
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
In this paper, we study the periodic and stable dynamics of an interactive wild and sterile mosquito population model with density-dependent survival probability. We find a release amount upper bound , depending on the release waiting period T, such that the model has exactly two periodic solutions, with one stable and another unstable, provided that the release amount does not exceed
. A numerical example is also given to illustrate our results.
1. Introduction
Mosquitoes have been listed as one of the deadliest animals around the world, due to their ability of transmitting mosquito-borne diseases through female mosquitoes' bitings. Take dengue as an example, the number of cases reported by World Health Organization increased dramatically over the last two decades, from 505,430 cases in 2000, to over 2.4 million in 2010, and 5.2 million in 2019 [Citation30]. Since there have neither safe vaccines nor effective drugs to combat dengue, biological methods including Sterile Insect Technique (SIT) and Incompatible Insect Technique (IIT) have played vital roles in preventing the transmission of mosquito-borne diseases [Citation1–5,Citation8,Citation11,Citation19,Citation27–29]. Both SIT and IIT involve releasing sterile male mosquitoes into the wild to mate with wild females, which result in no progeny for those females. It occurs that the number of wild mosquitoes will decrease gradually.
As sustainable and biologically safe approaches, the implementations of SIT, IIT and combined IIT-SIT for controlling wild mosquitoes are really acceptable and generalized. How to employ ideas of modelling to study the field has attracted great attention of many researchers, and various mathematical models have been formulated, we may refer to [Citation6,Citation20,Citation21,Citation32,Citation34,Citation38,Citation39] for ordinary differential equations, [Citation14,Citation16,Citation18,Citation31,Citation33,Citation37] for delay differential equations, [Citation9,Citation15,Citation17,Citation22,Citation23] for reaction-diffusion equations, [Citation12,Citation13] for stochastic equations, [Citation25,Citation26,Citation35,Citation36,Citation40] for difference equations and references therein.
Based on a thought-provoking modelling idea proposed in [Citation31] that the number of sterile mosquitoes at time t can be treated as a given function rather than an independent variable constrained by a differential equation, and by taking the fact that density-dependent death of wild mosquitoes mainly occurs in the aquatic stages (eggs, larvae, pupae)[Citation7,Citation24] into consideration, the authors in [Citation21] studied the following ordinary differential equation model
(1)
(1)
for exploring the interactive dynamics of wild and sterile mosquitoes. In model (Equation1
(1)
(1) ),
and
denote the numbers of wild and sterile mosquitoes at time t, respectively, a is the maximum number of survived offspring produced per individual,
is the mating probability between wild mosquitoes, ξ denotes the carrying capacity parameter such that
describes the density-dependent survival probability, and μ is the natural mortality of wild mosquitoes.
Although model (Equation1(1)
(1) ) looks simple, we find that it can generate rich dynamics. Obviously,
plays a key role in determining the asymptotic behaviours of model (Equation1
(1)
(1) ), it is very important for us to make clear the structure of
. To this end, let
and T be the sexual lifespan of released sterile mosquitoes and the waiting period between two consecutive releases, respectively. Then there exists three different release strategies:
,
, and
. Moreover, we assume that the release begins at time t = 0 such that
for t<0, and a constant amount c of sterile mosquitoes is impulsively and periodically released at discrete time points
. When
,
for all
. And model (Equation1
(1)
(1) ) with
has been investigated in [Citation21]. For the case when
, we have
and model (Equation1
(1)
(1) ) becomes
which has recently been discussed in [Citation43], where
.
We study the release strategy of in this paper. Let
represent the largest integer portion of x. Then there exists an integer
and a nonnegative number
such that
. When
, we get
When
, if r = 0, then
, under which model (Equation1
(1)
(1) ) can be reduced to the one in [Citation21]. If
, then
(2)
(2)
See Figure for illustration. Then model (Equation1
(1)
(1) ) reads as
Figure 1. A schematic illustration for figuring out the structure of .
