

ABSTRACT
The present study aimed to assess the influences of corn straw application on the soil microbial organisms, soil enzyme activities and the grain yield. Four treatments were evaluated: (i) The straw was ploughed into soil using a fence hydraulic turning plow with ploughing depth of 30-40 cm(PD). (ii) The self-developed straw deep returning machine was used to bury 30-40 cm in the sub-surface layer of soil (SD). (iii) The straw was mulched and no tillage sowing(M). (iv)Without straw application(CK). Soil samples of different deep(0-20 cm, 20-40 cm soil layer) were taken during the corn growth stage to determinesoil biological characteristics.Our results suggested that soil microorganisms were not increased by straw mulching. Straw deep ploughing and returning (PD treatment) could effectively improve the phospholipid fatty acids(PLFAs) of bacteria, actinomycetes, and fungi, the activities of urease,invertase,dehydrogenase and polyphenoloxidase, even the grain yield. In 20-40 cm subsoil layer, the effects were more obvious than those of topsoil. The mean yield of PD treatment was higher than SD,M and CK. The results showed that the PLFA signatures and soil enzyme were both sensitive to the changes of soil environment condition by the application of straw. In the actual field production, we should adopt the appropriate way of straw returning to the field to achieve not only the improvement of soil quality, but also the increase of grain yield.
ABSTRACT
Article highlights
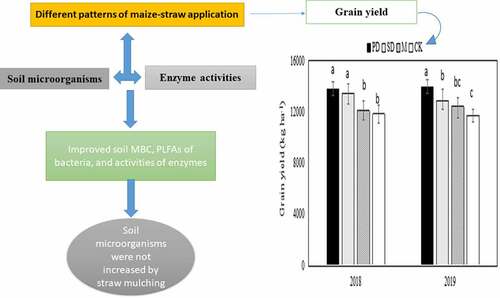
Soil microorganisms were not increased by straw mulching
PD treatment effectively improve soil MBC, PLFAs and enzymes activity.
The effects of subsoil (20-40 cm) were more obvious than those of topsoil.
The mean yield of PD treatment was higher than SD, M and CK.
Acknowledgements
The present study was supported by the National Key R & D Program “Establishment of key technical models for improving yield and efficiency of spring maize” (2017YFD0300607).