
ABSTRACT
Despite impressive progress, malaria continues to impose a substantial burden of mortality and morbidity, particularly in sub-Saharan Africa, and new tools will be needed to achieve elimination. Gene drive is a natural process by which some genes are inherited at a greater-than-Mendelian rate and can spread through a population even if they cause harm to the organisms carrying them. Many different synthetic gene drive systems have been proposed to suppress the number of mosquitoes and/or reduce vector competence. As with any control measure, due attention should be paid to the possible evolution of resistance. No gene drive construct has yet been reported that is ‘field-ready’ for release, and when such constructs are developed, they should be assessed on a case-by-case basis. Gene drive approaches to vector control promise to have a number of key features that motivate their continued development, and scrutiny, by all concerned.
KEYWORDS:
Introduction to malaria
Burden. In 2015 3.2 billion people were at risk of malaria, with 95 countries or territories still with on-going transmission. There were about 214 million malaria cases, and 438,000 deaths, of which 88% and 90% respectively occurred in Africa, including 292,000 deaths in Africa of children under 5 years of age (WHO Citation2015: Table 2.1, 2.3). Pregnant women are also particularly vulnerable, and the economic burden of malaria is substantial.
Biology. Malaria is caused by Plasmodium parasites. There are five Plasmodium species that attack humans, of which four exist in Africa, the most important being Plasmodium falciparum, and the others being Plasmodium vivax, Plasmodium ovale, and Plasmodium malariae. The parasite is transmitted by the bite of an infected mosquito. The only way for a mosquito to acquire the parasite is to bite an infected person, and the mosquito then has to live about 10 days while the parasite develops within it and is ready to be transmitted to another person. Only a relatively small number of mosquito species can transmit malaria, and only female mosquitoes bite people and contribute to transmission. Different continents have different mosquito vector species; in Africa, the most important vectors are members of the Anopheles gambiae species complex, plus Anopheles funestus. In some locations other species are also important. By comparison, over 800 species of mosquito have been described from Africa (http://www.mosquitocatalog.org). Finally, at least in Africa, malaria is largely a rural disease (e.g. Tatem et al. Citation2013).
Current interventions. Key interventions against malaria include insecticide-treated nets (ITNs); indoor residual spraying (IRS); larval control; chemopreventive therapies for pregnant women and children; and treatment using artemisinin-based combination therapies (ACTs; WHO Citation2015). These interventions have saved many millions of lives, but malaria still persists even in places where coverage of the different interventions exceeds 80%. Subpopulations of mosquitoes may be escaping current tools due to atypical biting behaviour. And there is increasing resistance, both of the parasites to the drugs and of the mosquitoes to the insecticides, so it is possible that the recent advances could be reversed. Furthermore, these interventions are expensive for developing countries. Expert opinion suggests that without additional tools, malaria elimination will be out of reach in some locations no matter how much effort we deploy (Alonso et al. Citation2011). According to the WHO Global Technical Strategy for Malaria, 2016-2030 (WHO Citation2016), even under the most optimistic scenario of increased investment in existing interventions (with the resources available increasing to about $8.7 billion per year, about 3.5 times more than is currently available), death rates and case incidence can be reduced by 90%, and malaria eliminated from 35 countries, but it will still exist in 58 countries (requiring on-going expenditures; ).
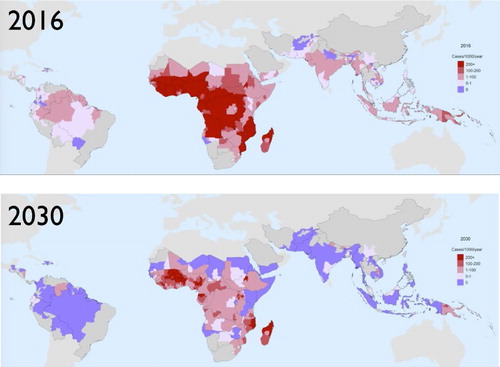
Figure 1. Comparison of the current incidence of P. falciparum malaria with that projected in 2030 under the most optimistic scenario considered by the WHO Global Technical Strategy for Malaria 2016–2030, scaling up existing interventions with some near-term innovation in longer lasting nets and better chemoprevention therapies, assuming no loss-of-effect due to drug or insecticide resistance. From Griffin et al. (Citation2016).
Genetic approaches to malaria control
One novel approach to malaria control that has been much discussed is to complement chemical-based vector control with genetic approaches (e.g. Adelman Citation2016). Many different genetic approaches can be imagined. For example, one could:
Interfere with survival or reproduction, thereby reducing the numbers of mosquitoes.
Bias the sex ratio towards males.
Change females’ host preference, so they feed on non-humans.
Specifically arrange for infected (or infectious) female mosquitoes to die.
Modify the females so that they could no longer support parasite development, either by knocking out a mosquito gene needed for parasite transmission (e.g. a receptor recognised by the parasite as it moves through the mosquito), or by introducing (‘knocking in’) a gene that interferes with parasite development.
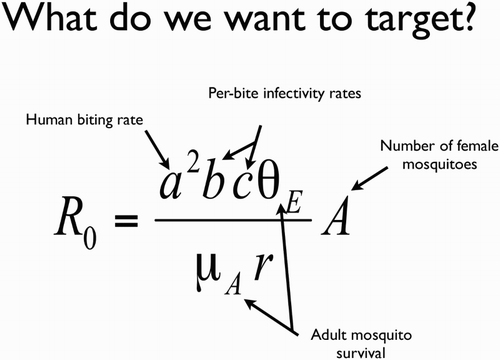
The basic goal of any of these approaches is to reduce the vectorial capacity of the mosquito (or R0 of the disease) as much as possible, by targeting one or more of the underlying parameters ().