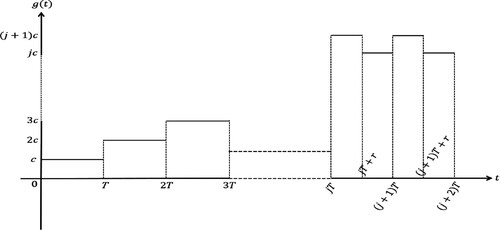
We assume 0<r<T in the remaining of this paper. Since our main concern is the asymptotic dynamics of the model, we may assume that the initial time point is . Then
takes the form of (Equation2
(2)
(2) ).
Let the release amount upper bound be defined as
Then model (Equation1
(1)
(1) ) can be rewritten as
(3)
(3)
.
We, in this paper, mainly study the long-time dynamics of model (Equation3(3)
(3) ) under the assumptions of
and
. The paper is arranged as follows. In Section 2, we define the Poincaré map and give three lemmas. In Section 3, the theoretical results show that if
, then the model has exactly two T-periodic solutions, with one stable and another unstable. Finally, a brief discussion is given in Section 4.
2. Preliminaries
It is apparent that the origin, denoted by , is the unique equilibrium of model (Equation3
(3)
(3) ). We first give the definition of a solution of model (Equation3
(3)
(3) ). A function
is said to be a solution of model (Equation3
(3)
(3) ) if it satisfies the first equation of model (Equation3
(3)
(3) ) on
, and the second equation of model (Equation3
(3)
(3) ) on
[Citation10]. Let
denote the solution of model (Equation3
(3)
(3) ) with initial value
. Furthermore, we define
and
for
. Then
is a continuous and piecewise differentiable function defined on
.
Set the two auxiliary functions and
be defined as
Clearly, both
and
are continuous and differentiable on
. A T-periodic solution occurs if
, i.e.
. Consequently, existence of fixed points of h ensures existence of T-periodic solutions of model (Equation3
(3)
(3) ). For characterizing the asymptotic behaviours of solutions, we define sequences
and
by
(4)
(4)
which satisfy
for
, see [Citation34] for details. Following the lines in [Citation34], for sequences defined in (Equation4
(4)
(4) ), we may similarly obtain the next lemma.
Lemma 2.1
For any given initial value u>0, the following conclusions hold.
| (1) | If | ||||
| (2) | If | ||||
| (3) | If | ||||
By a similar argument to that in the proof of Lemma 2.9 in [Citation32], we arrive at the following lemma, which provides a necessary and sufficient condition for the origin to be asymptotically stable.
Lemma 2.2
The origin is asymptotically stable if and only if there is
such that
Since our focus is to find the fixed points of h, we need to know the different relationships between and u for all u>0. Moreover, from Lemmas 2.1 and 2.2, we find that the sign of
is crucial to the comparison between
and u. To obtain the expression for
, we will first solve the first equation of model (Equation3
(3)
(3) ) with initial value
for getting the expression of
, then solve the second equation of model (Equation3
(3)
(3) ) with initial value
to get the relationship between
and
. To go ahead without the burden of frequently switching between the first equation and the second equation of model (Equation3
(3)
(3) ), we rewrite model (Equation3
(3)
(3) ) as follows
(5)
(5)
with k = j + 1 when
, or k = j when
, where
, and
.
Since , we know that
has two positive real roots. Denote the two roots by
and
, then
Moreover, we have
(6)
(6)
By letting
in Lemma 2.2, and using (Equation3
(3)
(3) ) and (Equation6
(6)
(6) ), we have the following lemma.
Lemma 2.3
The origin is always locally asymptotically stable.
Biologically, Lemma 2.3 means that the suppression on wild mosquitoes is always successful provided that the initial density of wild mosquitoes is contained within the region of attraction of .
Recall that the sign of is our deepest concern, and to get this sign, it suffices to obtain the expression for
. In the following, we are devoted to seeking the expression. To this end, we need to consider the following two cases.
