
Abstract
Cydia molesta is an important pest of peach and pear fruits late in growing season. We identified and quantified volatiles from immature and mature fruits of peach and pear using gas chromatography-mass spectrometry. Antennally, active compounds were identified by gas chromatography-electroantennogram detection and were further tested in the field. Consistent electroantennographic activity was obtained for nine compounds from headspace collections of the mature fruits of two peach and two pear varieties. Esters were the major components, but qualitative and quantitative differences were found among four odor profiles. Blends mimicking pear fruit volatiles were more attractive to both sexes than blends mimicking peach fruit volatiles in both orchards. Our finding indicated that mixtures mimicking peach and pear fruit volatiles attracted both females and males of C. molesta, and these host volatiles may also be involved in the host switch behavior from peach to pear in mid-late season.
Introduction
Plant-derived volatiles play a significant role in guiding female of insect herbivores to their host plants for oviposition (Angioy et al. Citation2003; Casado et al. Citation2008; Cha et al. Citation2008; Bruce et al. Citation2011; Zhang et al. Citation2011; Yoneya & Takabayashi Citation2013). Insect herbivores with multiple generations per year are confronted with seasonal variations in the volatile organic compounds (VOCs) blends emitted by their different host plant species during the growing season (Piñero & Dorn Citation2009; Cha et al. Citation2010). Thus, the behavior of the insects must be adapted to respond to the natural variations of these blends (Tasin et al. Citation2007, Citation2009; Najar-Rodriguez et al. Citation2010, Citation2011). In particular, females that need to reproduce on different hosts are expected to modulate the response to the compounds ratios within distinct threshold values (Cha et al. Citation2010; Najar-Rodriguez et al. Citation2010, Citation2011). So, a comparison of the dynamics in volatile emission by different host plants of herbivores could provide further insight into the behavioral relationship of this insect herbivore with its different hosts.
The oriental fruit moth (OFM), Cydia (=Grapholitha) molesta (Busck) (Lepidoptera: Tortricidae), an oligophagous species with multiple generations per year, presumably originating from north-west China, has become widely distributed throughout the stone fruit-growing area in the world (Rothschild & Vickers Citation1991). All of its host plants belong to the family Rosaceae, including mainly peach, pear, apple, but also nectarine, plum, cherry, and others. In the early growing season, the moths first infest the growing peach and apple shoots. In the mid-late season, some moths stay in peach orchard and females lay eggs on mature peach fruits, and the others also can shift to mature pear fruits in the vicinity of peach trees (Makaji Citation1987; Zhao et al. Citation1989; Yang & Liu Citation2010; Lu et al. Citation2012). After peach harvest, a larger OFM population migrates into the pear orchard. In total, the populations show some seasonal dynamics in host plant choice, with switches from stone fruit orchards to pome fruit orchards from mid to late season (Myers et al. Citation2006). In China and some other Asian countries, inter-planting and mixed cultivation of pears and peaches is very common. Such a situation poses a serious threat to pear cultivation in the vicinity of peach trees (Zhao et al. Citation1989; Yang & Liu Citation2010). Pears are substantial post-peach host of OFM during the late fruit-growing season. What is the role of VOCs of peach and pear fruits in the peach-pear system? Such knowledge should be helpful to elucidate mechanisms underlying the host shift of OFMs from peach to pear.
Odor profiles released from peach trees at different phenological stages had been well studied, mainly peach shoots (Natale et al. Citation2003, Citation2004; Piñero & Dorn Citation2007; Il'ichev et al. Citation2009). However, little information is available on the seasonal dynamics of volatile emissions by peach and pear fruits (Lu et al. Citation2012). The objectives of the present study were (1) to identify and analyze the VOCs from peach and pear fruits, (2) to compare the variation between two plant tissues, and (3) to access potentially attractive VOCs to OFM in the field.
