 2008 . Influence of an Arabidopsis dominant negative athb2 mutant on tomato plant development . Acta Hort. (ISHS) , 789 : 263 – 276 .
- James C. 2009 . Global status of commercialized biotech/GM crops . ISAAA Brief No. 41 . Ithaca , NY : ISAAA .
- Kennedy , GG . 2003 . Tomato, pests, parasitoids, and predators . Annu Rev Entomol. , 48 : 51 – 72 .
- Lange , WH and Bronson , L . 1981 . Insect pests of tomatoes . Annu Rev Entomol. , 26 : 345 – 371 .
- Liu , XD , Zhai , BP , Zhang , XX and Zong , JM . 2005 . Impact of transgenic cotton plants on a non-target pest, Aphis gossypii Glover . Ecol Entomol. , 30 : 307 – 315 .
- Lövei , GL and Arpaia , S . 2005 . The impact of transgenic plants on natural enemies: a critical review of laboratory studies . Entomol Exp Appl. , 114 : 1 – 14 .
- Lövei , GL , Andow , DA and Arpaia , S . 2009 . Transgenic insecticidal crops and natural enemies: a detailed review of laboratory studies . Environ Entomol. , 38 : 293 – 306 .
- Lumbierres , B , Albajes , R and Pons , X . 2004 . Transgenic Bt maize and Rhopalosiphum padi (Hom., Aphididae) performance . Ecol Entomol. , 29 : 309 – 317 .
- Maessen , GDF . 1997 . Genomic stability and stability of expression in genetically modified plants . Acta Bot Neerl. , 46 : 3 – 24 .
- Nap , JP , Metz , PLJ , Escaler , M and Conner , AJ . 2003 . The release of genetically modified crops into the environment – Part I. Overview of current status and regulations . Plant J. , 33 : 1 – 18 .
- Naranjo , SE . 2009 . Impacts of Bt crops on non-target invertebrates and insecticide use patterns. CAB reviews: perspectives in agriculture . Vet Sci Nutr Nat Res. , 4 : 1 – 23 .
- Obrist , LB , Dutton , A , Albajes , R and Bigler , F . 2006 . Exposure of arthropod predators to Cry1Ab toxin in Bt maize fields . Ecol Entomol. , 31 : 143 – 154 .
- Porcar , M , Grenier , AM , Federici , B and Rahbé , Y . 2009 . Effects of Bacillus thuringiensis -endotoxins on the pea aphid (Acyrthosiphon pisum) . App Environ Microbiol. , 75 : 4897 – 4900 .
- Ramirez-Romero , R , Desneux , N , Chaufaux , J and Kaiser , L . 2008 . BT-maize effects on biological parameters of the non-target aphid Sitobion avenae (Homoptera: Aphididae) and Cry1Ab toxin detection . Pest Biochem Physiol. , 91 : 110 – 115 .
- Raps , A , Kehr , J , Gugerli , P , Moar , WJ , Bigler , F and Hilbeck , A . 2001 . Immunological analysis of phloem sap of Bacillus thuringiensis corn and of the nontarget herbivore Rhopalosiphum padi (Homoptera: Aphididae) for the presence of Cry1Ab . Mol Ecol. , 10 : 525 – 533 .
- Rohlf FJ Sokal RR. 1995 . Statistical tables, , 3rd ed . NY : W. H. Freeman and Co ., pp. 199 .
- Romeis , J , Bartsch , D , Bigler , F , Candolfi , MP , Gielkens , MMC , Hartley , SE , Hellmich , RL , Huesing , JE , Jepson , PC Layton , R . 2008 . Assessment of risk of insect-resistant transgenic crops to nontarget arthropods . Nat Biotechnol. , 26 : 203 – 208 .
- Romeis J, Meissle M. 2010 . Non-target risk assessment of Bt crops – cry protein uptake by aphids . J Appl Entomol . 135 : 1 – 6
- SAS . 1989 . SAS/STAT user's guide . Version 6, 4th ed ., Vol. 2 . Cary , NC : SAS Institute Inc ., pp. 846 .
- Sasso , R , Iodice , L , Digilio , MC , Carretta , A , Ariati , L and Guerrieri , E . 2007 . Host-locating response by the aphid parasitoid Aphidius ervi to tomato plant volatiles . J Plant Interact. , 2 : 175 – 183 .
- Sasso , R , Iodice , L , Woodcock , CM , Pickett , JA and Guerrieri , E . 2009 . Electrophysiological and behavioural responses of Aphidius ervi (Hymenoptera: Braconidae) to tomato plant volatiles . Chemoecology , 19 : 195 – 201 .
- Schmidt JEU . 2006 . The influence of transgenic plants expressing Bacillus thuringiensis d-endotoxins on arthropod diversity and trophic interactions in crop ecosystems . Dissertation ETH No.16554. Zurich, Switzerland .
- Sokal RR Rohlf FJ. 1981 . Biometry. , 2nd ed . NY : W. H. Freeman and Co ., pp. 859 .
- Walgenbrach , JF . 1997 . Effect of potato aphid (Homoptera: Aphididae) on yield, quality, and economics of staked-tomato production . J Econ Entomol. , 90 : 996 – 1004 .
- Young , JM , Kuykendall , LD , Martínez -Romero , E , Kerr , A and Sawada , H . 2001 . A revision of Rhizobium Frank 1889, with an emended description of the genus, and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R. rubi, R. undicola and R. vitis . Int J Syst Evol Microbiol , 51 : 89 – 103 .
- Zhang , GF , Wan , FH , Lövei , GL , Liu , WX and Guo , JY . 2006 . Transmission of Bt toxin to the predator Propylaea japonica (Coleoptera: Coccinellidae) through its aphid prey feeding on transgenic Bt cotton . Environ Entomol. , 35 : 143 – 150 .
- Zhang , GF , Wan , FH , Murphy , ST , Guo , JY and Liu , W-X . 2008 . Reproductive biology of two nontarget insect species, Aphis gossypii (Homoptera: Aphididae) and Orius sauteri (Hemiptera: Anthocoridae), on Bt and non-Bt cotton cultivars . Environ Entomol. , 37 : 1035 – 1042 .