
ABSTRACT
Leptospirosis is a widespread zoonotic and neglected infectious disease of human and veterinary concern that is caused by pathogenic Leptospira species. After entrance in the host, pathogenic leptospires evade the host natural defense mechanisms in order to propagate and disseminate to multiple organs. Myeloperoxidase is an enzyme stored in neutrophils azurophilic granules, and is released upon neutrophil activation to produce mainly hypochlorous acid, a strong oxidant and potent antimicrobial agent. In the present investigation, we studied the modulation of myeloperoxidase activity by L. interrogans serovar Copenhageni. We show that leptospires and their culture supernatants are able to inhibit both peroxidase and chlorination activities of myeloperoxidase, without interfering with neutrophil degranulation. By leptospiral outer membrane protein extraction and fractionation, we identified the proteins LipL21 and LipL45 as myeloperoxidase inhibitors, constituting new Leptospira virulence factors. Accordingly, we propose a function for the protein LipL21, one of the most expressed leptospiral outer membrane proteins. Our results show a novel innate immune evasion mechanism by which leptospires interfere with the host response in order to cope with the host oxidative stress and efficiently achieve dissemination and colonization.
1. Introduction
Leptospirosis is one of the most prevalent zoonotic diseases worldwide, caused by the pathogenic spirochete Leptospira. The disease is frequently acquired through contact with contaminated water or soil, causing infection on humans mainly via skin or mucosa [Citation1,Citation2]. After entering the host, leptospires rapidly disseminate through the blood stream. This is followed by the bacterial clearance from blood and their migration into hosts' tissues, being the kidneys and liver the preferential sites for colonization. The infection may vary from non-symptomatic to different clinical conditions ranging from a mild flu-like symptomatology to a severe form known as Weil's disease. Severe leptospirosis is characterized by jaundice, hypotension, acute lung injury, bleeding complications and multi-organ failure, reaching up to 70% mortality rates [Citation1,Citation3–5].
The molecular mechanisms of leptospiral pathogenesis are poorly understood. After entrance in the host, pathogenic leptospires evade the host natural defense mechanisms, in order to propagate and disseminate to multiple organs, finally reaching immunologically privileged sites, such as the proximal renal tubules [Citation1]. Understanding host-pathogen interactions is an important approach to identify new virulence factors.
Polymorphonuclear neutrophils (PMNs) have a critical role in innate immunity, constituting one of the body's first line of defence against bacterial pathogens [Citation6]. Once activated by a chemotactic signal, PMNs adhere to the endothelium, where they start to infiltrate into the infected tissue and secrete antimicrobial and pro-inflammatory mediators or finally phagocytize invading bacteria [Citation7–10].
Myeloperoxidase (MPO) is a heme-containing and glycosylated enzyme that mainly catalyses the production of hypochlorous acid (HOCl) in the presence of hydrogen peroxide (H2O2) and chloride anions [Citation11–13]. MPO is packaged in the neutrophils azurophilic granules and is released into phagosomes during phagocytosis of exogenous structures or secreted extracellularly under stress conditions [Citation14,Citation15]. Hypochlorous acid (HOCl), the enzymes' major product, is a strong oxidant that reacts with proteins, lipids and DNA, being a potent antimicrobial agent, providing a front-line defence against pathogens [Citation15,Citation16]. MPO activity has also considerable tissue damaging potential when released extracellularly during excessive inflammation [Citation17].
As PMNs constitute the largest population of circulating phagocytes, they are expected to play an important role in leptospiral clearance. It was reported that PMNs are able to kill both non-pathogenic and pathogenic strains of Leptospira by oxygen dependent and independent mechanisms, although opsonization by specific antibodies is needed for phagocytosis of these microorganisms [Citation18–20]. Although non-pathogenic Leptospira strains were found to be more susceptible to killing by hydrogen peroxide (H2O2) and by released contents of PMNs primary granules in vitro, the same effect is observed towards pathogenic L. interrogans. The mechanisms of bacterial killing by H2O2 and neutrophil primary granules degranulation occurs via mechanisms independent of MPO, therefore not a consequence of halogenation reactions [Citation18].
In this work, we evaluated the modulation of MPO activity by L. interrogans serovar Copenhageni. We show that leptospires and their secreted products inhibit both chlorination and peroxidase MPO activities. We also identified the protein LipL21 as a potent MPO inhibitor, constituting a new leptospiral virulence factor. Our results show a new mechanism by which leptospires interfere with the host immune response in order to efficiently disseminate and colonize the host.
2. Material and methods
2.1. Bacteria and culture conditions
L. interrogans serovar Copenhageni L1-130 was kindly provided by Dr. Mathieu Picardeau (Institute Pasteur, France). Leptospires were cultured at 28°C in Elinghausen-McCullough-Johnson-Harris (EMJH) medium (BD, Difco) supplemented with 10% Leptospira enrichment EMJH medium (BD, Difco), 0.3 g/L peptone (BD, Difco) and 0.2 g/L meat extract (Sigma-Aldrich).
2.2. Leptospiral membrane proteins isolation
L. interrogans membrane proteins were isolated by the detergent Triton X-114 (TX-114) method previously described for Treponema pallidum, with modifications [Citation21]. Briefly, leptospires were suspended at a concentration of 2.5 × 109/mL in TX-114 extraction solution (1% TX-114, 10 mM Tris [pH 7.5], 150 mM NaCl, 2 mM EGTA) at 4°C. The suspension was placed on a rotating platform for 4 h at 4°C, then centrifuged at 17,000 x g at 4°C for 30 min. The resulting supernatant was mixed with CaCl2 to a final concentration of 10 mM and additional TX-114 up to 2% concentration. The samples were incubated at 37°C for 30 min in a water bath and then centrifuged at 25°C and 2,000 x g for 10 min to separate the phases. The upper aqueous was discarded and the lower TX-114 phase washed with 20% of its volume in 10 mM Tris (pH 7.5) to remove any aqueous material remaining at the detergent-aqueous phase interface. The TX-114 fraction was precipitated with 10 volumes of acetone for 16 h at −20°C, centrifuged at 12,600 x g at 4°C for 30 min, and the pellet was air dried prior to solubilisation in phosphate-buffered saline (PBS) pH 7.4 containing urea.