Case 1: . In this case, Equation (Equation5
(5)
(5) ) can be transformed to
(7)
(7)
By partial fraction decomposition of
, (Equation7
(7)
(7) ) gives
(8)
(8)
where
Making the identity transformation for (Equation8
(8)
(8) ), we get
(9)
(9)
Integrating (Equation9
(9)
(9) ) from jT to jT + r with k = j + 1, and recall that
(10)
(10)
we have
(11)
(11)
For notation simplicity, let
(12)
(12)
Taking the partial derivative with respect to u of both sides of (Equation12
(12)
(12) ), we get
(13)
(13)
Furthermore, substituting (Equation12
(12)
(12) ) and k = j + 1 into (Equation11
(11)
(11) ), we have
(14)
(14)
To get the expression for
, we take the derivative with respect to u of both sides of (Equation14
(14)
(14) ), which yields
(15)
(15)
Substituting (Equation13
(13)
(13) ) with k = j + 1 and (Equation14
(14)
(14) ) into (Equation15
(15)
(15) ), we obtain
(16)
(16)
For the solution going from to
, we integrate (Equation9
(9)
(9) ) from jT + r to
with k = j, which reaches
(17)
(17)
By (Equation12
(12)
(12) ) and k = j, (Equation17
(17)
(17) ) becomes
(18)
(18)
Similarly, to get
, we take the derivative with respect to u of both sides of (Equation18
(18)
(18) ), which gives
(19)
(19)
Substituting (Equation12
(12)
(12) ) with k = j and (Equation18
(18)
(18) ) into (Equation19
(19)
(19) ), we get
(20)
(20)
Case 2: . In this case, Equation (Equation5
(5)
(5) ) is
where , and
. We first observe that the relationship between
and
is the same as case 1, i.e. with
, (Equation20
(20)
(20) ) still holds in this case. Thus, for getting the expression for
, we only need to know the expression for
. To this end, we turn to analyse the next equation
where
. By the methods of separation of variables and partial fraction decomposition of the rational function, we obtain
(21)
(21)
where
Further computations from (Equation21
(21)
(21) ) offer
(22)
(22)
Integrating (Equation22
(22)
(22) ) from jT to jT + r with k = j + 1, and by (Equation10
(10)
(10) ), we reach
(23)
(23)
Set
Then, we have, by calculating the partial derivative of
with respect to u,
(24)
(24)
and (Equation23
(23)
(23) ) becomes
(25)
(25)
Differentiating on both sides of (Equation25
(25)
(25) ) with respect to u yields
(26)
(26)
Substituting (Equation24
(24)
(24) ) and (Equation25
(25)
(25) ) into (Equation26
(26)
(26) ) with k = j + 1, we obtain
The above preliminaries are sufficient to pave the way for presenting our main results.
3. Exact two periodic solutions
In this section, we give a sufficient condition for model (Equation3(3)
(3) ) to admit exactly two periodic solutions, with one stable and the other unstable, which is harboured in the following theorem.
Theorem
Assume that . Then model (Equation3
(3)
(3) ) has exactly two T-periodic solutions, with the smaller one unstable and the larger one asymptotically stable.
The theoretical proof of the Theorem will be given after the following three indispensable lemmas are introduced.
First, following the lines in [Citation32,Citation34,Citation41], when , we have
which ensures the existence of two T-periodic solutions to model (Equation3
(3)
(3) ). To sum up, we have
Lemma 3.1
Assume , then model (Equation3
(3)
(3) ) has two T-periodic solutions, with their corresponding initial values lie in
and
.
Next, we prove the uniqueness of the corresponding T-periodic solutions initiated from and
in the following two lemmas.
Lemma 3.2
Assume that . Then model (Equation3
(3)
(3) ) has a unique T-periodic solution
with
.
Proof.
