

Abstract
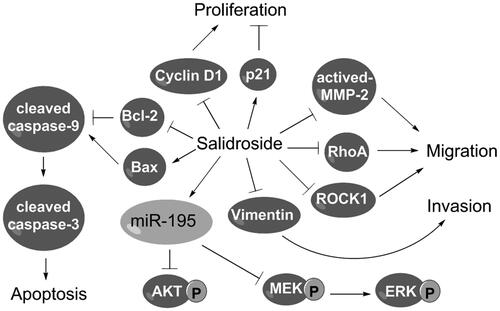
Salidroside, a glycoside of tyrosol, is isolated from Rhodiola rosea and shows anti-cancer functions in several cancers. However, the potentials of salidroside in the migration and invasion of lung cancer cells and its underlying mechanisms remain unknown. We aimed to investigate the functions and mechanisms of salidroside in non-small cell lung cancer (NSCLC). Human NSCLC cell line A549 was treated with different doses of salidroside. Cell viability, colony formation, apoptosis, migration and invasion were detected by CCK-8, crystal violet-staining assay, flow cytometry and transwell assay, respectively. qRT-PCR and western blot analysis were performed to assess the regulatory effects of salidroside on miR-195 expression and the activation of AKT and the MEK/ERK signal pathway. We found that, salidroside remarkably reduced cell viability, colony formation and Cyclin D1 expression, but increased p21 expression and apoptosis in A549 cells. Additionally, salidroside inhibited the migration and invasion of A549 cells by regulating expressions of migration- and invasion-related proteins. Finally, salidroside inhibited phosphorylation of AKT, MEK and ERK by upregulating miR-195 expression in A549 cells. In conclusion, salidroside inhibited the survival, migration and invasion of NSCLC cells. Salidroside blocked AKT and the MEK/ERK signal pathway by upregulating miR-195 expression in A549 cells.
GRAPHICAL ABSTRACT
Keywords:
Disclosure statement
No potential conflict of interest was reported by the authors.