2.3. Protein fractionation by anion exchange chromatography
The solubilized TX-114 fraction diluted in PBS, applied onto a PD 10 column (GE Healthcare) and eluted with 20 mM Tris-HCl pH 8.0. The sample was loaded onto a HiTrap Q HP column (GE Healthcare) and eluted stepwise in fractions using increasing NaCl concentrations. The eluates were concentrated, desalted and dialyzed to PBS using Amicon Ultra-15 Centrifugal Filter Units (Merck Millipore). Fractions were analysed by SDS-PAGE.
2.4. Cloning, expression and purification of the recombinant proteins
The genes encoding for the leptospiral proteins LipL21, LipL45 and Lsa63 were amplified from L. interrogans serovar Copenhageni genome using the primers pairs listed at . The primers were designed in a way to remove the predicted signal peptides as predicted by the server SMART (http://smart.embl-heidelberg.de/). The amplified inserts encoding for LipL45 and Lsa63 were cloned into the pET101 vector (Invitrogen), while LipL21 into pAE [Citation22]. The positive clones were transformed into BL21-SI Star (DE) bacteria, and induced for 1 h (LipL21), 4 h (Lsa63) or overnight (LipL45) at 37°C by the addition of 0.5 mM IPTG. The bacteria were pelleted, lysed by sonication (50 mM Tris-HCl pH 7.5, 200 mM NaCl, 5% glycerol, 0.05% Triton X-100) and centrifuged (12.000 x g, 15 min, 4°C) for the separation of soluble proteins from inclusion bodies. The recombinant proteins were purified from the soluble fractions. Purification was performed by affinity chromatography using HisTrap HP (GE Healthcare) column in Äkta prime equipment (GE Healthcare) with gradient imidazole elution. Fractions containing the eluted recombinant proteins were pooled and dialyzed against PBS.
Table 1. Gene locus and sequence of the oligonucleotides employed for amplification of the target genes from L. interrogans serovar Copenhageni genomic DNA.
2.5. Circular dichroism (CD) spectroscopy
The proteins were dialysed against 10 mM sodium phosphate buffer and the CD spectroscopy of the far-UV spectrum was obtained in a Jasco J-810 spectropolarimeter (Japan Spectroscopic). The resulting spectra are presented as the averages of ten scans recorded from 190 to 260 nm. The residual molar ellipticity is expressed in degree cm2 dmol−1. Spectral data were analysed with the software CAPITO for estimation of the secondary structure content.
2.6. Peptide mass fingerprinting (PMF)
The purified recombinant protein LipL45 was separated by 12% SDS-PAGE, and the 45 and 31 kDa bands were excised from the gel. The in gel digestion of the proteins and preparation for PMF was performed as previously described [Citation23]. The samples were analyzed by MALDI-TOF mass spectrometry (MS) using 2.5 dihydroxybenzoic acid as the matrix on an MALDI-TOF Autoflex Speed Instrument (Bruker). The MS was performed on the reflectron and positive ion mode.
2.7. Neutrophils isolation
Human neutrophils were purified from citrated blood using Polymorphprep™ gradient (AXIS-SHIELD), according to the manufacturer's instructions. Informed consent was obtained from all blood donors.
2.8. Stimulation of neutrophils
Isolated neutrophils (8 × 105 cells/well in 100 µL PBS) were incubated for 3 h at 37°C in 5% CO2 atmosphere with 200 µL of stimuli: leptospires (1 × 108, 5 × 107, 2.5 × 107 or 1 × 107 bacteria/mL), bacterial mid-log phase culture supernatants (approximately 1 × 108 cells/mL) (at 100% or diluted to 50% or 20% with fresh EMJH culture medium), 25 nM Phorbol 12-myristate 13-acetate (PMA) (Sigma-Aldrich), PBS or EMJH. Notably, PMA is a very known stimulator of PMNs by protein kinase C activation [Citation24]. After incubation, the plates were centrifuged (1000 x g, 5 min) and the supernatants were stored for elastase and MPO activity assays.
Alternatively, to the generation of neutrophils exudates for posterior inhibition assays, neutrophils were stimulated with 25 nM PMA for 5 h and the supernatants were stored at −20°C until use.
2.9. Neutrophil-released elastase activity assay
Elastase activity was determined using the EnzChek Elastase Assay Kit (Invitrogen), according to instructions of the manufacturer. A volume of 25 µL of the supernatants originated from neutrophils stimulation was employed. Purified elastase (0.2 U/mL) was used as positive control of the assay. Fluorescence was measured at different time points.
2.10. Neutrophil-released myeloperoxidase activity assay
MPO chlorination and peroxidase activities were determined using the Enzchek MPO Activity Assay Kit (Invitrogen), according to instructions of the manufacturer. For detection of chlorination, the kit provides nonfluorescent 3´-(p-aminophenyl) fluorescein (APF), which is selectively cleaved by hypochlorite (–OCl) to yield fluorescein. Peroxidase activity is detected using nonfluorescent Amplex® UltraRed reagent, which is oxidized by the H2O2 to form a fluorescent product. Fluorescence relative to substrate degradation were determined by excitation at 485 nm and emission at 535 nm. Supernatants from neutrophils stimulated with leptospires or bacterial culture supernatants (50 µL) were employed.
Alternatively, exudates of neutrophils stimulated with 25 nM PMA were incubated (20 min at 37°C) with L. interrogans (1 × 108, 5 × 107, 2.5 × 107 or 1 × 107 bacteria/mL), bacterial culture supernatants (at 100%, 50% or 20% dilution), PBS or EMJH. After removal of bacteria by centrifugation, chlorination MPO activity was determined.