The existence of such that
can be deduced from Lemma 3.1. Denote
. Then
is a T-periodic solution of model (Equation3
(3)
(3) ). Next, we prove the uniqueness of T-periodic solutions by contradiction.
Assume that there exists such that
see Figure (B) and (C) for illustration. Then, the growth rate of the Poincaré map h at
with i = 1, 2 satisfies one of the following two conclusions:
(27)
(27)
which correspond to Figure (B) and (C), respectively.
Figure 2. When , schematic illustrations for proving the uniqueness of T-periodic solutions initiated from
, where
. Panel (A) graphs that model (Equation3
(3)
(3) ) has a unique T-periodic solution, panels (B) and (C) describe that model (Equation3
(3)
(3) ) has exactly two T-periodic solutions, which correspond to cases (i) and (ii) in (Equation27
(27)
(27) ), respectively.
![Figure 2. When 0<c≤G∗, schematic illustrations for proving the uniqueness of T-periodic solutions initiated from (E1(j),E1(j+1)), where E1(k)=E1(k,c),k=j,j+1. Panel (A) graphs that model (Equation3(3) {dwdt=−aξww+(j+1)c[(w−A2)2+μ(j+1)aξ(c−G∗)],t∈[(i−1)T,(i−1)T+r),dwdt=−aξww+jc[(w−A2)2+μjaξ(c−j+1jG∗)],t∈[(i−1)T+r,iT),(3) ) has a unique T-periodic solution, panels (B) and (C) describe that model (Equation3(3) {dwdt=−aξww+(j+1)c[(w−A2)2+μ(j+1)aξ(c−G∗)],t∈[(i−1)T,(i−1)T+r),dwdt=−aξww+jc[(w−A2)2+μjaξ(c−j+1jG∗)],t∈[(i−1)T+r,iT),(3) ) has exactly two T-periodic solutions, which correspond to cases (i) and (ii) in (Equation27(27) (i)h′(u1)≥1,h′(u2)=1;(ii)h′(u1)=1,h′(u2)≥1,(27) ), respectively.](/cms/asset/47950c4f-dca7-48e5-bc3b-b9fe73c42a9e/tjbd_a_2023666_f0002_oc.jpg)
To obtain a contradiction, we need to take the derivative of . Set
Then we have, from (Equation16
(16)
(16) ) and (Equation20
(20)
(20) ),
Define
(28)
(28)
Then at points
, we have
. Since
we have
. From (Equation27
(27)
(27) ), cases (i) and (ii) imply
(29)
(29)
and
(30)
(30)
respectively.
However, from (Equation28(28)
(28) ), we have
which, along with
for
, yield
(31)
(31)
Since
always holds. Otherwise, we have
. Hence, from (Equation31
(31)
(31) ) and the facts
, we have
. A contradiction to (Equation29
(29)
(29) ) and (Equation30
(30)
(30) ). Obviously, (Equation31
(31)
(31) ) can also exclude the possibility of three or more T-periodic solutions to model (Equation3
(3)
(3) ). The proof is complete.
Lemma 3.3
Assume that . Then model (Equation3
(3)
(3) ) has a unique T-periodic solution
with
.
Proof.
From Lemma 3.1, there exists such that
Denote
. Then
, and
is a T-periodic solution of model (Equation3
(3)
(3) ). Hence, we only need to prove the uniqueness of T-periodic solutions to model (Equation3
(3)
(3) ) for
. Similarly, assume that model (Equation3
(3)
(3) ) has another T-periodic solution in this case, and we denote it by
. Then as illustrated in Figure (A) and (B), there are two cases:
(32)
(32)
and
Meanwhile, the facts
with i = 1, 2 and
for
imply
(33)
(33)
and
(34)
(34)
hold, which respectively correspond to cases (i) and (ii) in (Equation32
(32)
(32) ).
Figure 3. When , schematic illustrations for proving the uniqueness of T-periodic solutions initiated from
, where
. Panels (A) and (B) manifest that model (Equation3
(3)
(3) ) has exactly two T-periodic solutions, which correspond to cases (i) and (ii) in (Equation32
(32)
(32) ), respectively.