Figure 2. The Ross–McDonald equation for the basic reproductive number for malaria. R0 is the expected number of new infections deriving from a single infection in an otherwise infection-free population; a is the rate at which an adult female bites a human; b is the probability that an infectious mosquito biting a human will transmit the parasite; c is the probability that an infected human will infect a mosquito that bites it; μA is the daily mortality rate for adult female mosquitoes; θE is the probability a female mosquito survives the period between acquiring an infection and becoming infectious; and A is the number of adult female mosquitoes per person. Potential genetic approaches to vector control target one or more of these parameters. r is the rate of recovery of humans, which is unlikely to be affected by vector control. For further details on the equation see Smith and McKenzie (Citation2004).
Whichever one of these approaches might be pursued, there are the twin challenges of modifying individual mosquitoes to manifest the change, and then having the modification spread through natural populations of the malaria mosquitoes. Malaria is a predominantly rural disease across vast regions of Africa, and it would be formidably difficult to spread the modification by inundative releases – i.e. by swamping the natural population with the desired modification. It also seems unlikely that the modification will be beneficial to the mosquito (and it is not clear it would be desirable if it was), and so one cannot rely on spread by natural selection. The alternative is to use the process of gene drive.
Introduction to gene drive
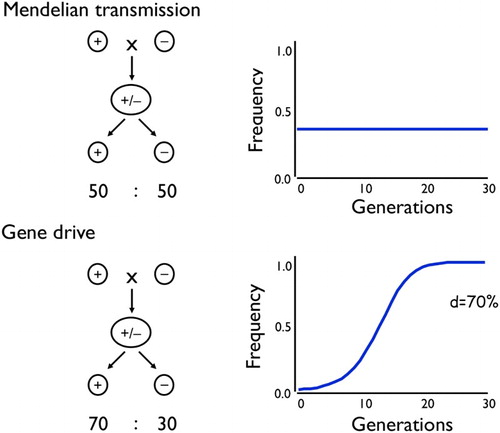
Most genes show Mendelian transmission: diploids have two copies of each gene, one from each parent, and they pass on one copy or the other to their offspring in an equal 50:50 ratio. In this case the process of gene transmission is ‘fair’, and does not in itself change the frequencies of alternative genes in a population – the two copies are transmitted by an individual at the same rate that they were inherited by the individual (). But some genes are transmitted to more than 50% of the (surviving) progeny. ‘Gene drive’ refers to this process of preferential or biased inheritance from one generation to the next. Now the outputs no longer match the inputs and there is a change in allele frequencies simply due to the process of gene transmission from one generation to the next. All else being equal, such genes will increase in frequency in a population (). Indeed, if the drive is strong enough, a gene can spread even if it causes some harm to the individuals carrying it. And the spread can be rapid: in a closed random mating population, a gene getting transmitted to 100% of progeny and having no fitness effect can increase in frequency from 1% to 99% in just nine generations. Given that mosquitoes have a generation about once a month, this implies very rapid spread – fast enough to be attractive for public health interventions. It is these properties that make gene drive such a promising tool for controlling malaria transmission.
Figure 3. Mendelian transmission of genes does not in itself lead to changes in allele frequency over time, whereas gene drive can. d is the proportion of a heterozygote’s progeny that inherit the driving gene (Mendelian transmission has d = 50%).
Gene drive is a natural process that has independently evolved many times in many species, with a wide diversity of underlying mechanisms. Genetic elements showing drive include transposable elements (found in the vast majority of eukaryotic genomes), gamete killers (well-studied examples include the t-haplotype in mice and Segregation Distorter in Drosophila), driving sex chromosomes (known from many dipteran species), maternal-effect killers (such as MEDEA in Tribolium), B chromosomes (found in many plants, insects, and other organisms), homing endonuclease genes (found in many microbes), and a variety of others (Burt and Trivers Citation2006). For some of these we know in quite some detail the molecular basis for the preferential inheritance; for others the molecular details are sketchy or unknown.
Synthetic gene drive systems
A wide diversity of synthetic gene drive systems have been proposed for vector control, and no doubt many more will be proposed in the future. Here we focus on those for which there has been promising laboratory work demonstrating technical feasibility.
Homing for gene knock-outs
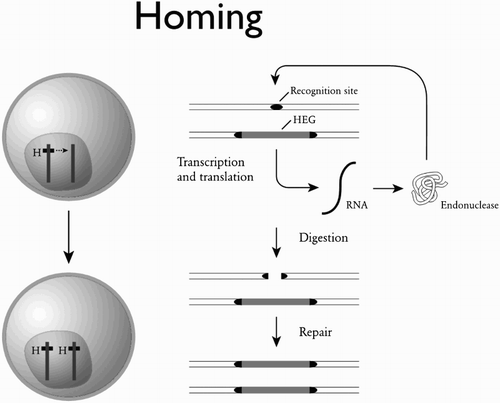
First discovered in yeast (Dujon Citation1989), the homing reaction is a particularly simple mechanism of achieving drive based on the activity of enzymes (‘endonucleases’) that cut specific sequences of DNA (). Many microbes have homing endonuclease genes, that spread in populations not because they are useful to the microbe, but because of the gene drive produced by the homing reaction. In animals a synthetic version of the homing reaction was first demonstrated in Drosophila melanogaster (Chan et al. Citation2011, Citation2013) and in A. gambiae (Windbichler et al. Citation2011) using a homing endonuclease (and its recognition site) from yeast. Homing against introduced sequences has also been shown in Drosophila using zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs; Simoni et al. Citation2014). More recently, high rates of homing have been demonstrated using an RNA-guided Cas9 nuclease against a body colour gene in Drosophila (Gantz and Bier Citation2015), an eye colour gene in A. stephensi (an Indian vector of malaria; Gantz et al. Citation2015), and three endogenous female fertility genes in A. gambiae (Hammond et al. Citation2016).