2.11. Effect of iron, membrane protein fractions and recombinant proteins on purified MPO activity
To test the effect of iron complementation on MPO inhibition by leptospires, 50 ng/mL purified MPO (50 µL) (R&D Systems) were incubated with increased concentrations of FeCl3 as a source of Fe3+ (0, 5, 10, 20, 40 and 100 µM) in the presence or absence of 50 µL 1 × 108 leptospires /mL for 20 min at 37°C. Leptospires were removed by centrifugation, and 50 µL of the supernatants were used for chlorination activity assay by Enzchek MPO Activity Assay Kit (Invitrogen).
To test the inhibitory effect of Leptospira membrane protein fractions after TX-114 fractionation, 50 ng/mL purified MPO (50 µL) were incubated with 10 µL of each protein fraction for 20 min at 37°C, and 50 µL of the supernatants were used for chlorination activity assay.
When recombinant proteins were employed, 50 ng/mL purified recombinant human MPO was incubated for 20 min at 37°C with the leptospiral recombinant proteins LipL21, LipL45 and Lsa63 at final concentrations of 50, 250, 500 or 1000 ng/mL. MPO incubated with purified E. coli K12 lipopolysaccharide (LPS) (Invivogen), MPO without additions or PBS alone were used as controls. Fifty microliters of the mixtures were used for chlorination activity assay.
2.12. MPO western blot
MPO (0.5 µg/µL in 10 µL) was incubated with 100 µL of L. interrogans suspensions (1 × 108/mL). At specified time points (5, 15, 30 and 60 min), 20 µL aliquots were removed and stored at −20°C for posterior analysis. Samples were separated by 10% SDS-PAGE, transferred to membranes and blocked for 2 h at room temperature with PBS + 0,05% Tween 20 (PBS-T) containing 2% BSA. Membranes were then incubated overnight at 4°C with antibodies against human MPO (Santa Cruz Biotechnology) at 1:200 dilution. After a washing step, membranes were incubated with secondary antibodies. Reactivity was detected by chemiluminescence.
2.13. Statistics
Statistical differences between groups were analyzed using GraphPad Prism 6.07 using one-way ANOVA followed by Dunnett's multiple comparison test. Differences between groups were considered significant if P < 0.05.
3. Results
3.1. L. interrogans bacteria and their secreted products inhibit MPO activity
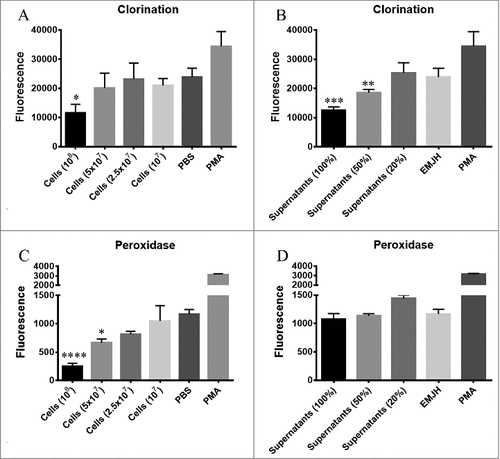
To first analyse the effect of leptospires in MPO degranulation and activity, the exudates of PMNs treated with different concentrations of L. interrogans bacteria or their culture supernatants were recorded using a chlorination activity assay, as this method is more specific for MPO than those based on peroxidase substrates. We observed that the exudates of PMNs incubated with bacteria or bacterial culture supernatants rendered less MPO activity than PMNs incubated only with PBS ( and ). This effect was dose-dependent, suggesting that the bacteria can produce/secrete a factor which can inhibit MPO chlorination activity. Cells stimulated with PMA, a very well-known strong activator of neutrophils by protein kinase C activation [Citation24], served as a positive control. As expected, PMA control elicited higher MPO activity, as it triggers neutrophils degranulation and no possible inhibitor was added. To determine if the inhibitory effect is directed specifically to the MPO chlorination activity or whether it interferes with its peroxidation cycle, the peroxidase activity of the PMNs exudates was also analysed. As seen in and , leptospires and their secreted products inhibited the peroxidase activity of PMNs-secreted MPO in a dose-dependent manner.
Figure 1. MPO activity in the supernatants of neutrophils stimulated with L. interrogans cells or their secreted products. Human purified PMNs were stimulated for 3 h at 37°C with different concentrations of L. interrogans serovar Copenhageni cells (A and C) or their culture supernatants (B and D). PMNs stimulated with PMA, PBS (when leptospires were assayed) or EMJH (when supernatants were assayed) alone were used as controls. The co-incubation supernatants were assayed for MPO chlorination (A and B) or peroxidase activity (C and D), expressed as fluorescence measurement of the substrate degradation (485 nm excitation and 535 nm emission). Fluorescence is indicated in arbitrary units. The bars represent the means ± standard deviation of three independent experiments performed in triplicates. Statistics is provided by comparison with the PBS control group: #P < 0.05, ##P < 0.01, ###P < 0.001 and ####P < 0.0001.
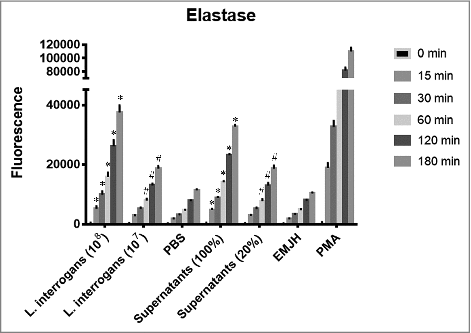
Apart from MPO, also elastase is stored in neutrophils azurophilic granules. Therefore, the two proteins are supposed to be concomitantly released upon degranulation [Citation10]. Corroborating the idea that the leptospires are efficient in stimulating PMNs and that the observed lower MPO activity is due to enzymatic inhibition and not due to reduced PMNs degranulation, we measured the elastase activity in the same cells exudates used for the MPO activity assays. We show that elastase activity increases in a time and dose-dependent manner when incubated with leptospires or their secreted products (). When PMNs were incubated with PBS, a reduced elastase activity was measured. We therefore conclude that leptospires are able to activate PMNs and trigger the release of the contents stored in the azurophilic granules.