Materials and methods
Insects
The OFMs used in this study were obtained from an experimental orchard at the Institute of Forestry and Pomology (IFP), Beijing Academy of Agriculture and Forestry (BAAF), Beijing, China (39°58′N, 116°13′E). Larvae were collected from field-infested shoots of peach, Prunus persica L. Batsch cv. Dajiubao, in late June 2011 and 2012 and were maintained in a climatic chamber at 24 ± 1°C and 65–70% relative humidity (RH) under a photo:scoto regime of 16 L:8 D. Larvae were mass-reared on apple, Malus domestica L. Borkh. cv. Hongfushi, in a glass container (27 cm diameter, 13 cm height) for three instars and then transferred individually to smaller glass containers (2.5 cm diameter, 8 cm height) until eclosion. Adults were maintained in a bell-shaped glass container (diameter of the two openings 6 cm and 15 cm, 41 cm height). Both sides of the container were covered with fine nylon mesh. From a hole pierced through the mesh in the small side, 15% honey solution was provided on water-soaked cotton. The moths were reared in the laboratory for three generations before testing. For electrophysiological experiments, gas chromatography-electroantennogram detection (GC-EAD), 2- to 3-d-old females were used.
Chemicals
Decane (99%), tridecane (99%), tetradecane (99%), pentadecane (99%), hexadecane (99%), octadecane (99%), nonadecane (99%), 2-methylpropyl acetate (99%), 3-methylbutyl acetate (97%), ethyl pentanoate (98%), (Z)-3-hexenyl acetate (98%), methyl octanoate (99%), methyl salicylate (99%), and 6-methyl-5-hepten-2-one (98%) were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA). Heptadecane (99%), 1-hexanol (99%), 2-ethyl-1-hexanol (99%), octanal (98%), nonanal (97%), decanal (97%), hexyl acetate (99%), limonene (98%), and racemic linalool (97%) were obtained from Fluka Production GmbH (Buchs, Switzerland). Ethyl butanoate (99%), butyl acetate (99%) were bought from Acros Organics (NJ, USA). Ethyl hexanoate (98%) was purchased from Tokyo Chemical Industry Co. (Tokyo, Japan). Compounds that did not elicit antennal responses, and for which no standards were available, were tentatively identified using the NIST-database.
Plant materials
Peach and pear were cultivated in an experimental orchard at the IFP. The peach varieties were Common peach (CP) from P. persica (L.) Batsch cv. Shenzhoubaimi and Flat peach (FP) of P. persica (L.) cv. Wuyuexianbiangan. The two pear varieties were Xiangli (XL) of Pyrus bretschneideri and Xingao (XG) from Pyrus pyrifolia. All of these varieties were commonly grown in the Beijing area. No insecticides or any specific treatment against OFM were used in the orchard during the tests. Unripe and ripe fruits were healthy and were picked from trees according to the phenological development of corresponding varieties in the Beijing area in 2011 and 2012 (Cao et al. Citation2000, Citation2006; Jiang et al. Citation2002; Yin Citation2005). The fruits were picked within 20 min before sampling of VOCs.
Collection of VOC
We used a push–pull system to collect headspace VOCs. Fruits (ca. 1500 g) were placed immediately after harvesting into a 2000 ml glass jar for extraction. Air aspirated with a vacuum pump (Qianxi Air Company, Beijing, China) was filtered through an activated-charcoal filter, then passed through the jar at 300 ml/min, and finally through a sorbent cartridge (Porapak Q, 50 mg, 80/100 mesh, Supelco, Bellefonte, PA, USA). The sorbents were held between plugs of glass wool in a glass tube (10 cm long, 0.5 cm inner diameter). The samples were collected for 8 h at 24 ± 1°C and 65–70% RH. Volatiles were desorbed by eluting the sorbent cartridge with 500 µl hexane (HPLC grade, Sigma-Aldrich) at room temperature. Five samples were collected for each unripe and ripe varieties, respectively. The samples were analyzed by GC-EAD and gas chromatography-mass spectrometry (GC-MS). For quantitative analyses, 0.5 µg benzaldehyde (99%, Fluka, Switzerland) was added as an internal standard to each sample. Preliminary analyses showed that this compound was not detectable in the headspace of the pear and peach fruits studied here. Sample volumes were reduced to 50 µl using a slow stream of nitrogen and then analyzed. If not used immediately, extracts were sealed in glass vials and stored at −18°C until used.