Figure 4. The homing reaction. Diploid cells that are initially heterozygous for the presence of a gene encoding a DNA-cutting enzyme (an ‘endonuclease’) can be converted to homozygotes when the gene is in the middle of its own recognition sequence. The presence of the gene protects the chromosome it is on from being cut, and only chromosomes not containing the gene are cut. The cell’s repair machinery can then use the intact chromosome as a template for repair, resulting in the gene being copied across to the chromosome where previously it was absent. HEG: homing endonuclease gene. After Burt and Trivers (Citation2006).
Homing can be used for vector control in a number of different ways. Conceptually, the simplest is to use it to produce a population-wide gene knock-out (). The effect of this on the mosquito population and on malaria transmission will then depend upon the gene being targeted.
Figure 5. A homing construct for population-wide gene knock-outs. A gene encoding a sequence-specific nuclease that recognises a sequence in a target gene is inserted in the middle of its own recognition sequence, which protects the chromosome it is on from being cut, but also knocks-out the function of the target gene. The nuclease must be expressed in the germline in order to show drive. After Burt (Citation2003).
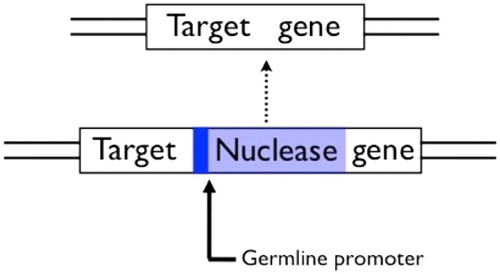
One possibility is to suppress mosquito populations by targeting one or more genes needed for survival or reproduction. Modelling shows that if the knock-out phenotype is recessive (i.e. survival or reproduction is close to normal if only one copy of the gene is disrupted, and substantially impaired only when both copies are disrupted), and if the homing reaction is confined to the germline (i.e. the tissues giving rise to sperm and eggs), then it is even possible for a homing endonuclease that causes lethality or sterility to increase in frequency in a population, potentially suppressing the population as it does so (Burt Citation2003; Eckhoff et al. Citation2017; Godfray, North, and Burt Citation2017).
Another possibility is to disrupt malaria transmission by targeting one or more genes needed for the Plasmodium parasite to invade into, develop within, or exit out of the mosquito vector. A recent review lists 38 genes that when knocked-down (e.g. by RNAi) show some reduction in oocyst number or sporozoite count (Sreenivasamurthy et al. Citation2013), and some of these genes may be suitable for this approach.
In principle, other effects are possible by targeting other sorts of genes (e.g. possibly host choice, by targeting olfactory genes). In order for the homing reaction to lead to preferential inheritance of the enzyme construct, it is essential that the enzyme be expressed in the germline. Thus far, the promoters used for this in Anopheles have been from the B2-tubulin gene (Catteruccia, Benton, and Crisanti Citation2005) and from the vasa gene (Papathanos et al. Citation2009).
Homing for knock-ins
The homing reaction can also be used to spread (‘knock-in’) a novel ‘cargo’ gene through a population. The cargo could be an effector gene that disrupts parasite transmission through the mosquito. A recent review lists 28 effector genes that interfere to some extent with malaria parasite transmission (Wang and Jacobs-Lorena Citation2013), including anti-microbial peptides, single-chain antibodies, immune system activators, and peptides that bind to mosquito proteins (putative parasite receptors) in the midgut or salivary glands.
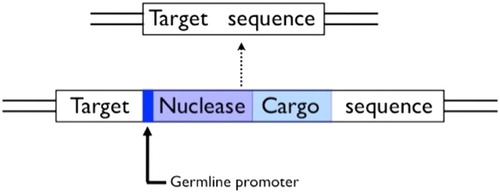
Conceptually, the simplest way to use the homing reaction to spread a cargo gene through a population is to include the cargo as part of the homing construct (Burt Citation2003; ). Many of the homing experiments cited above demonstrate how cargo genes can be carried along with a homing construct.
Figure 6. A homing construct for population-wide gene knock-ins. Note the cargo gene will itself need control sequences to ensure expression in the appropriate tissues.
One issue that arises with this approach is the fate of spontaneous mutations that occur in the construct itself, particularly loss-of-function mutations. In the simpler one-gene constructs that can be used for population knock-outs, mutations that arise in the nuclease gene which abolish enzyme function will fail to home, and therefore will be selected out and disappear. Similarly, in homing constructs carrying a cargo, mutations that abolish nuclease function will be selected out. However, mutations that abolish the function of the cargo gene(s) are not expected to affect the homing rate, and so may accumulate over time, particularly if the cargo genes impose some cost to the mosquito and the non-functional forms have higher fitness. One could end up with cargo-less constructs spreading through the population. For this reason it will be important with this strategy to choose effectors that do not impose much of a fitness cost, and to minimise the rate at which loss-of-function mutations arise, through choice of construct, location in genome, and/or promoter used (Beaghton et al. Citation2017). This issue can also in part be ameliorated using the homing reaction to target a gene involved in survival or reproduction, and then linking the effector(s) to a resistant gene (Burt Citation2003), a trans-acting suppressor of the nuclease, or a sequence that mutates the nuclease gene (e.g. Wu, Luo, and Gao Citation2016).