Figure 2. Elastase activity in the supernatants of neutrophils stimulated with L. interrogans cells or their secreted products. Human purified PMNs were stimulated for 3 h at 37°C with different concentrations of L. interrogans serovar Copenhageni cells or their culture supernatants. PMNs stimulated with PMA, PBS (when leptospires were assayed) or EMJH (when supernatants were assayed) alone were used as controls. The co-incubation supernatants were assayed for elastase activity expressed as fluorescence measurement of the substrate degradation (485 nm excitation and 535 nm emission) at different time points. Fluorescence is indicated in arbitrary units. The bars represent the means ± standard deviation of three independent experiments performed in triplicates. Statistics is provided by comparison with the PBS control group at each time point: #P < 0.05 and #P < 0.0001.
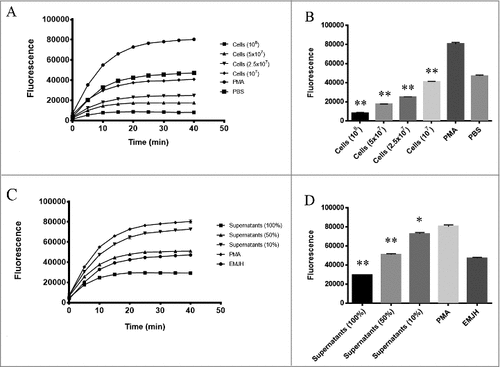
In another set of experiments, isolated human PMNs were first stimulated to degranulate upon incubation with PMA. The PMA-treated neutrophils exudates were then incubated with different concentrations of L. interrogans or leptospiral culture supernatants. Once again, we found that MPO chlorination activity is strongly reduced by the addition of L. interrogans or their secreted products (), further supporting or data that leptospires interfere with MPO activity rather than with MPO degranulation. Collectively, our results strongly imply that L. interrogans presents mechanisms to potently inhibit neutrophil-derived MPO chlorination and peroxidase activities.
Figure 3. L. interrogans cells and their secreted products inhibit the activity of neutrophil-secreted MPO. Human purified PMNs were stimulated for 5 h at 37°C with 25 nM PMA. PMNs exudates were then incubated for 20 min at 37°C with L. interrogans cells in different concentrations (A and B) or their culture supernatants (C and D) in different dilutions. The mixtures were assayed as MPO chlorination activity, expressed as fluorescence measurement of the substrate degradation (485 nm excitation and 535 nm emission). A and C: kinetics of MPO activity measured at different time points. B and D: MPO activity measured at the end-point time (40 min). Fluorescence is indicated in arbitrary units. The bars represent the means ± standard deviation of three independent experiments performed in triplicates. Statistics is provided by comparison with the PMA with no additions control group: #P < 0.001 and ##P < 0.0001.
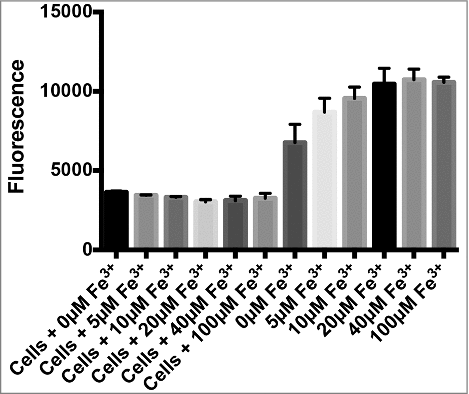
3.2. Leptospiral MPO inhibition is not related to iron sequestration
The bacterial siderophore enterobactin was described to act as MPO inhibitor by efficiently sequestration of the iron from the heme group bared by MPO [Citation25]. We therefore decided to analyse if Leptospira-mediated MPO inhibition could arise from similar mechanisms of iron chelation, even though leptospires are believed to lack siderophores [Citation26,Citation27]. Purified MPO was incubated with increasing concentrations of FeCl3 as a source of Fe3+ in the presence or absence of L. interrogans. The supplementation of the reaction with excess of Fe3+ did not prevent MPO inhibition by leptospires (), implying that the inhibitory effect might be due to a mechanism unrelated to iron sequestration.
Figure 4. Evaluation of MPO integrity and effect of iron supplementation on L. interrogans MPO inhibition. (A) Purified human MPO was incubated in the presence or absence of L. interrogans cells for different times (5, 15, 30 and 60). The supernatants were then subjected to SDS-PAGE and transferred to membranes. The integrity of MPO was evaluated by western blot analysis using specific antibodies. (B) Purified human MPO was incubated with increasing concentrations of FeCl3 as a source of Fe3+ (0, 5, 10, 20, 40 and 100 µM) in the presence or absence of 50 µL 1 × 108 leptospires /mL for 20 min at 37°C. The MPO chlorination activity was measured in the supernatants, expressed as fluorescence measurement of the substrate degradation (485 nm excitation and 535 nm emission). Fluorescence is indicated in arbitrary units. The bars represent the means ± standard deviation of three independent experiments performed in triplicates.