![Figure 3. When 0<c≤G∗, schematic illustrations for proving the uniqueness of T-periodic solutions initiated from (E2(j+1),E2(j)), where E2(k)=E2(k,c),k=j,j+1. Panels (A) and (B) manifest that model (Equation3(3) {dwdt=−aξww+(j+1)c[(w−A2)2+μ(j+1)aξ(c−G∗)],t∈[(i−1)T,(i−1)T+r),dwdt=−aξww+jc[(w−A2)2+μjaξ(c−j+1jG∗)],t∈[(i−1)T+r,iT),(3) ) has exactly two T-periodic solutions, which correspond to cases (i) and (ii) in (Equation32(32) (i)h′(v1)≤1,h′(v2)=1;(ii)h′(v1)=1,h′(v2)≤1,(32) ), respectively.](/cms/asset/19dcfc4a-b9e6-4476-ab8b-39b3cf144263/tjbd_a_2023666_f0003_oc.jpg)
However, taking the derivative of yields
and hence
for
, which leads to a contradiction to
in (Equation33
(33)
(33) ) and a contradiction to
in (Equation34
(34)
(34) ). Further,
is sufficient to exclude the possibility of three or more T-periodic solutions to model (Equation3
(3)
(3) ). We complete the proof.
With the above three lemmas, we are now ready to give the detailed proof of the theorem.
Proof
Proof of the Theorem
From Lemmas 3.1–3.3, we have already known that model (Equation3(3)
(3) ) has exactly two T-periodic solutions, namely,
and
, and
(35)
(35)
as shown in Figure . Let
, then from Lemma 2.2 and (Equation35
(35)
(35) ), we obtain that
is unstable. Hence, we only need to prove that
is asymptotically stable.
Figure 4. A schematic illustration for figuring out the proof of the asymptotic stability to the larger T-periodic solution , which is the solid red curve, where
.
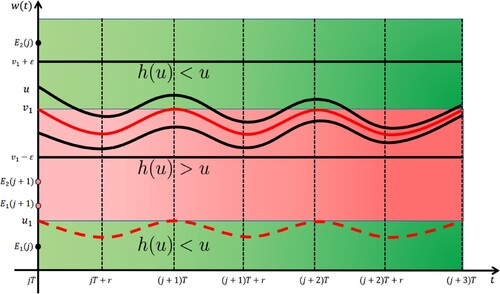
For convenience, we denote in the following.
We first prove that is stable. For any
, we only need to show that
implies
(36)
(36)
where
. To this end, we choose
, and prove (Equation36
(36)
(36) ) by contradiction. In fact, if not, then there exists
such that
Without loss of generality, we assume that
Since (Equation3
(3)
(3) ) is T-periodic, we may assume
.
Let i>j be a positive integer such that . Then there are three possible cases to consider: (I)
; (II)
; (III)
. We first show that (I) is impossible. In fact, if (I) holds, then we have
which shows that
However, from Lemmas 2.1, 3.1–3.3, we know that sequence
is strictly increasing, which leads to a contradiction. Thus, (I) is impossible.
We then prove that (II) is also impossible. If not, then we have
(37)
(37)
Furthermore, from (Equation12
(12)
(12) ) and (Equation13
(13)
(13) ), with k = j, we can derive that
which manifests that function
is strictly decreasing with respect to u. Thus, from (Equation18
(18)
(18) ) and (Equation37
(37)
(37) ), we have
that is, we have
, or, equivalently,
, which gives a similar contradiction to that in the proof of case (I). Thus, (II) is also impossible.