Driving Y chromosome for male bias
Another potential use for sequence-specific endonucleases in developing synthetic gene drive systems is to use them to create a driving sex chromosome. Because malaria is transmitted only by female mosquitoes, a population-wide distortion of the sex ratio towards males would have a direct impact in reducing malaria transmission. In addition, because the number of mosquitoes in one generation is largely a function of the number and productivity of females in the previous generation, a male-biased sex ratio may also lead to a reduction in the total number of mosquitoes, giving an additional reduction in malaria transmission (Deredec, Burt, and Godfray Citation2008, Citation2011; North, Burt, and Godfray Citation2013; Beaghton, Beaghton, and Burt Citation2016).
Maleness in Anopheles mosquitoes is thought to be determined by a dominant gene on the Y chromosome (Krzywinska et al. Citation2016). Males have X and Y sex chromosomes, and females have two Xs. Normally, the X and Y chromosomes of a male are transmitted 50:50 to the next generation, but if there is a gene causing the Y chromosome to be preferentially transmitted, then there will be a preponderance of male offspring. If the gene responsible is on the Y chromosome, then it too will be preferentially transmitted, and, all else being equal, it will spread through the population, replacing the normal Y. As it does so, the population as a whole will become male-biased. This is a driving Y chromosome.
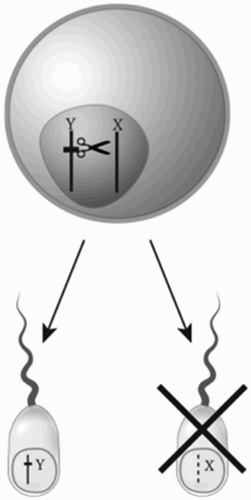
In Aedes mosquitoes there is a naturally occurring driving Y chromosome (or, more accurately, a driving male-determining region of a chromosome) that, in some crosses, leads to more than 90% male progeny. The underlying molecular mechanism has not yet been reported, but cytogenetically, drive is associated with breakage of the X chromosome at male meiosis (Newton, Wood, and Southern Citation1976). This led to the hypothesis that if a nuclease that specifically recognises and breaks the X chromosome can be expressed during meiosis, then that might lead to preferential transmission of the Y chromosome and a male bias (). Proof-of-principle demonstration of this route to drive in A. gambiae was reported by Galizi et al. (Citation2014), who showed that expression of an engineered variant of the PpoI nuclease from a slime mold, expressed during spermatogenesis using the B2-tubulin promoter sequences, led in some lines to males producing 95% male offspring. The PpoI enzyme recognises a sequence in the ribosomal RNA-encoding repeat, which, fortuitously, is found only on the X chromosome in A. gambiae.
Figure 7. A gene on the Y chromosome that encodes a nuclease that recognises and cleaves a repeated sequence found only on the X chromosome may lead to failure of the X-bearing sperm to develop properly and preferential transmission of the modified Y chromosome – a driving Y.
Other gene drive systems for spreading effector genes
Other sorts of gene drive systems have been proposed for spreading an effector through pest populations, including chromosomal rearrangements that display under-dominance, and various combinations of toxins and antidotes that mimic under-dominance systems, MEDEA systems, or variants thereof (Marshall and Akbari Citation2016).
In a maternal effect dominant embryonic arrest (MEDEA) system the progeny of heterozygous females die unless they themselves inherit the MEDEA element. A synthetic MEDEA element has been created in Drosophila by combining a microRNA-based repressor of myd88, a protein normally supplied by the mother into the embryo, with a zygotically expressed myd88 gene that is not affected by the microRNA and supplies the missing protein (Chen et al. Citation2007). Cage experiments showed the element could spread rapidly through populations. Two other MEDEA systems, using different components, have also been developed in Drosophila (Akbari et al. Citation2014) and Akbari et al. (Citation2013) developed a maternal-effect lethal under-dominance system in Drosophila in which two maternally expressed toxins, located on separate chromosomes, are each linked to a zygotic antidote able to rescue the lethality of the other toxin. Threshold-dependent invasion into population cages was observed.
These sorts of drivers are ‘weaker’ than those based on homing or driving sex chromosomes, in the sense that population models can be constructed in which, if introduced at very low frequencies, they may not spread, and if they spread at one location, they may not spread to a neighbouring population.
Resistance and reversal
A recurrent theme in the history of malaria control is the evolution of resistance, of either the Plasmodium parasite to a drug or the mosquito vector to an insecticide. It is therefore important to consider the prospects for resistance evolving against a driving construct, and take steps to minimise the likelihood of that happening (Burt Citation2003). For homing constructs aiming to suppress the numbers of mosquitoes by targeting a gene important for survival or reproduction, probably the most obvious way for resistance to arise is for mutations to occur at the target site that render the site less able to be cleaved by the enzyme. Such mutations may pre-exist in the population, before release, or may be created by the action of the nuclease, such as via a non-homologous end-joining (NHEJ) repair pathway (Champer et al. Citation2017; Hammond et al. Citation2017). Deredec, Burt, and Godfray (Citation2008, Citation2011) have modelled this situation, and if the mutations are functional for the mosquito (i.e. restore survival or reproduction), then they can spread to fixation, and the homing construct will be lost. It is therefore important that the sequences targeted be important for function. For CRISPR-based homing systems it is also possible to have multiple gRNAs, targeting multiple sites in the same target gene, which should help retard the evolution of resistance (Esvelt et al. Citation2014; Marshall et al. Citation2017). For homing constructs carrying an anti-Plasmodium effector gene, one might consider targeting a neutral part of the genome, but then one cannot rely on natural selection to remove the products of NHEJ repair, and these may end up spreading to fixation, leading to loss of the effector (Beaghton et al. Citation2017). One must also consider the possibility of the Plasmodium parasite evolving to be resistant to the effector(s), as they have to many drugs.