3.3. The leptospiral protein LipL21 strongly inhibits MPO
In the next series of experiments, we sought to identify the leptospiral proteins able to act as inhibitors of human MPO. We decided to focus only on the surface-exposed components. The separation of the supernatants to identify the active agent(s) was not performed because the Leptospira secretome is poorly characterized and there are no well-established protocols. We purified the L. interrogans serovar Copenhageni L1-130 membrane proteins by TX-114 solubilisation, employing a method modified from Cunningham et al [Citation21]. The extracted leptospiral membrane proteins were then fractionated by anion exchange chromatography, being eluted by increasing NaCl concentrations. After buffer exchange to PBS, each fraction was tested as the ability to inhibit the chlorination activity of purified recombinant human MPO. The fractions eluted with 300, 350, 400 and 450 mM NaCl were able to decrease MPO activity (). The positive fractions were analysed by SDS-PAGE and the predicted inhibitor candidates were identified by MS/MS mass spectrometry to be the proteins LipL21 and LipL45 ( and ). Other unidentified leptospiral proteins on the positive fractions could also act as MPO inhibitors.
Figure 5. Inhibition of MPO activity by L. interrogans membrane proteins fractions. L. interrogans serovar Copenhageni membrane proteins were extracted by TX-114 and then fractionated by anion exchange chromatography with elution in different NaCl concentrations (50–1000 mM). The membrane proteins fractions were dialysed against PBS and incubated for 20 min at 37°C with purified recombinant human MPO. (A) The mixtures were assayed as MPO chlorination activity, expressed as fluorescence measurement of the substrate degradation (485 nm excitation and 535 nm emission). Fluorescence is indicated in arbitrary units. The bars represent the means ± standard deviation of three independent experiments performed in duplicates. Statistics is provided by comparison with 100% MPO activity: #P < 0.05 and ##P < 0.01. (B) The fractions were separated by 8% and 15% SDS-PAGE, and the protein bands predicted to function and MPO inhibitors by comparative and exclusion analysis, were identified by mass spectrometry (C).
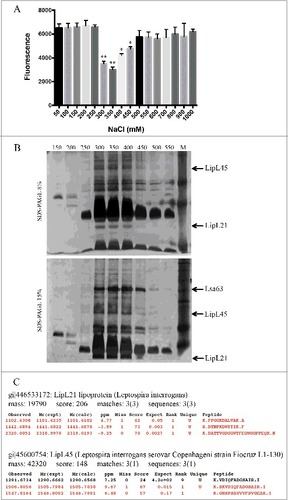
The genes coding for LipL21 and LipL45 were cloned into pAE or pET101 vectors, respectively. All recombinant proteins were produced in E. coli in their soluble form. Expression was induced by addition of IPTG and purification performed by metal chelating chromatography (). It was previously reported [Citation28] that degradation of LipL45 leads to the generation of a 31 kDa fragment (, lane 3). The gene for LipL45 was cloned into pET101 vector, which adds a C-terminal his-tag to the protein. As the 31 kDa band is recognized in Western blot analysis by an anti-his tag antibody, the fragment has to be originated from the C-terminal portion of LipL45 (). Moreover LipL45 processing was analysed by the comparison of the PMF spectra of the full LipL45 protein and the 31 kDa fragment. The existence of coincident peaks shown in reaffirms that the 31 kDa fragment is derived from the full LipL45. Additional analysis of these peaks by MS/MS fragmentation also corroborated that the 31 kDa fragment is a LipL45 degradation product (not shown). The leptospiral protein Lsa63 was included as a negative control. The structural integrity of the three purified recombinant proteins was assessed by CD spectroscopy. Spectra data analysis revealed that the proteins are structured (), with calculated alpha helix and beta sheet contents similar to the predicted by the amino acid sequences. The features of the leptospires recombinant proteins are summarised in the . The three proteins were experimentally demonstrated to localize at the leptospiral outer membrane [Citation28–31] while their predicted localization may vary. Both LipL21, LipL45 and Lsa63 have homologs in the non-pathogenic and saprophytic Leptospira biflexa serovar Patoc, although their sequence identity is low (47, 55 and 39%, respectively).
Figure 6. SDS-PAGE analysis of the purified recombinant proteins. The leptospiral proteins LipL21, LipL45 and Lsa63 were expressed in E. coli BL21 (DE3) and purified by Ni2+ chelating affinity chromatography. The purified proteins were dialysed against PBS and analysed by 12% SDS-PAGE stained with coomassie blue. The molecular mass protein markers are indicated on the left.
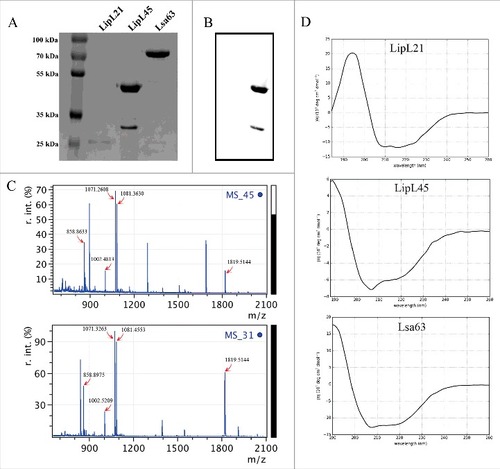
Table 2. Features of the leptospiral recombinant proteins analysed as MPO inhibitors.
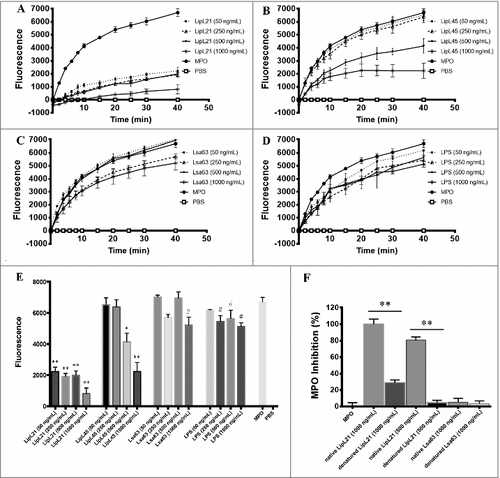
When increasing concentrations of the purified recombinant proteins LipL21, LipL45 and Lsa63 were incubated with a fixed amount of MPO (50 ng/mL), a strong inhibitory effect of the MPO chlorination activity was observed even at low LipL21 concentration (50 ng/mL) ( and ), while LipL45 had a significant inhibitory effect only at a higher concentration (500 ng/mL) ( and ). The controls, Lsa63 and purified LPS, had no effect on MPO activity (, and ). Corroborating our results, when heat-denatured, LipL21 inhibitory activity is significantly reduced, suggesting that the proper protein conformation is needed for the MPO inhibition (). Our results show that both LipL21 and LipL45 are able to inhibit MPO, being LipL21 the most potent protein in this task. Bioinformatics analysis of LipL21 and LipL45 did not reveal any conserved domain or predicted function that could explain the mechanisms by which they act as MPO inhibitors ().
