

ABSTRACT
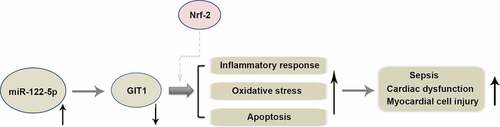
Myocardial injury resulting from sepsis is the leading cause of death worldwide. Micro RNA miR-122-5p is involved in various physiological and pathological processes and is highly expressed in the heart of septic rats. However, its function in sepsis-caused myocardial injury remains elusive. Herein, a rat model of septic myocardial injury was established by intraperitoneal injection of lipopolysaccharide (LPS), and cardiomyocyte H9c2 was exposed to LPS to induce sepsis-related inflammatory injury in vitro. Inhibition of miR-122-5p suppressed LPS-triggered myocardial injury evidenced by decreased heart weight index (HWI), reduced inflammatory cell infiltration and cell rupture, and reduced cardiac marker enzymes cTnI and LDH. MiR-122-5p inhibition inhibited ROS production and enhanced the activities of antioxidant enzymes CAT, SOD and GSH-px in LPS-treated rats and H9c2 cells. MiR-122-5p inhibition reduced the production of pro-inflammatory cytokines TNF-α, IL-6 and IL-1β, and inhibited cell apoptosis along with decreased cleaved-caspase 3 induced by LPS. Moreover, increased GIT1 expression was found following miR-122-5p inhibition. We further verified GIT1 as a target of miR-122-5p, and silencing GIT1 partially reversed the benefits of miR-122-5p loss in LPS-injured H9c2 cells. The HO-1 and NQO-1 expression and Nrf-2 activation were enhanced by miR-122-5p inhibition, which was reversed by GIT1 depletion, indicating the involvement of Nrf-2/HO-1 signaling in regulating miR-122-5p/GIT1-mediated cardioprotection. Taken together, our data suggest that inhibition of miR-122-5p may mitigate sepsis-triggered myocardial injury through inhibiting inflammation, oxidative stress and apoptosis via targeting GIT1, which provides a possible therapeutic target for sepsis.
GRAPHICAL ABSTRACT
KEYWORDS:
Highlights
Inhibition of miR-122-5p suppressed LPS-triggered myocardial injury.
MiR-122-5p inhibition reduced inflammatory response, oxidative stress and apoptosis in heart.
GIT1 was verified as a target of miR-122-5p.
Nrf-2/HO-1 is involved in regulating miR-122-5p/GIT1-mediated cardioprotection.
Acknowledgements
This study was supported by grants from the National Natural Science Foundation of China (No.81971810), and the 345 Talent Project of Shengjing Hospital of China Medical University.
Disclosure statement
The author(s) claim no competing interests in this study.
Author contributions
All authors equally contribute to the study conception and design. Material preparation, data collection and analysis were performed by WS, TZ, NY, TZ, RW. The draft of the manuscript was written by WS, and all authors commented on the draft paper. All authors read and approved the final manuscript.
Ethical approval
All animal studies in this study were in accordance with the protocols of ethical committees of Shengjing Hospital of China Medical University.
Data availability statement
The datasets generated during and/or analyzed during this study are available from the corresponding author on reasonable request.
Supplementary material
Supplemental data for this article can be accessed here.