Closely related to the issue of resistance is the question of whether the effects of a gene drive construct might be reversed after release. As noted above, for homing constructs that target genes important for survival or reproduction, a sequence that is resistant to cleavage and functional for the mosquito (whether it arises de novo or is intentionally released) can spread by natural selection, allowing the population to recover (Burt Citation2003). Alternatively, one might release a trans-acting suppressor of the nuclease, or trans-acting mutator. For CRISPR-based systems, the latter could be as simple as a gRNA that targets the Cas9 nuclease (Wu, Luo, and Gao Citation2016). These and other potential strategies deserve careful modelling (Vella et al. Citation2017).
Moving to deployment
Work in the laboratory has provided a number of proof-of-principle demonstrations of key aspects of gene drive interventions for malaria control, but as yet there has been no report of a construct that has the potential to be ‘field-ready’. Even when such a construct is developed, there will still be a number of questions that those responsible for making decisions on deployment will want to address.
Example questions to at least consider before making a decision on release of a driving construct for Anopheles mosquito population suppression include:
Is the phenotype (e.g. male bias, female sterility) maintained:
in diverse genetic backgrounds?
in realistic, variable environments?
in other members of the A. gambiae complex?
Is there a genetic basis for escapees?
What is the potential timescale over which the target mosquito population(s) might evolve resistance, and what would be done if it does evolve?
What are the effects and dynamics of the construct in cage studies?
What are the effects and dynamics in a self-limiting (non-driving) open field release?
What release strategy might be needed to ensure spread and prevent re-invasion? What are the logistical and cost implications?
What are the likely community or ecosystem effects (e.g. on competitors, predators)?
More generally, what are the human health and environmental risks, and can they be managed or mitigated to an acceptable level?
This list is not intended to be exhaustive (see also WHO Citation2010; David et al. Citation2013; EFSA Citation2013; NASEM Citation2016). It is worth noting that current chemical vector control interventions (insecticide-treated bednets, IRS, and larval control) also suppress populations (e.g. Bayoh et al. Citation2010; Russell et al. Citation2011), providing precedents for both the epidemiological efficacy and ecological consequences of this approach.
Vector competence is a more variable or labile phenotype than lethality, sterility, or sex ratio, and more extensive studies may be required to ensure field efficacy. Example questions to at least consider before making a decision on release of a driving construct with an effector include:
Is the phenotype (reduced vector competence) maintained:
in diverse genetic backgrounds?
in realistic, variable environments?
in other members of the A. gambiae complex?
against a lab clone of P. falciparum (e.g. standard membrane feeding assay)?
against a diverse array of field genotypes (e.g. direct membrane feeding assay & direct feeding assay)
against other African species of Plasmodium?
If not, will there be competitive release (i.e. will suppression of one species lead to increase in another)?
Is there a genetic basis for escapees?
What is the potential timescale over which the target Plasmodium species might evolve resistance, and what would be done if it does evolve? Could selection for resistance lead to increased virulence in humans?
Is there an effect on transmission of other pathogens transmitted by A. gambiae (e.g. filariasis and O’nyong’nyong virus)?
What is the rate of dissociation between driver and effector in variable environments, what is the effect of the effector on mosquito fitness (e.g. from cage studies), and what do these imply for the temporal and spatial spread of intact elements?
What are the effects and dynamics in a self-limiting (non-driving) open field release?
What release strategy might be needed to ensure spread?
More generally, what are the human health and environmental risks, and can they be managed or mitigated to an acceptable level?
Again, this list is not intended to be exhaustive. For any gene drive approach, there will be questions of acceptability, the logistics of production, delivery, release and monitoring, long-term sustainability, the existence of a framework for responsible deployment, cost–benefit analyses, and issues around trans-border spread. For the latter, conventional biological control programmes offer some sort of precedent (Box 1). For the specific case of malaria control in Africa, capacity building in the end user disease endemic countries will be critical. Training of scientists from these countries and their involvement in research and deployment initiatives from the beginning will be essential for co-ownership. Training needs to be followed by the transfer of technologies and the installation of appropriate infrastructures (e.g. laboratories, mosquito production facilities, and field stations). Gene drive constructs pose issues that are new for regulators anywhere in the world, and African regulators should be involved from the outset in grappling with them. Equally important is strengthening capacities in communications and stakeholder engagement, which will play a central role in the success of deployment.
Although novel in many ways, gene drive approaches to mosquito and malaria control do have similarities with some well-established methodologies (Burt Citation2014).
Sterile insect technique (SIT)
SIT programmes involve the release of large numbers of sterile males into a target population; mating of these males with native females leads to a reduction in the females’ reproductive output, resulting in the reduction or elimination of the target population. SIT programmes have been highly successful against a number of agricultural insect pests, including the New World screwworm fly in North America; the Mediterranean fruit fly and other tephritids in many countries around the world; pink bollworm in the USA; and codling moth in Canada (Alphey et al. Citation2010). SIT has also been used to eliminate a tsetse fly vector of trypanosomiasis from Zanzibar.