Figure 7. Inhibition of MPO activity by L. interrogans purified recombinant proteins. Fifty ng/mL purified recombinant human MPO were incubated for 20 min at 37°C with the leptospiral recombinant proteins LipL21, LipL45 and Lsa63 at final concentrations of 50, 250, 500 or 1000 ng/mL. MPO incubated with purified bacterial LPS, MPO without additions or PBS alone were used as controls. The mixtures were then assayed as MPO chlorination activity, expressed as fluorescence measurement of the substrate degradation (485 nm excitation and 535 nm emission). A-D: kinetics of MPO activity measured at different time points. E: MPO activity measured at the end-point time (40 min). F: LipL21 and Lsa63 were heat-denatured before MPO inhibition assays. LipL21 inhibition at 1000 ng/mL was considered 100%. Fluorescence is indicated in arbitrary units. The bars represent the means ± standard deviation of three independent experiments performed in triplicates. Statistics is provided by comparison with the MPO positive control: #P < 0.0001, ##P < 0.001 and #P < 0.05.
4. Discussion
The induction of a host innate immune response by invading pathogens generates a defensive oxidative burst. Accounting for the destructive capacity of neutrophils, H2O2 and a variety of more potent secondary oxidants are produced [Citation14,Citation32,Citation33]. Most of the H2O2 generated by neutrophils is consumed by MPO. Apart from the classical heme peroxidase activity, MPO is unique in its chlorination activity, readily oxidizing chloride ions to (HOCl) [Citation11]. HOCl is the most bactericidal oxidant produced by neutrophils, readily reacting with thiols, thioethers and amines [Citation34].
Leptospires must have evolved mechanisms to effectively escape from or neutralize the host oxidative burst defense mechanism. During the host oxidative burst, reactive oxygen species (ROS) are generated from free radicals of molecular oxygen. The generated oxygen radicals can then be used to produce other ROS, including H2O2, hydroxyl radicals, and HOCl, all of which mediating bacterial killing. When performing genomic analyses, leptospires were found to lack superoxide dismutase [Citation35], what would increase the leptospires' susceptibility to the oxidative burst generated during the infection [Citation36]. However, this vulnerability may be overcome by other mechanisms to cope with oxidative stress, such as expression of metalloporphyrins, peroxidase and catalase [Citation37]. The catalase KatE of L. interrogans has been shown to be important for H2O2 detoxification and bacterial survival [Citation38].
In this study, we describe a novel mechanism by which L. interrogans evades the innate immune response of the host and copes with the oxidative stress. We consistently demonstrated that leptospires can inhibit the host potent neutrophil-derived antibacterial enzyme MPO in a dose-dependent fashion. Such inhibition of MPO has effect on both peroxidase and chlorination activities of the enzyme. A study by Murgia and colleagues (2002) previously described that MPO lacks microbicidal activity towards leptospires. However, no mechanism was proposed showing how the bacteria can counteract MPO-induced production of ROS. We now demonstrate that this is due to inhibition of MPO's enzymatic activity. Our results further show that the two leptospiral proteins, LipL21 and LipL45, respectively, are responsible for this ability. While LipL21 is a potent MPO inhibitor, with an activity even at very low concentrations, LipL45 functions at higher concentrations and has only a moderate effect. Hence our study supports the notion of leptospiral outer membrane protein LipL21 as the most prominent factor protecting the bacteria against oxidative stress through blocking the production of the potent oxidizing agent HOCl.
Bioinformatics analysis of LipL21 and LipL45 did not reveal any conserved domain, homologs or predicted function that could explain the mechanisms by which they act as MPO inhibitors. Experiments trying to understand the mechanisms of MPO inhibition are currently under development. LipL21 and LipL45 were reported to be localized at the leptospiral surface, thus potentially mediating host-pathogen direct interactions [Citation28,Citation29]. LipL21 is one of the most abundant outer membrane proteins of Leptospira, exhibiting more than 8,000 copies per cell [Citation39]. LipL45 is also expressed in high levels by leptospires (approximately 3,000 copies per cell) [Citation39]. LipL45 was shown to be initially expressed as a 45 kD lipoprotein which is further processed to become a 31 kD peripheral membrane protein by removal of the carboxyterminal portion [Citation28]. This processing was also observed in our assays for recombinant protein production. Both LipL21 and LipL45 were found to be expressed by leptospires during infection [Citation28,Citation29,Citation40] hence suggesting the participation and importance of these proteins for L. interrogans pathogenesis and virulence.
To our knowledge, the only bacterial proteins able to inhibit MPO reported to date are siderophores. The siderophore enterobactin was described to act as MPO inhibitor by efficient sequestration of the iron from the heme group bared by MPO [Citation25]. As leptospires have been reported to lack siderophores [Citation26,Citation27] our data present a novel mechanism by which these bacteria interact with MPO activity, independent of iron chelation. As one of the most abundant outer membrane proteins of L. interrogans, LipL21 action on the protection against the host oxidative burst by inhibition of MPO activity might be a crucial step for leptospiral virulence and infectious process.