GC-MS
Headspace VOCs of the unripe and ripe fruits of the peach and pear varieties and mixtures of synthetic compounds were analyzed with an Agilent Technologies 5973 MS (Agilent) coupled to an Agilent Technologies 6890 N GC (Agilent) equipped with polar DB-WAX fused-silica column (30 m × 0.25 mm ID, 0.25 µm film, J&W Scientific Inc., Folsom, CA, USA) or nonpolar DB-5 fused-silica column (30 m × 0.25 mm ID, 0.25 µm film, J&W Scientific Inc., Folsom, CA, USA). The temperature program was as follows: 50°C for 1 min, then 3°C/min to 120°C, then 10°C/min to 240°C and finally held at 240°C for 10 min. Windows NT/MASS Spectral Search Program (Version 1.7) software was used for data analysis. Injections were made in the splitless mode. Helium was used as the carrier gas (1.0 ml/min). For electron impact mass spectra, the ionization voltage was 70 eV, and the temperatures of the ion source and of the interface were 230°C and 280°C, respectively. The emission current was 34.6 µA. Identification of VOCs was verified by comparison with authentic samples.
GC-EAD
VOCs were analyzed with an Agilent Technologies 6890 N GC with a flame ionization detector (FID), using a polar DB-WAX (30 m × 0.25 mm ID, 0.25 µm film, J&W Scientific, Folsom, CA, USA) or nonpolar DB-5 (30 m × 0.25 mm ID, 0.25 µm film, J&W Scientific Inc., Folsom, CA, USA) capillary column coupled with an EAD (Syntech, Hilversum, The Netherlands). The oven temperature program was the same as in the GC-MS analysis. Nitrogen was used as the carrier gas (1.0 ml/min). A micromanipulator assembly (MP-15, Syntech) was connected to a stimulus controller (CS-55, Syntech). All signal sources were connected to a serial data acquisition interface (IDAC-4, Syntech). The antennae from the heads of OFM females were excised using micro-scissors. A few segments from the tips of antennae were clipped off and mounted on the antenna holder with two metal electrodes using conductive gel (Spectra 360, Parker Lab, NJ, USA), and then the electrode holder was inserted into the EAD probe. Only one antenna was used at the same time. Testing began after a relative stable baseline had been achieved. The outlet of the GC column was split in a 2:1 ratio between the EAD, a cut antenna of an OFM female, and the FID. The mounted antenna was positioned in the charcoal-filtered and humidified air stream that carried the VOCs eluting from the GC column. The antennal and FID signals were amplified and recorded simultaneously using Syntech software (GC-EAD 32, version 4.4, Syntech, Hilversum, The Netherlands). Identities of EAD-active compounds were verified by comparison of mass spectra and retention times with those of synthetic standards. Each sample was tested six times on six different antennae. Each tested antenna was derived from a different female.
In addition, a blend of nine EAD-active synthetic compounds (1-hexanol, nonanal, ethyl butanoate, butyl acetate, 3-methylbutyl acetate, ethyl hexanoate, hexyl acetate, (Z)-3-hexenyl acetate, and linalool) was analyzed by GC-EAD (10 ng of each).
Field experiment
On the basis of the results of the GC-EAD analyses, VOCs from the mature fruits of two peach and two pear varieties eliciting antennal responses in female OFM were formulated in blends for the field tests. Four blends of synthetic compounds were prepared in the ratios of GC-EAD-active VOCs as emitted by the corresponding varieties (). Sleeve-type rubber septa (10 mm depth, 6 mm inter diameter, ca. 400 µl) purchased from Shunyi Rubber Company, Beijing, China, were used as dispensers. The formulations contained 100 mg of the most abundant compound and the others in the same proportion as in the natural volatile mixture. Preliminary field experiments showed that such concentrations can provide an adequate emission under field conditions. The mixtures of compounds were prepared 1–2 h prior to bioassays in the field trial. They were added into the rubber septa and then fixed upward on the bottom of sticky delta trap in the dusk, during which the adults of OFM are most active, for all the treatments. Unbaited traps (HPLC-grade hexane, Sigma-Aldrich) and OFM sex-pheromone lures (Geruibiyuan Technology Company, Beijing, China) were used as controls.