Biological control
Gene drive approaches to vector control have clear parallels with classical biological control programmes. These involve releasing relatively small numbers of a natural enemy (predator, parasitoid, pathogen, etc.) to attack an invasive pest species. The natural enemy propagates itself over a period of generations, increasing in abundance over time; it is thus self-spreading and self-sustaining, and effects on the target population can be long-lived. Implementation is relatively inexpensive and the benefit/cost ratios of some programmes have been more than 100 (Zeddies et al. Citation2001). Classical biological control has been used to suppress over 200 species of invasive insects and 40 species of weeds in many countries around the world (van Driesche et al. Citation2008).
Key features
These various genetic approaches to vector control have the potential to provide a number of desirable features, including the following:
they act to reduce disease transmission, and thus can contribute to the goals of disease elimination and eradication;
they are widely applicable, able to act in diverse settings, whether hypo- or holoendemic, urban or rural, against mosquitoes that feed indoors or outdoors, during daytime or night-time, and can control mosquito populations that are otherwise difficult to access;
they provide area-wide control, and therefore protection without obvious biases relating to a person’s age, wealth, or education;
they should be compatible with and complementary to other disease control measures, both current (e.g. chemical-based vector control) and under development (e.g. vaccines); and
they can be relatively easy to deliver and deploy, with little or no change required in how people behave, and as a result have the potential to be highly cost-effective.
It is these key features that motivate continued development of gene drive approaches to malaria control. As with any new technology for public health, any specific construct that is put forward for release must be scrutinised by regulators, other stakeholders, and the general public, and we strongly endorse efforts in this direction.
Disclosure statement
No potential conflict of interest was reported by authors.
Notes on contributors
Austin Burt is Professor of Evolutionary Genetics at Imperial College London.
Mamadou Coulibaly is a Vector Biologist at the University of Sciences, Techniques and Technologies of Bamako, Mali.
Andrea Crisanti is Professor of Molecular Parasitology at Imperial College London.
Abdoulaye Diabate is a Vector Ecologist at l''Institut de Recherche en Sciences de la Sante, Burkina Faso. He is the head of the medical entomology laboratory.
Jonathan Kayondo is a Senior Research Officer (Population Biologist) at the Uganda Virus Research Institute, Entebbe, Uganda, with a training background in Biochemistry, Chemistry and Molecular Genetics.
ORCID
Mamadou Coulibaly http://orcid.org/0000-0001-9325-2845
Additional information
Funding
References
- Adelman, Z. N. ed. 2016. Genetic Control of Malaria and Dengue. London: Academic Press.
- Akbari, O. S., C.-H. Chen, J. M. Marshall, H. Huang, I. Antoshechkin, and B. A. Hay. 2014. “Novel Synthetic Medea Selfish Genetic Elements Drive Population Replacement in Drosophila; A Theoretical Exploration of Medea – Dependent Population Suppression.” ACS Synthetic Biology 3 (12): 915–928. doi:10.1021/sb300079h.
- Akbari, O. S., K. D. Matzen, J. M. Marshall, H. Huang, C. M. Ward, and B. A. Hay. 2013. “A Synthetic Gene Drive System for Local, Reversible Modification and Suppression of Insect Populations.” Current Biology 23 (8): 671–677. doi:10.1016/j.cub.2013.02.059.
- Alonso, P. L., G. Brown, M. Arevalo-Herrera, F. Binka, C. Chitnis, F. Collins, O. K. Doumbo, et al. 2011. “A Research Agenda to Underpin Malaria Eradication.” PLoS Medicine 8 (1): e1000406. doi:10.1371/journal.pmed.1000406.
- Alphey, L., M. Benedict, R. Bellini, G. G. Clark, D. A. Dame, M. W. Service, and S. L. Dobson. 2010. “Sterile-insect Methods for Control of Mosquito-borne Diseases: An Analysis.” Vector-Borne and Zoonotic Diseases 10 (3): 295–311. doi:10.1089/vbz.2009.0014.
- Bayoh, M. N., D. K. Mathias, M. R. Odiere, F. M. Mutuku, L. Kamau, J. E. Gimnig, J. M. Vulule, W. A. Hawley, M. J. Hamel, and E. D. Walker. 2010. “Anopheles gambiae: Historical Population Decline Associated with Regional Distribution of Insecticide-treated Bed Nets in Western Nyanza Province, Kenya.” Malaria Journal 9. doi:10.1186/1475-2875-9-62.
- Beaghton, A., P. J. Beaghton, and A. Burt. 2016. “Gene Drive Through a Landscape: Reaction-Diffusion Models of Population Suppression and Elimination by a Sex Ratio Distorter.” Theoretical Population Biology 108: 51–69. doi:10.1016/j.tpb.2015.11.005.
- Beaghton, A., A. Hammond, T. Nolan, A. Crisanti, H. C. J. Godfray, and A. Burt. 2017. “Requirements for Driving Antipathogen Effector Genes into Populations of Disease Vectors by Homing.” Genetics 205 (4): 1587–1596. doi:10.1534/genetics.116.197632.
- Burt, A. 2003. “Site-specific Selfish Genes as Tools for the Control and Genetic Engineering of Natural Populations.” Proceedings of the Royal Society B: Biological Sciences 270 (1518): 921–928. doi:10.1098/rspb.2002.2319.
- Burt, A. 2014. “Heritable Strategies for Controlling Insect Vectors of Disease.” Philosophical Transactions of the Royal Society B-Biological Sciences 369 (1645). doi:10.1098/rstb.2013.0432.
- Burt, A., and R. Trivers. 2006. Genes in Conflict: The Biology of Selfish Genetic Elements. Cambridge: Belknap Press of Harvard University Press.