Although a common and non-specific finding in acute bacterial infections, PMN count is usually increased during the leptospiremic phase of leptospirosis and is correlated to disease severity [Citation41,Citation42]. The production of ROS is elevated in leptospirosis patients and correlates to tissue injury levels, although the contribution of PMNs was not evaluated [Citation43]. Also, the study by Scharrig et al [Citation44]. has shown that neutrophil extracellular traps (NETs) are critical for the prevention of early bacteria dissemination in a murine model of the disease. Therefore, the participation of neutrophils in the initial phase of leptospirosis seems to be an important defense mechanism, mainly driven by secretion of antimicrobial compounds, ROS production and NETs formation, rather than phagocytosis. Indeed, a recent study reported that macrophages but not neutrophils are the main phagocytes killing L. interrogans in vitro, which are the major infiltrating cells in infected tissues [Citation45]. The evasion from neutrophilic innate armory seems to be essential for the progression of leptospirosis, and the present manuscript presents a novel mechanism by which leptospires can achieve this by inhibiting MPO.
In conclusion, we have demonstrated that the lipoprotein LipL21, expressed at L. interrogans outer membrane, by inhibiting the MPO-catalysed reaction with H2O2, define a novel defence mechanism of leptospires against the host innate immune system. Inhibition of MPO might be an important aspect of the leptospiral immune escape strategy and survival. Our results represent an important contribution to the understanding of leptospiral pathogenesis and host-pathogen interactions.
Conflicts of interest
None declared.
Author contributions
Conceived the experiments: MV, HH. Designed the experiments: MV, AT, AC, GQ. Performed the experiments: MV, AT, AC, GQ. Analyzed the data: MV, AC, GQ, HH. Contributed with reagents/materials/analysis tools: DT, AN, HH. Wrote the paper: MV, HH. All authors revised the manuscript.
Acknowledgments
We thank Dr. Mathieu Picardeau for kindly providing the Leptospira strains and Pia Andersson for excellent technical assistance.
Additional information
Funding
References
- Faine S, Adler B, Bolin C, et al. Leptospira and Leptospirosis. Melbourne, Australia: MediSci; 1999. p. 259.
- Plank R, Dean D. Overview of the epidemiology, microbiology, and pathogenesis of Leptospira spp. in humans. Microbes Infect. 2000;2:1265–76. doi:10.1016/S1286-4579(00)01280-6.
- Levett PN. Leptospirosis. Clin Microbiol Rev. 2001;14:296–326. doi:10.1128/CMR.14.2.296-326.2001.
- Vinetz JM. Leptospirosis. Current opinion in infectious diseases. 2001;14:527–38. doi:10.1097/00001432-200110000-00005.
- Toyokawa T, Ohnishi M, Koizumi N. Diagnosis of acute leptospirosis. Expert Rev Anti Infect Ther. 2011;9:111–21. doi:10.1586/eri.10.151.
- Burg ND, Pillinger MH. The neutrophil: function and regulation in innate and humoral immunity. Clin Immunol. 2001;99:7–17. doi:10.1006/clim.2001.5007.
- Crockett-Torabi E. Selectins and mechanisms of signal transduction. Journal of leukocyte biology. 1998;63:1–14.
- Ley K. Integration of inflammatory signals by rolling neutrophils. Immunol Rev. 2002;186:8–18. doi:10.1034/j.1600-065X.2002.18602.x.
- Wahl SM, Feldman GM, McCarthy JB. Regulation of leukocyte adhesion and signaling in inflammation and disease. Journal of leukocyte biology. 1996;59:789–96.
- Sheshachalam A, Srivastava N, Mitchell T, Lacy P, Eitzen G. Granule protein processing and regulated secretion in neutrophils. Front Immunol. 2014;5:448. doi:10.3389/fimmu.2014.00448.
- Hampton MB, Kettle AJ, Winterbourn CC. Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood. 1998;92:3007–17.
- Ravnsborg T, Houen G, Hojrup P. The glycosylation of myeloperoxidase. Biochim Biophys Acta. 2010;1804:2046–53. doi:10.1016/j.bbapap.2010.07.001.
- Van Antwerpen P, Slomianny MC, Boudjeltia KZ, et al. Glycosylation pattern of mature dimeric leukocyte and recombinant monomeric myeloperoxidase: glycosylation is required for optimal enzymatic activity. J Biol Chem. 2010;285:16351–9. doi:10.1074/jbc.M109.089748.
- Segal AW. How neutrophils kill microbes. Annu Rev Immunol. 2005;23:197–223. doi:10.1146/annurev.immunol.23.021704.115653.
- Klebanoff SJ. Myeloperoxidase: friend and foe. J Leukoc Biol. 2005;77:598–625. doi:10.1189/jlb.1204697.
- Arnhold J, Flemmig J. Human myeloperoxidase in innate and acquired immunity. Arch Biochem Biophys. 2010;500:92–106. doi:10.1016/j.abb.2010.04.008.
- Nicholls SJ, Hazen SL. Myeloperoxidase, modified lipoproteins, and atherogenesis. J Lipid Res. 2009;50 Suppl:S346–51. doi:10.1194/jlr.R800086-JLR200.
- Murgia R, Garcia R, Cinco M. Leptospires are killed in vitro by both oxygen-dependent and -independent reactions. Infect Immun. 2002;70:7172–5. doi:10.1128/IAI.70.12.7172-7175.2002.
- Wang B, Sullivan J, Sullivan GW, Mandell GL. Interaction of leptospires with human polymorphonuclear neutrophils. Infect Immun. 1984;44:459–64.
- Wang B, Sullivan JA, Sullivan GW, Mandell GL. Role of specific antibody in interaction of leptospires with human monocytes and monocyte-derived macrophages. Infect Immun. 1984;46:809–13.
