

Abstract
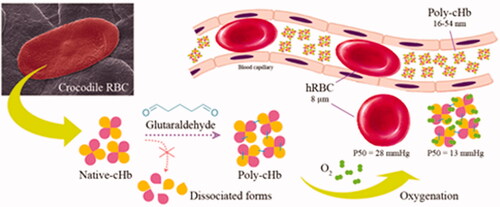
Hemoglobin-based oxygen carriers (HBOCs) are modified stroma-free hemoglobin molecules used in developing a blood substitute for therapeutic usage. In order to prevent hemoglobin dissociation, glutaraldehyde (GTA) was used to generate high-molecular weight heterogeneous crocodile hemoglobin (Poly-cHb). This work, Poly-cHb was created using various GTA concentrations, ranging from 0.025-0.150% (v/v). Physicochemical properties were investigated that were comparable GTA polymerized human hemoglobin (Poly-hHb). This study has revealed that GTA polymerization increases the molecular size of Native-cHbs from 14.10 nm over a range from 16.31 to 54.27 nm. Moreover, this polymerization alters the secondary structure and heme environment by decreasing the helicity ratio from 1.00 to 0.95 at the highest condition and exhibits hypochromic shift of the Soret band to be 0.88 times lower than the native. However, all Poly-cHbs still possessed higher oxygen affinity than that of Poly-hHbs with average P50 values of 13 and 21 mmHg, respectively. Although, polymerization affected the overall Poly-cHb structure slightly, but compensated by decreasing the denaturation level to lower than 10%. Thus, it is interesting to note that Poly-cHb may advantageously provide effective oxygen carriage and ability for pasteurization, which may benefit the search for new alternative hemoglobin sources for HBOC development.
GRAPHICAL ABSTRACT
Acknowledgements
I appreciate to especially thank Sriracha Moda Co., Ltd. (Chon Buri, Thailand) for supplying the crocodile blood sample and Department of Biochemistry, Faculty of Science, Khon Kaen University for providing the scientific instruments.
Disclosure statement
No potential conflict of interest was reported by the authors.