- Catteruccia, F., J. P. Benton, and A. Crisanti. 2005. “An Anopheles Transgenic Sexing Strain for Vector Control.” Nature Biotechnology 23 (11): 1414–1417. doi:10.1038/nbt1152.
- Champer, J., R. Reeves, S. Y. Oh, C. Liu, J. X. Liu, A. G. Clark, and P. W. Messer. 2017. “Novel CRISPR/Cas9 Gene Drive Constructs Reveal Insights into Mechanisms of Resistance Allele Formation and Drive Efficiency in Genetically Diverse Population.” PLoS Genetics 13 (7). doi:10.1371/journal.pgen.1006796.
- Chan, Y.-S., D. S. Huen, R. Glauert, E. Whiteway, and S. Russell. 2013. “Optimising Homing Endonuclease Gene Drive Performance in a Semi-refractory Species: The Drosophila melanogaster Experience.” Plos One 8 (1). doi:10.1371/journal.pone.0054130.
- Chan, Y.-S., D. A. Naujoks, D. S. Huen, and S. Russell. 2011. “Insect Population Control by Homing Endonuclease-based Gene Drive: An Evaluation in Drosophila melanogaster.” Genetics 188 (1): 33–44. doi:10.1534/genetics.111.127506.
- Chen, C. H., H. X. Huang, C. M. Ward, J. T. Su, L. V. Schaeffer, M. Guo, and B. A. Hay. 2007. “A Synthetic Maternal-effect Selfish Genetic Element Drives Population Replacement in Drosophila.” Science 316 (5824): 597–600. doi: 10.1126/science.1138595
- David, A. S., J. M. Kaser, A. C. Morey, A. M. Roth, and D. A. Andow. 2013. “Release of Genetically Engineered Insects: A Framework to Identify Potential Ecological Effects.” Ecology and Evolution 3 (11): 4000–4015. doi:10.1002/ece3.737.
- Deredec, A., A. Burt, and H. C. J. Godfray. 2008. “The Population Genetics of Using Homing Endonuclease Genes in Vector and Pest Management.” Genetics 179 (4): 2013–2026. doi:10.1534/genetics.108.089037.
- Deredec, A., H. C. J. Godfray, and A. Burt. 2011. “Requirements for Effective Malaria Control with Homing Endonuclease Genes.” Proceedings of the National Academy of Sciences 108 (43): E874–E880. doi:10.1073/pnas.1110717108.
- Dujon, B. 1989. “Group I Introns as Mobile Genetic Elements — Facts and Mechanistic Speculations — A Review.” Gene 82: 91–114. doi: 10.1016/0378-1119(89)90034-6
- Eckhoff, P. A., E. A. Wenger, H. C. J. Godfray, and A. Burt. 2017. Impact of Mosquito Gene Drive on Malaria Elimination in a Computational Model with Explicit Spatial and Temporal Dynamics.” Proceedings of the National Academy of Sciences 114 (2): E255–E264. doi:10.1073/pnas.1611064114.
- EFSA. 2013. “Guidance on the Environmental Risk Assessment of Genetically Modified Animals.” EFSA Journal 11 (5): 3200. doi:10.2903/j.efsa.3200 doi: 10.2903/j.efsa.2013.3200
- Esvelt, K. M., A. L. Smidler, F. Catteruccia, and G. M. Church. 2014. “Concerning RNA-guided Gene Drives for the Alteration of Wild Populations.” Elife 3, doi:10.7554/eLife.03401.
- Galizi, R., L. A. Doyle, M. Menichelli, F. Bernardini, A. Deredec, A. Burt, B. L. Stoddard, N. Windbichler, and A. Crisanti. 2014. “A Synthetic Sex Ratio Distortion System for the Control of the Human Malaria Mosquito.” Nature Communications 5. doi:10.1038/ncomms4977.
- Gantz, V. M., and E. Bier. 2015. “The Mutagenic Chain Reaction: A Method for Converting Heterozygous to Homozygous Mutations.” Science 348 (6233): 442–444. doi:10.1126/science.aaa5945.
- Gantz, V. M., N. Jasinskiene, O. Tatarenkova, A. Fazekas, V. M. Macias, E. Bier, and A. A. James. 2015. “Highly Efficient Cas9-mediated Gene Drive for Population Modification of the Malaria Vector Mosquito Anopheles stephensi.” Proceedings of the National Academy of Sciences 112 (49): E6736–3E43. doi:10.1073/pnas.1521077112.
- Godfray, H. C. J., A. North, and A. Burt. 2017. "How Driving Endonuclease Genes can be Used to Combat Pests and Disease Vectors.” BMC Biology 15. doi:10.1186/s12915-017-0420-4.
- Griffin, J. T., S. Bhatt, M. E. Sinka, P. W. Gething, M. Lynch, E. Patouillard, E. Shutes, et al. 2016. “Potential for Reduction of Burden and Local Elimination of Malaria by Reducing Plasmodium falciparum Malaria Transmission: A Mathematical Modelling Study.” The Lancet Infectious Diseases 16 (4): 465–472. doi:10.1016/s1473-3099(15)00423-5.
- Hammond, A., R. Galizi, K. Kyrou, A. Simoni, C. Siniscalchi, D. Katsanos, M. Gribble, et al. 2016. “A CRISPR-Cas9 Gene Drive System Targeting Female Reproduction in the Malaria Mosquito Vector Anopheles gambiae.” Nature Biotechnology 34 (1): 78–83. doi:10.1038/nbt.3439.