Table 1. Components and blend ratios for each blend used in the field experiment.
The field test was conducted in a 7-hectare peach orchard and a 6.5-hectare pear orchard at the IFP with a history of OFM infestation. A sticky delta trap (35 cm long × 20 cm high × 20 cm wide, Geruibiyuan Technology Company, Beijing, China) was used to trap the insects. The trials were carried out in a randomized complete-block design. Blocks were at least 120 m apart. In each block, baited traps were installed approximately 1.5 m above the ground and were set up at a distance of at least 30 m to minimize interference between traps. Each treatment was replicated four times in four blocks.
The traps were monitored twice weekly. Captured OFMs were transferred to the laboratory, and their number and sex were recorded. The trials lasted 2 weeks. The data of captures over a 14-d period were pooled. The field test was conducted from 15 to 28 August 2012 in peach and pear orchards simultaneously, during mid-late fruits phonological stages for both peach and pear.
Data analysis
The antennal responses of C. molesta females to 10 ng of EAD-active standard compounds, and mean numbers of OFM males and females captured in traps baited with different lures in the field were analyzed by one-way ANOVA. The means were separated by Tukey's multiple range tests (P < 0.05). All data were analyzed with the statistical program SPSS (version 16.0).
Results
Chemical identification of VOCs from different pear and peach varieties
Several compounds were identified by GC-MS analysis from different pear and peach varieties, belonging to different chemical classes: hydrocarbons, alcohols, aldehydes, esters, benzene derivatives, ketones, and terpenoids ().
Table 2. Relative quantities of volatile compounds collected in the headspace of immature and mature fruits of pear and peach.
Significant difference was found between pear and peach VOCs, but the profiles of the two varieties of either pear or peach were similar to each other. The two varieties consisted of the same chemical classes, but differed greatly between ripe and unripe fruits.
For unripe pear fruits, (E)-2-nonen-1-ol, octanal, decanal, and 2-methylpropyl acetate were characteristic and absent from ripe fruits, whereas the concentrations of other VOCs, such as several hydrocarbons, 2-ethyl-1-hexanol, nonanal, and 6-methyl-5-hepten-2-one decreased with maturation. For unripe peach fruits, octanal, decanal, methyl salicylate, and limonene were characteristic and absent from ripe fruits, whereas the concentrations of other VOCs, such as linolool and 2-ethyl-1-hexanol decreased with maturation.
In mature pear fruits, 1-hexanol, 2-methyl-1-hexadecanol, 3-methyl-1-butyl acetate, ethyl pentanoate, ethyl hexanoate, hexyl acetate, methyl octanoate, and hexyl hexanoate were characteristic and absent from unripe fruits. In mature peach fruits, (E)-2-nonen-1-ol, 2-methylpropyl acetate, ethyl butanoate, butyl acetate, 3-methyl-1-butyl acetate, ethyl pentanoate, ethyl hexanoate, hexyl acetate, methyl octanoate, and hexyl hexanoate were characteristic and absent from unripe fruits.
Esters were the dominant VOCs in ripe fruits, and the concentrations increased with maturation. In particular, butyl acetate and ethyl butanoate were the most abundant VOC identified in XL and XG varieties, respectively. (Z)-3-hexenyl acetate was the most abundant VOC identified in both peach varieties ().
Antennal responses to VOCs from ripe pear and peach fruits
All the EAD-active compounds consist of four classes: alcohols, aldehydes, esters, and terpenoids. Hydrocarbons, benzenoids, and ketones cannot elicit the antennal response. In total, nine compounds from the headspace of ripe fruits belonging to two pear varieties and two peach varieties elicited antennal responses from OFM females: 1-hexanol, nonanal, ethyl butanoate, butyl acetate, 3-methylbutyl acetate, ethyl hexanoate, hexyl acetate, (Z)-3-hexenyl acetate, and linalool ().
