Abstract
Bryophytes and lichens are ubiquitous in subarctic ecosystems, but their roles in controlling energy fluxes are rarely studied at the species level despite large, recent observed shifts in subarctic vegetation. We quantified the surface and subsurface temperatures and spectral reflectance of common moss and lichen species at field sites in Alaska and Sweden. We also used MODIS observations to determine if the removal of Cladonia spp. by reindeer overgrazing impacts land surface albedo and temperature. Radiometric surface temperature of a feather moss (Pleurozium schreberi) exceeded 50 °C on occasion when dry, up to 20 °C higher than co-located Sphagnum fuscum or C. rangiferina. Spectral reflectance of S. fuscum was on average higher than Polytrichum piliferum across the 350–1400 nm range, with substantial within-species variability. MODIS albedo was significantly higher on the Norwegian (relatively undisturbed) side versus the Finnish (disturbed) side of a border reindeer fence by an average of 1% during periods without snow cover. MODIS nighttime land surface temperatures were often significantly higher on the Norwegian side of the fence by an average of 0.7 °C despite higher albedo, likely due to poor conductance of heat to the subsurface as observed in C. rangiferina in the field. Changes to bryophyte and lichen community composition alter the surface energy balance, and future work must determine how to best incorporate these effects into Earth system models.
Introduction
Bryophytes and lichens often dominate the surface of boreal and tundra ecosystems (CitationDuring and Tooren, 1990; CitationEsseen et al., 1997) and as a consequence are commonly associated with permafrost and/or carbon-rich soils (CitationZimov et al., 2005; CitationSchuur et al., 2008; CitationTarnocai et al., 2009). The transport of heat from the land surface determines in part the thermal environment of soil, and the partitioning of energy at the land surface determines in part regional climate (CitationBonan et al., 1990; CitationBeringer et al., 2005; CitationBonan, 2008). Controls on the subarctic surface energy balance by bryophytes and lichens must be quantified for a comprehensive understanding of the surface and subsurface thermal environment and biosphere-atmosphere energy exchange in a changing climate (CitationBeringer et al., 2001).
In process-based studies of permafrost thaw, mosses are often thought of as an insulating surface layer, whose thermal properties are lost in the instance of fire or other disturbances (CitationBlok et al., 2011). Bryophyte and lichen species are rarely incorporated into models of ecosystem biogeochemistry or physics (CitationBeringer et al., 2001; CitationCornelissen et al., 2007), due in part to a lack of data and the pronounced spatial variability of the species composition and water content of many subarctic ecosystems (CitationAsner et al., 2003). We hold that the thermal characteristics of bryophyte and lichen species may differ sufficiently to warrant a species-level characterization for modeling studies (CitationKershaw, 1978; CitationVogelmann and Moss, 1993; CitationO'Donnell et al., 2009; CitationElumeeva et al., 2011). Studies of surface and subsurface temperatures and subsurface heat fluxes of bryophytes and lichens at the species level has rarely been undertaken to date despite extensive work on the radiative properties of bryophytes versus other surface cover types (CitationKershaw, 1985; CitationRees, 1993), transmission of radiation through bryophyte canopies and associated carbon dynamics (CitationSkre et al., 1983), and characterization of bryophyte spectral reflectance (CitationPetzold and Rencz, 1975; CitationVogelmann and Moss, 1993). A study by Street (Citation2011) however, indicated large differences in surface temperature and heat flux between moss species growing in neighboring patches in subarctic Finland.
To begin progress on bryophyte and lichen energy flux partitioning at the species level, we measured surface and subsurface temperatures and modeled subsurface heat flux in common bryophytes and lichens under field conditions in an area of extensive permafrost degradation in the Innoko National Wildlife Refuge, Alaska. For a more comprehensive study of bryophyte radiative properties, we measured the spectral reflectance of mosses and soils from a tundra ecosystem in Abisko, Sweden. To explore the potential importance of bryophyte and lichen community composition in the Earth system, we investigated differences in albedo and land surface temperature products from the moderate resolution imaging spectroradiometer (MODIS) along a Cladonia spp. disturbance gradient on the Norway/Finland border (CitationKayhko and Pellikka, 1994).
We hypothesize that surface temperatures will differ among bryophyte and lichen species due to differences in albedo, waterholding capacity, and evaporative cooling among species (CitationNichols and Brown, 1980; CitationKim and Verma, 1996; CitationLiljedahl et al., 2011; CitationStreet, 2011). We test this experimental hypothesis using field observations of radiometric surface temperature (Tsurf ), subsurface temperature, spectral reflectance of moss patches, and models of subsurface heat flux (G). We further predict that changes to land surface albedo and temperature from changes in bryophyte community composition as a result of reindeer grazing will be pronounced enough to distinguish from space, and test this prediction using remote sensing data products from MODIS.