- Hammond, A. M., K. Kyrou, M. Bruttini, A. North, R. Galizi, X. Karlsson, N. Kranjc, F. M. Carpi, R. D'Aurizio, and A. Crisanti. 2017. “The Creation and Selection of Mutations Resistant to a Gene Drive over Multiple Generations in the Malaria Mosquito.” PLoS Genetics 13 (10). doi:10.1371/journal.pgen.1007039.
- Krzywinska, E., N. J. Dennison, G. J. Lycett, and J. Krzywinski. 2016. “A Maleness Gene in the Malaria Mosquito Anopheles gambiae.” Science 353 (6294): 67–69. doi:10.1126/science.aaf5605.
- Marshall, J. M., and O. S. Akbari. 2016. “Gene Drive Strategies for Population Replacement.” In Genetic Control of Malaria and Dengue, edited by Z.N. Adelman, 169–200. London: Academic Press.
- Marshall, J. M., A. Buchman, H. M. Sánchez, and O. S. Akbari. 2017. “Overcoming Evolved Resistance to Population-Suppressing Homing-Based Gene Drives.” Scientific Reports 7 (3776). doi:10.1038/s41598-017-02744-7.
- NASEM (National Academies of Sciences, Engineering and Medicine). 2016. Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values. Washington, DC: The National Academies Press.
- Newton, M. E., R. J. Wood, and D. I. Southern. 1976. “A Cytogenetic Analysis of Meiotic Drive in the Mosquito, Aedes aegypti (L.).” Genetica 46 (3): 297–318. doi: 10.1007/BF00055473
- North, A., A. Burt, and H. C. J. Godfray. 2013. “Modelling the Spatial Spread of a Homing Endonuclease Gene in a Mosquito Population.” Journal of Applied Ecology 50 (5): 1216–1225. doi:10.1111/1365-2664.12133.
- Papathanos, P. A., N. Windbichler, M. Menichelli, A. Burt, and A. Crisanti. 2009. “The Vasa Regulatory Region Mediates Germline Expression and Maternal Transmission of Proteins in the Malaria Mosquito Anopheles gambiae: A Versatile Tool for Genetic Control Strategies.” BMC Molecular Biology, 10–65. doi:10.1186/1471-2199-10-65.
- Russell, T. L., N. J. Govella, S. Azizi, C. J. Drakeley, S. P. Kachur, and G. F. Killeen. 2011. “Increased Proportions of Outdoor Feeding among Residual Malaria Vector Populations Following Increased Use of Insecticide-treated Nets in Rural Tanzania.” Malaria Journal 10. doi:10.1186/1475-2875-10-80.
- Simoni, A., C. Siniscalchi, Y.-S. Chan, D. S. Huen, S. Russell, N. Windbichler, and A. Crisanti. 2014. “Development of Synthetic Selfish Elements Based on Modular Nucleases in Drosophila melanogaster.” Nucleic Acids Research 42 (11): 7461–7472. doi:10.1093/nar/gku387.
- Smith, D. L., and F. E. McKenzie. 2004. “Statics and Dynamics of Malaria Infection in Anopheles Mosquitoes.” Malaria Journal 3. doi:10.1186/1475-2875-3-13.
- Sreenivasamurthy, S. K., G. Dey, M. Ramu, M. Kumar, M. K. Gupta, A. K. Mohanty, H. C. Harsha, et al. 2013. “A Compendium of Molecules Involved in Vector-pathogen Interactions Pertaining to Malaria.” Malaria Journal 12. doi:216 10.1186/1475-2875-12-216.
- Tatem, A. J., P. W. Gething, D. L. Smith, and S. I. Hay. 2013. “Urbanization and the Global Malaria Recession.” Malaria Journal 12. doi:10.1186/1475-2875-12-133.
- van Driesche, R., M. Hoddle, and T. Center. 2008. Control of Pests and Weeds by Natural Enemies: An Introduction to Biological Control. Wiley-Backwell.
- Vella, M. R., C. E. Gunning, A. L. Lloyd, and F. Gould. 2017. “Evaluating Strategies for Reversing CRISPR-Cas9 Gene Drives.” Scientific Reports 7 (11038). doi:10.1038/s41598-017-10633-2.
- Wang, S., and M. Jacobs-Lorena. 2013. “Genetic Approaches to Interfere with Malaria Transmission by Vector Mosquitoes.” Trends in Biotechnology 31: 185–193. doi: 10.1016/j.tibtech.2013.01.001
- WHO. 2010. “Progress and Prospects for the Use of Genetically Modified Mosquitoes to Inhibit Disease Transmission.” Geneva: FNIH/WHO/TDR.
- WHO. 2015. “World Malaria Report 2015.” Geneva: WHO.
- WHO. 2016. “Global Technical Strategy for Malaria 2016-2030.” Geneva.
- Windbichler, N., M. Menichelli, P. A. Papathanos, S. B. Thyme, H. Li, U. Y. Ulge, B. T. Hovde, et al. 2011. “A Synthetic Homing Endonuclease-based Gene Drive System in the Human Malaria Mosquito.” Nature 473 (7346): 212–215. doi:10.1038/nature09937.
- Wu, B., L. Luo, and X. J. Gao. 2016. “Cas9-triggered Chain Ablation of CAS9 as a Gene Drive Brake.” Nature Biotechnology 34 (2): 137–138. doi:10.1038/nbt.3444.
- Zeddies, J., R. P. Schaab, P. Neuenschwander, and H. R. Herren. 2001. “Economics of Biological Control of Cassava Mealybug in Africa.” Agricultural Economics 24 (2): 209–219. doi: 10.1111/j.1574-0862.2001.tb00024.x