The EAD-active VOCs were different in quality and quantity for both pear and peach varieties. Only nonanal was common components among four varieties. 1-hexanol, ethyl butanoate, butyl acetate, 3-methylbutyl acetate, hexyl acetate, and ethyl hexanoate were present in pear fruits volatiles, while linalool and (Z)-3-hexenyl acetate were present in peach fruits volatiles.
In total, different esters were frequently present among GC-EAD-active volatiles in the mature fruits of all varieties. All EAD-active esters in ripe fruits were either characteristic or dominant with maturation. Although ethyl butanoate, butyl acetate, 3-methylbutyl acetate, hexyl acetate, and ethyl hexanoate also existed in peach variety FP, these compounds could not elicit female antennal response due to the low concentration. In contrast, these components in the headspace samples of the pear variety XG did elicited electroantennography (EAG) response (; ).
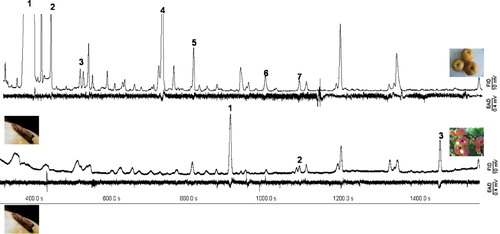
In order to verify and quantify the EAG response, a second series of GC-EAD analyses was performed using 10 ng of each VOC. The strongest antennal response in OFM mated females was evoked by linalool ().
Table 3. Antennal responses of Cydia molesta females to synthetic compounds identified from mature pear and peach fruit as determined by GC-EAD.
Field experiment
In peach orchard, the traps baited with the blends mimicking the odor of pear variety XG (1-hexanol, nonanal, ethyl butanoate, butyl acetate, 3-methylbutyl acetate, ethyl hexanoate, and hexyl acetate with a 3:1:100:8:4:12:4 ratio) and pear variety XL (nonanal, ethyl butanoate, and butyl acetate with a 3:1:100 ratio) caught significantly more OFM males than the control and the traps baited with the blends mimicking the odor of peach variety FP (nonanal, (Z)-3-hexenyl acetate, and linalool with a 24:100:35 ratio) and peach variety CP ((Z)-3-hexenyl acetate and linalool with a 100:1 ratio). Only few females were trapped with no significant difference between traps, including the control ().
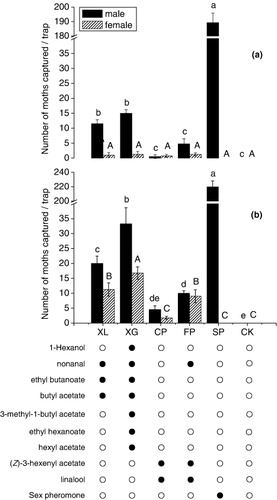
A larger OFM population apparently was present in the pear orchard than in the peach orchard, based on the number of male moths trapped with the sex pheromone (, ). As in the field test in peach orchard, traps baited with mixtures of the XL and XG varieties caught significantly more OFMs than the control, and the traps baited with the other two peach mixtures, but the total captures, females in particular, were significantly greater in pear orchard than peach orchard (, ).
Discussion
The OFM, Cydia (=Grapholitha) molesta (Busck) (Lepidoptera: Tortricidae), is an oligophagous species with multiple generations per year. In North China, the moths first infest the growing peach and apple shoots in the early growing season. In the mid-late season, populations show seasonal dynamics, with switch from peach to pear orchards, mainly after fruit ripening. Ripening pear or peach fruits offer a chemical environment that is attractive to the moths late in the season. Olfactory cues from these plant tissues should play an important role in orientation to the primary and secondary host plants (Piñero & Dorn Citation2009; Najar-Rodriguez & Dorn Citation2013).
We identified the VOCs from mature pear and peach fruits and nine EAD-active VOCs were found in our study. We found the chemical background offered by fruits presented the seasonal dynamics as to fruit maturity and different plant species. In the further research, we compared the attraction of mature fruit volatiles from the two different plant species. The results indicated that both peach and pear fruit volatiles were attractive to OFM; however, the lures mimicking pear variety always caught more OFM than the control and the lures mimicking peach variety in both orchards. So we think that stronger attraction of both sexes to ripening pear fruits could be a crucial factor to host shifts of OFM from mid to late season. A larger OFM population based on the number of male moths trapped with the sex pheromone in the pear orchard than in the peach orchard suggested that seasonal migration from peach to pear tree was in progress during our experimental stage.