Finally, we prove (III) is impossible as well. We first prove that the case of is impossible. Integrating (Equation9
(9)
(9) ) from iT to
with k = j + 1, we obtain
(38)
(38)
similarly, integrating (Equation9
(9)
(9) ) from
to
with k = j + 1, we get
(39)
(39)
from (Equation12
(12)
(12) ) and (Equation13
(13)
(13) ), with k = j + 1, we have
which implies that function
is strictly increasing with respect to u. Furthermore, since
(40)
(40)
(Equation38
(38)
(38) ) and (Equation39
(39)
(39) ) tell us that
, therefore, follows from the increasing monotonicity of
with respect to u, we have
, a similar contradiction to that in the proof of case (I) occurs again. Thus, the case of
is impossible.
We then prove the case of is also impossible. Integrating (Equation9
(9)
(9) ) from
to
with k = j, we have
(41)
(41)
Similarly, integrating (Equation9
(9)
(9) ) from
to iT with k = j, we reach
(42)
(42)
Recall that function
is strictly decreasing with respect to u, thus, (Equation40
(40)
(40) ), (Equation41
(41)
(41) ) and (Equation42
(42)
(42) ) give
, hence, we obtain
. A similar contradiction as that in the proof of case (I) can be achieved. Thus, the case of
is also impossible. Hence, (Equation36
(36)
(36) ) is true, which implies that
is stable.
For the attractivity of , we need to prove
(43)
(43)
For the case when
, from (Equation35
(35)
(35) ), we have
, hence,
. In fact, from Lemma 2.1, we know that sequence
is strictly increasing, so
this shows that (Equation43
(43)
(43) ) is true for the case when
. For the case when
, from (Equation35
(35)
(35) ), we have
, the remaining proof is similar to that of
and is omitted. We complete the proof.
In the following, we will carry out a numerical example to illustrate our theoretical results.
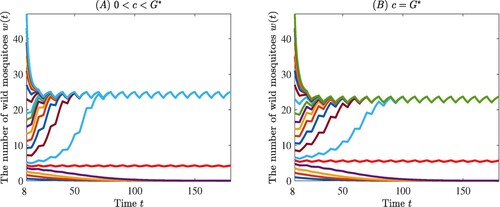
Example 1
Let
By taking
, we have
. Then we choose
or
, the condition of the Theorem is satisfied and
is asymptotically stable, along with an asymptotically stable T-periodic solution, which are respectively shown in panels (A) and (B) of Figure .
Figure 5. Panels (A) and (B) are used to support our theoretical results, where the two curves in red colour represent the unstable T-periodic solution .
4. Concluding remarks
Based on the modelling idea in [Citation31] that the number of sterile mosquitoes released is treated as a given nonnegative function rather than a variable satisfying an independent dynamical equation, the interactive dynamics of wild and sterile mosquitoes with or without time delay has been recently studied in [Citation18,Citation21,Citation32–34,Citation41]. A common assumption is that the sterile mosquitoes are impulsively and periodically released at discrete time points kT, . Such impulsive and periodic release strategy is consistent with the actual field release situation [Citation42].
Model (Equation1(1)
(1) ) is derived by combining the ideas in [Citation6] and [Citation31]. We introduce two release amount thresholds
and
, and the waiting period threshold
. In [Citation43], for the case of
, the authors obtained sufficient conditions for the global asymptotic stability of the origin and the existence of a globally asymptotically stable T-periodic solution, respectively. It is well known for obtaining sufficient conditions on the existence of exact two periodic solutions is mathematically challenging. In this paper, for the case of
, we show that model (Equation1
(1)
(1) ) has exactly two T-periodic solutions and analyse their stability, the bigger one is stable and the smaller one is unstable. The result, combining with that of [Citation43], indicates the better understanding of how to determine a better release strategy for biological control of wild mosquitoes from practical release perspective. Further investigations, for the case of
, are understudy, this will be a topic of our future study. We hope that our theoretical results can provide real guidance to help the practical workers make the better release strategy to gain a better effect for the wild mosquito population suppression.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- S. Ai, J. Li, J. Yu and B. Zheng, Stage-structured models for interactive wild and periodically and impulsively released sterile mosquitoes, Discrete Contin. Dyn. Syst. Ser. B, doi: 10.3934/dcdsb.2021172.