- Cunningham TM, Walker EM, Miller JN, et al. Selective release of the Treponema pallidum outer membrane and associated polypeptides with Triton X-114. J Bacteriol. 1988;170:5789–96. doi:10.1128/jb.170.12.5789-5796.1988.
- Ramos CR, Abreu PA, Nascimento AL, et al. A high-copy T7 Escherichia coli expression vector for the production of recombinant proteins with a minimal N-terminal His-tagged fusion peptide. Braz J Med Biol Res. 2004;37:1103–9. doi:10.1590/S0100-879X2004000800001.
- Vieira ML, Pimenta DC, de Morais ZM, et al. Proteome analysis of Leptospira interrogans virulent strain. Open Microbiol J. 2009;3:69–74. doi:10.2174/1874285800903010069.
- Nauseef WM, Volpp BD, McCormick S, et al. Assembly of the neutrophil respiratory burst oxidase. Protein kinase C promotes cytoskeletal and membrane association of cytosolic oxidase components. J Biol Chem. 1991;266:5911–7.
- Singh V, Yeoh BS, Xiao X, et al. Interplay between enterobactin, myeloperoxidase and lipocalin 2 regulates E. coli survival in the inflamed gut. Nat Commun. 2015;6:7113. doi:10.1038/ncomms8113.
- Louvel H, Bommezzadri S, Zidane N, et al. Comparative and functional genomic analyses of iron transport and regulation in Leptospira spp. J Bacteriol. 2006;188:7893–904. doi:10.1128/JB.00711-06.
- Sritharan M, Asuthkar S, Sridhar V. Understanding iron acquisition by pathogenic leptospires: a review. Indian J Med Microbiol. 2006;24:311–6. doi:10.4103/0255-0857.29406.
- Matsunaga J, Young TA, Barnett JK, et al. Novel 45-kilodalton leptospiral protein that is processed to a 31-kilodalton growth-phase-regulated peripheral membrane protein. Infect Immun. 2002;70:323–34. doi:10.1128/IAI.70.1.323-334.2002.
- Cullen PA, Haake DA, Bulach DM, et al. LipL21 is a novel surface-exposed lipoprotein of pathogenic Leptospira species. Infect Immun. 2003;71:2414–21. doi:10.1128/IAI.71.5.2414-2421.2003.
- Cullen PA, Xu X, Matsunaga J, et al. Surfaceome of Leptospira spp. Infect Immun. 2005;73:4853–63. doi:10.1128/IAI.73.8.4853-4863.2005.
- Vieira ML, de Morais ZM, Goncales AP, et al. Lsa63, a newly identified surface protein of Leptospira interrogans binds laminin and collagen IV. J Infect. 2010;60:52–64. doi:10.1016/j.jinf.2009.10.047.
- Hyslop PA, Hinshaw DB, Scraufstatter IU, et al. Hydrogen peroxide as a potent bacteriostatic antibiotic: implications for host defense. Free Radic Biol Med. 1995;19:31–7. doi:10.1016/0891-5849(95)00005-I.
- Klebanoff SJ. Myeloperoxidase: Contribution to the microbicidal activity of intact leukocytes. Science. 1970;169:1095–7. doi:10.1126/science.169.3950.1095.
- Klebanoff SJ. Myeloperoxidase-halide-hydrogen peroxide antibacterial system. J Bacteriol. 1968;95:2131–8.
- Nascimento AL, Verjovski-Almeida S, Van Sluys MA, et al. Genome features of Leptospira interrogans serovar Copenhageni. Braz J Med Biol Res. 2004;37:459–77. doi:10.1590/S0100-879X2004000400003.
- Stewart PE, Carroll JA, Dorward DW, et al. Characterization of the Bat proteins in the oxidative stress response of Leptospira biflexa. BMC Microbiol. 2012;12:290. doi:10.1186/1471-2180-12-290.
- Picardeau M. Virulence of the zoonotic agent of leptospirosis: still terra incognita? Nat Rev Microbiol. 2017;15:297–307. doi:10.1038/nrmicro.2017.5.
- Eshghi A, Lourdault K, Murray GL, et al. Leptospira interrogans catalase is required for resistance to H2O2 and for virulence. Infect Immun. 2012;80:3892–9. doi:10.1128/IAI.00466-12.
- Malmstrom J, Beck M, Schmidt A, et al. Proteome-wide cellular protein concentrations of the human pathogen Leptospira interrogans. Nature. 2009;460:762–5. doi:10.1038/nature08184.
- Nally JE, Whitelegge JP, Bassilian S, et al. Characterization of the outer membrane proteome of Leptospira interrogans expressed during acute lethal infection. Infect Immun. 2007;75:766–73. doi:10.1128/IAI.00741-06.
- Craig SB, Collet TA, Wynwood SJ, et al. Neutrophil counts in leptospirosis patients infected with different serovars. Trop Biomed. 2013;30:579–83.
- Craig SB, Graham GC, Burns MA, et al. Haematological and clinical-chemistry markers in patients presenting with leptospirosis: A comparison of the findings from uncomplicated cases with those seen in the severe disease. Ann Trop Med Parasitol. 2009;103:333–41. doi:10.1179/136485909X435058.
- Araujo AM, Reis EA, Athanazio DA, et al. Oxidative stress markers correlate with renal dysfunction and thrombocytopenia in severe leptospirosis. Am J Trop Med Hyg. 2014;90:719–23. doi:10.4269/ajtmh.13-0667.
- Scharrig E, Carestia A, Ferrer MF, et al. Neutrophil Extracellular Traps are Involved in the Innate Immune Response to Infection with Leptospira. PLoS Negl Trop Dis. 2015;9:e0003927. doi:10.1371/journal.pntd.0003927.
- Chen X, Li SJ, Ojcius DM, et al. Mononuclear-macrophages but not neutrophils act as major infiltrating anti-leptospiral phagocytes during leptospirosis. PLoS One. 2017;12:e0181014. doi:10.1371/journal.pone.0181014.