The odor profiles perceived by C. molesta during the whole phenological development vary. First, at the beginning of the season, it mainly infests the growing shoots of peach. Natale et al. (Citation2003) reported a three-component mixture of (Z)-3-hexenyl acetate, (Z)-3-hexenol, and benzaldehyde from peach shoots in a 4:1:1 ratio elicited a significant bioactivity in dual choice area by a bioassay-assisted fractionation using different sorbent polymers. Piñero and Dorn (Citation2007) found the five-compound mixture of benzaldehyde, benzonitrile, (Z)-3-hexenyl acetate, (Z)-3-hexenol, and (E)-2-hexenal from peach shoots was as attractive as natural volatiles in a Y-tube olfactory meter using subtraction approach (Piñero & Dorn Citation2007). In field test, Il'ichev et al. (Citation2009) reported (Z)-3-hexenyl acetate, (E)-β-farnesene, and (E)-β-ocimene in proportion 1:2:2 were the best lures (Il'ichev et al. Citation2009). Second, in mid-late season, it mainly infests fruits of peach and pear. Lu et al. (Citation2012) identified and evaluated the pear fruit volatiles attractive to the OFM. The mixture of 1-hexanol, nonanal, ethyl butanoate, butyl acetate, ethyl hexanoate, hexyl acetate, hexyl butanoate, and farnesene (different isomers) with a 1:1:100:70:7:5:1:4 ratio from the variety JM and the mixture of nonanal, ethyl butanoate, 3-methylbutyl acetate, ethyl hexanoate, hexyl acetate, and farnesene with a 1:100:1:32:1:2 ratio from the variety HJ were highly attractive to both sexes in the field. In the study, the mixture of 1-hexanol, nonanal, ethyl butanoate, butyl acetate, 3-methylbutyl acetate, ethyl hexanoate, and hexyl acetate with a 3:1:100:8:4:12:4 ratio and the mixture of nonanal, linalool, and (Z)-3-hexenyl acetate with a 24:100:35 ratio were more attractive VOCs from mature pear and peach fruits to OFM in the field, respectively.
Taken together, esters were the most crucial volatiles to OFMs during mid-late season. Headspace volatiles from mature fruits were greatly different from shoot VOCs. Esters were absolutely the most abundant components from mature fruits, and several of them elicited antennal responses. We also found the increase in ester production in mature pear fruits corresponded with pest damage in mature fruit. The seasonal flight period of OFM starts before the onset of pear-fruit ripening, and females oviposit on the mature pear fruit (Lu et al. Citation2012). All these findings suggest that esters are essential for seasonal host switch during the mid-late fruit-growth stage. In other words, plasticity exists in the host plant recognition system of OFMs. Although qualitative and quantitative differences were found between the VOCs of peach shoots and fruits of pear and peach, all VOCs were attractive to the moths, verifying the plasticity of the host plant recognition in OFM. Similar results had been found in several herbivore species (Cha et al. Citation2008, Citation2010). Some multivoltine insects with multiple generations per year such as the OFM exploit several plant species as their hosts in different phenological stages. This is an important strategy for optimization of resources in order to complete the life cycle and is important for understanding the evolution of VOC-based recognition of host plants by herbivorous insects and for development of a possible semiochemical-based synthetic lure for use in the field (Tasin et al. Citation2005, Citation2009; Falcãoa et al. Citation2013).