- L. Alphey, M. Benedict and R. Bellini, et al. Sterile-insect methods for control of mosquito-borne diseases: An analysis, Vector Borne Zoonotic Dis. 10 (2010), pp. 295–311.
- H. Barclay, Pest population stability under sterile releases, Res. Popul. Ecol. 24 (1982), pp. 405–416.
- H. Barclay, Mathematical models for the use of sterile insects, In: Sterile insect technique: Principles and practice in area-wide integrated pest management, V. Dyck, J. Hendrichs and A. Robinson, eds., Springer, Netherlands, 2005, pp. 147–174.
- L. Cai, S. Ai and G. Fan, Dynamics of delayed mosquitoes populations models with two different strategies of releasing sterile mosquitoes, Math. Biosci. Eng. 15 (2018), pp. 1181–1202.
- L. Cai, S. Ai and J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math. 74 (2014), pp. 1786–1809.
- CDC, Life cycle: The mosquito, Preprint 2019, Available from: https://www.cdc.gov/dengue/resources/factsheets/mosquitolifecyclefinal.pdf.
- K. Fister, M. McCarthy, S. Oppenheimer and C. Collins, Optimal control of insects through sterile insect release and habitat modification, Math. Biosci. 244 (2013), pp. 201–212.
- Z. Guo, H. Guo and Y. Chen, Traveling wavefronts of a delayed temporally discrete reaction-diffusion equation, J. Math. Anal. Appl. 496 (2021), pp. 124787.
- M. Hirsch, S. Smale and R. Devaney, Differential Equations, Dynamical Systems, and an Introduction to Chaos, 2nd ed., Academic Press, Orlando, 2003.
- A. Hoffmann, B. Montgomery and J. Popovici, et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission, Nature 476 (2011), pp. 454–457.
- L. Hu, M. Huang, M. Tang, J. Yu and B. Zheng, Wolbachia spread dynamics in stochastic environments, Theor. Popul. Biol. 106 (2015), pp. 32–44.
- L. Hu, M. Tang, Z. Wu, Z. Xi and J. Yu, The threshold infection level for wolbachia invasion in random environment, J. Differ. Equ. 266 (2019), pp. 4377–4393.
- M. Huang, J. Luo, L. Hu, B. Zheng and J. Yu, Assessing the efficiency of Wolbachia driven Aedes mosquito suppression by delay differential equations, J. Theoret. Biol. 440 (2018), pp. 1–11.
- M. Huang, M. Tang and J. Yu, Wolbachia infection dynamics by recation-diffusion equations, Sci. China Math 58 (2015), pp. 77–96.
- M. Huang, M. Tang, J. Yu and B. Zheng, A stage structured model of delay differential equations for Aedes mosquito population suppression, Discrete Contin. Dyn. Syst. 40 (2020), pp. 3467–3484.
- M. Huang, J. Yu, L. Hu and B. Zheng, Qualitative analysis for a Wolbachia infection model with diffusion, Sci. China Math. 59 (2016), pp. 1249–1266.
- Y. Hui, G. Lin, J. Yu and J. Li, A delayed differential equation model for mosquito population suppression with sterile mosquitoes, Discrete Contin. Dyn. Syst. Ser. B 25 (2020), pp. 4659–4676.
- I. Iturbe-Ormaetxe, T. Walker and S. O'Neill. Wolbachia and the biological control of mosquito-borne disease, EMBO Rep. 12 (2011), pp. 508–518.
- J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dyn. 11 (2017), pp. 316–333.
- G. Lin and Y. Hui, Stability analysis in a mosquito population suppression model, J. Biol. Dyn. 14 (2020), pp. 578–589.
- Y. Liu, Z. Guo, M. Smaily and L. Wang, A Wolbachia infection model with free boundary, J. Biol. Dyn. 14 (2020), pp. 515–542.