Although the lures were based on the EAD-active compounds for females, male captures were higher, which had been reported (Il'ichev et al. Citation2009; Lu et al. Citation2012). Similar results also were found for Cydia pomonella, an important apple borer (Coracini et al. Citation2004). Why are more male OFMs than females captured by traps with lures emitting plant-derived VOCs? First, the flight mode of both sexes toward host plants may be different. Males fly toward the plant in search of females, and their orientation behavior is adapted to locate a point source of sex pheromone. By contrast, females fly toward the plant in search of suitable oviposition sites. The flight mode of both sexes toward host plants should be an important factor for contacting on the traps. Second, physical structure could also affect the landing of females. In our study, OFMs always laid eggs on the smooth surface of waxed paper in the laboratory. The sticky base of the traps might be a repellent for female landing; therefore, trap design could be quite important in further research. Nevertheless, the high male capture rates in our study could be applied to establish a novel approach for monitoring males, especially in orchards treated with sex pheromone for mating disruption (Il'ichev et al. Citation2009; Lu et al. Citation2012).
Acknowledgments
We thank Fang Xu in our laboratory for their technical assistance in GC-EAD and GC-MS. We also thank Elina Verbon, Wageningen University, for their help in improving the paper. This work was supported by the National Natural Science Foundation of China [31270693, 81102747].
Related Research Data
References
- Angioy AM, Desogus A, Barbarossa IT, Anderson P, Hansson BS. 2003. Extreme sensitivity in an olfactory system. Chem Senses. 28:279–284. 10.1093/chemse/28.4.279
- Bruce TJA, Martin JL, Smart LE, Pickett JA. 2011. Development of semiochemical attractants for monitoring bean seed beetle, Bruchus ruimanus. Pest Manag Sci. 67:1303–1308. 10.1002/ps.2186
- Cao YF, Li SL, Huang LS, Sun BJ, Tan XW. 2000. Comprehensive evaluation of pear variety resource in China. China Fruit. 4:42–44.
- Cao YF, Liu FZ, Hu HJ, Zhang BB. 2006. Descriptors and data standard for pear (Pyrus spp.). Beijing: China Agriculture Press.
- Casado D, Gemeno C, Avilla J, Riba M. 2008. Diurnal variation of walnut tree volatiles and electrophysiological responses in Cydia pomonella (Lepidoptera: Tortricidae). Pest Manag Sci. 64:736–747. 10.1002/ps.1551
- Cha DH, Nojima S, Hesler SP, Zhang AJ, Linn Jr CE, Roelofs WL, Loeb GM. 2008. Identification and field evaluation of grape shoot volatiles attractive to female grape berry moth (Paralobesia viteana). J Chem Ecol. 34:1180–1189. 10.1007/s10886-008-9517-0
- Cha DH, Linn Jr CE, Teal PEA, Zhang AJ, Roelofs WL, Loeb GM. 2010. Eavesdropping on plant volatiles by aspecialist moth: Significance of ratio and concentration. PLoS One. 6: e17033. 10.1371/journal.pone.0017033
- Coracini M, Bengtsson M, Liblikas L, Witzgall P. 2004. Attraction of coding moth males to apple volatiles. Entomol Exp Appl. 110:1–10. 10.1111/j.0013-8703.2004.00124.x
- Falcãoa JCF, Dáttilob W, Izzoa TJ. 2013. Temporal variation in extrafloral nectar secretion in different ontogenic stages of the fruits of Alibertia verrucosa S. Moore (Rubiaceae) in a Neotropical savanna. J Plant Interact. doi:10.1080/17429145.2013.782513
- Il'ichev AL, Kugimiya S, Williams DG, Takabayashi J. 2009. Volatile compounds from young peach shoots attract males of oriental fruit moth in the field. J Plant Interact. 4:289–294. 10.1080/17429140903267814
- Jiang WB, Gao GL, Yu KJ, Ma K. 2002. Creation of pear variety resource in China over the past decade and its prospects. J Fruit Sci. 19:314–320.
- Lu PF, Huang LQ, Wang CZ. 2012. Identification and field evaluation of pear fruit volatiles attractive to the oriential fruit moth, Cydia molesta. J Chem Ecol. 38:1003–1016. 10.1007/s10886-012-0152-4
- Makaji T. 1987. Diagnosis of fruit tree diseases and pests with color plates. Tokyo: Agriculture Publication Incorporation.
- Myers CT, Hull LA, Krawczy KG. 2006. Effects of orchard host plants on preference of the oriental fruit moth (Lepidoptera: Tortricidae). J Ecol Entomol. 99:1176–1183. 10.1603/0022-0493-99.4.1176
- Najar-Rodriguez AJ, Dorn SA. 2013. Season-long volatile emissions from peach and pear trees in situ, overlapping profiles, and olfactory attraction of an oligophagous fruit moth in the laboratory. J Chem Ecol. 39:418–429. 10.1007/s10886-013-0262-7
- Najar-Rodriguez AJ, Galizia CG, Stierle J, Dorn S. 2010. Behavioral and neurophysiological responses of female oriental fruit moths to changing ratios of a key component in a bioactive mixture. J Exp Biol. 213:3388–3397. 10.1242/jeb.046284
- Najar-Rodriguez AJ, Galizia CG, Stierle J, Dorn S. 2011. Behavioral and neurophysiological responses of female oriental fruit moths to changing ratios of a key component in a bioactive mixture. J Exp Biol. 214:162. 10.1242/jeb.054262
- Natale D, Mattiacci L, Hern A, Pasqualini E, Dorn S. 2003. Response of female Cydia molesta (Lepidoptera: Tortricidae) to plant derived volatiles. Bull Entomol Res. 93:335–342. 10.1079/BER2003250
- Natale D, Mattiacci L, Pasqualini E, Dorn S. 2004. Apple and peach fruit volatiles and the apple constituent butyl hexanoate attract female oriental fruit moth, Cydia molesta, in the laboratory. J Appl Entomol. 128:335–342.
- Piñero JC, Dorn S. 2007. Synergism between aromatic compounds and green leaf volatiles derived from the host plant underlies female attraction in the oriental fruit moth. Entomol Exp Appl. 125:185–194. 10.1111/j.1570-7458.2007.00614.x
- Piñero JC, Dorn S. 2009. Response of female oriental fruit moth to volatiles from apple and peach trees at three phenological stages. Entomol Exp Appl. 131:67–74. 10.1111/j.1570-7458.2009.00832.x
- Rothschild GHL, Vickers RA. 1991. Biology, ecology and control of the oriental fruit moth. In: Van der Geest LPS, Evenhuis HH, editors. Tortricid pests: Their biology, natural enemies and control. Amsterdam: Elsevier; p. 389–412.
- Tasin M, Anfora G, Ioriatti C, Carlin S, De Cristofaro A. 2005. Antennal and behavioral responses of grapevine moth Lobesia botrana females to volatiles from grapevine. J Chem Ecol. 31:77–87. 10.1007/s10886-005-0975-3
- Tasin M, Bäckman AC, Anfora1 G, Carlin S, Ioriatti C, Witzgall P. 2009. Attraction of female grapevine moth to common and specific olfactory cues from 2 host plants. Chem Senses. 35:57–64. 10.1093/chemse/bjp082
- Tasin M, Bäckman AC, Coracini M, Casado D, Ioriatti C, Witzgall P. 2007. Synergism and redundancy in a plant volatile blend attracting grapevine moth females. Phytochemistry. 68:203–209. 10.1016/j.phytochem.2006.10.015
- Yang XY, Liu JL. 2010. The reason of serious damage of Grapholitha molesta in 2008. Shanxi Fruit. 1:38.
- Yin ZH. 2005. Phylogenetic relationship among peaches and nectarines (Prunus Persica L. Batsch) based on SSR markers and ITS sequences [dissertation]. Beijing: China Agricultural University.
- Yoneya K, Takabayashi JK. 2013. Interaction–information networks mediated by plant volatiles: A case study on willow trees. J Plant Interact. 8:197–202. 10.1080/17429145.2013.782514
- Zhang QH, Ma JH, Yang QQ, Byers JA, Klein MG, Zhao FY, Luo YQ. 2011. Olfactory and visual responses of the long-legged chafer Hoplia spectabilis Medvedev (Coleoptera: Scarabaeidae) in Qinghai Province, China. Pest Manag Sci. 67:162–169. 10.1002/ps.2045
- Zhao ZR, Wang YG, Yan GY. 1989. The report of primary study on Grapholitha molesta in north of Jiangshu Province. Entomol Knowl. 26:17–19.