- Y. Liu, F. Jiao and L. Hu, Modeling mosquito population control by a coupled system, J. Math. Anal. Appl. 506 (2022), pp. 125671.
- F. Liu, C. Yao, P. Lin and C. Zhou, Studies on life table of the natural population of Aedes albopictus, Acta Sci. Nat. Univ. Sunyat. 31 (1992), pp. 84–93.
- Y. Shi and J. Yu, Wolbachia infection enhancing and decaying domains in mosquito population based on discrete models, J. Biol. Dyn. 14 (2020), pp. 679–695.
- Y. Shi and B. Zheng, Discrete dynamical models on Wolbachia infection frequency in mosquito populations with biased release ratios, J. Biol. Dyn., 2021, doi: 10.1080/17513758.2021.1977400.
- C. Stone, Transient population dynamics of mosquitoes during sterile male releases: Modelling mating behaviour and perturbations of life history parameters, PLoS ONE 8 (2013), pp. e76228. doi: 10.1371/journal.pone.0076228.
- E. Waltz, US reviews plan to infect mosquitoes with bacteria to stop disease, Nature 533 (2016), pp. 450–451.
- S. White, P. Rohani and S. Sait, Modelling pulsed releases for sterile insect techniques: Fitness costs of sterile and transgenic males and the effects on mosquito dynamics, J. Appl. Ecol. 47 (2010), pp. 1329–1339.
- World Health Organization, Dengue and severe dengue, Preprint 2021, Available from: http://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue.
- J. Yu, Modelling mosquito population suppression based on delay differential equations, SIAM J. Appl. Math. 78 (2018), pp. 3168–3187.
- J. Yu, Existence and stability of a unique and exact two periodic orbits for an interactive wild and sterile mosquito model, J. Differ. Equ. 269 (2020), pp. 10395–10415.
- J. Yu and J. Li, Dynamics of interactive wild and sterile mosquitoes with time delay, J. Biol. Dyn. 13 (2019), pp. 606–620.
- J. Yu and J. Li, Global asymptotic stability in an interactive wild and sterile mosquito model, J. Differ. Equ. 269 (2020), pp. 6193–6215.
- J. Yu and B. Zheng, Modeling Wolbachia infection in mosquito population via discrete dynamical models, J. Differ. Equ. Appl. 25 (2019), pp. 1549–1567.
- B. Zheng, J. Li and J. Yu, One discrete dynamical model on Wolbachia infection frequency in mosquito populations, Sci. China Math., (2021), doi: 10.1007/s11425-021-1891-7.
- B. Zheng, M. Tang and J. Yu, Modeling Wolbachia spread in mosquitoes through delay differential equation, SIAM J. Appl. Math. 74 (2014), pp. 743–770.
- B. Zheng, M. Tang, J. Yu and J. Qiu, Wolbachia spreading dynamics in mosquitoes with imperfect maternal transmission, J. Math. Biol. 76 (2018), pp. 235–263.
- B. Zheng and J. Yu, Characterization of Wolbachia enhancing domain in mosquitoes with imperfect maternal transmission, J. Biol. Dyn. 12 (2018), pp. 596–610.
- B. Zheng and J. Yu, Existence and uniqueness of periodic orbits in a discrete model on Wolbachia infection frequency, Adv. Nonlinear Anal. 11 (2021), pp. 212–224.
- B. Zheng, J. Yu and J. Li, Modeling and analysis of the implementation of the Wolbachia incompatible and sterile insect technique for mosquito population suppression, SIAM J. Appl. Math. 81 (2021), pp. 718–740.
- X. Zheng, D. Zhang and Y. Li, et al. Incompatible and sterile insect techniques combined eliminate mosquitoes, Nature 572 (2019), pp. 56–61.
- Z. Zhu, B. Zheng, Y. Shi, R. Yan and J. Yu, Stability and periodicity in a mosquito population suppression model composed of two sub-models, Accepted.