Methods
We first describe the study sites and field observations, followed by an explanation of the remote sensing analysis, as well as theoretical considerations regarding G, the surface energy balance, and Tsurf . Results from multiple sites are used to overcome challenges of data acquisition in remote locations.
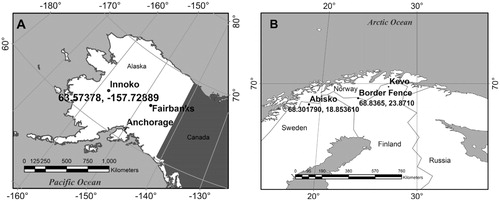
INNOKO
Innoko Flats National Wildlife Refuge (hereafter Innoko) is located in interior Alaska (, part A). The study area was located at 63.57378°N, 157.72889°W. Characteristic patches of Sphagnum fuscum, Cladonia rangiferina, and Pleurozium schreberi occupying similar microtopographic positions on the top of hummocks and co-located within meters of each other with minimal shading from the sparse black spruce (Picea mariana) overstory were chosen for this analysis. Tsurf was measured every minute during the study period from day of year (DOY) 149 to 155, 2011, using a SI-111 infrared radiometer (Apogee Instruments, Logan, Utah, U.S.A.) mounted on a tripod. The radiometer was pointed approximately at nadir to all patches, noting microtopographical variability, at a distance of 20–25 cm. The SI-111 has a 22° half angle field of view and is sensitive within a wavelength range of 8 to 14 µm. Measurements made while moving and immediately after moving the tripod to different patches were discarded. Subsurface temperature at 1 cm (T1 cm ) and 5 cm (T5 cm ) was measured every minute using Type E (chromel-constantan) thermocouples, and all data were logged using a Campbell CR1000 datalogger (Campbell Scientific, Logan, Utah). A thirty-minute digital filter was chosen for graphical representation of the continuous subsurface temperature measurements.
FIGURE 1. Maps of the (A) Innoko, Alaska, study area and the (B) Abisko, Sweden, and Norway/Finland Border Fence study areas.
ABISKO
Spectral reflectance of Sphagnum fuscum, Polytrichum piliferum, and soil was measured using an ASD Field Spectrometer (ASD Instruments, Boulder, Colorado, U.S.A.) in a subarctic tundra landscape near Abisko, Sweden, centered around 68.301790°N, 18.853610°E. Measurements were made during sunny periods in July 2007 on four plots of S. fuscum, three plots of P. piliferum, two patches of unsaturated soil, and one patch of saturated soil. Vegetative and edaphic characteristics of the study area are described in Fox et al. (Citation2008), Shaver et al. (Citation2007), Street et al. (Citation2007; Citation2011b), Spadavecchia et al., (Citation2008), and Williams et al. (Citation2008).
REMOTE SENSING
Reindeer/caribou (Rangifer tarandus) overgrazing can sharply diminish the abundance of slow-growing lichen communities (CitationHenry and Gunn, 1991). An area of particular interest for the study of the impacts of reindeer overgrazing on Cladonia spp. is a fence on the Norway/Finland border (, part B), where different seasonal reindeer management practices prevail (CitationKayhko and Pellikka, 1994). Year-round reindeer management on the Finnish side of the border has severely reduced Cladonia spp. presence, but winter-only grazing on the Norwegian side has maintained Cladonia spp. presence despite higher average annual grazing pressure (CitationKayhko and Pellikka, 1994). We studied the 25 km2 area centered around 68.8365°N, 23.8710°E described by the UNEP/GRID-Arendal Maps and Graphics Library (2010).
Albedo and land surface temperature (abbreviated ‘LST’ for continuity with the MODIS naming conventions) of the Norway/ Finland border fence area were explored using the MODIS MCD34A1 and MYD11A2 products. The MCD34A1 product is a 500 m 16-day composite of the Bidirectional Reflectance Distribution Function (BRDF)/Albedo parameters. Actual albedo values were computed from the MCD34A1 product following Schaaf et al. (Citation2002). The MYD11A2 product is a one km 8-day composite of day/night land surface temperature and emissivity characteristics. Observations for the time frame of 2001 until 2011 were retrieved using the MODIS server client and were filtered using the associated quality control products. Pixels that intersect the Norway/ Finland border, and pixels adjacent to those pixels, were removed from the analysis. To focus on the snow-free season, pixels with albedo values >0.3 that may be contaminated by snow presence were removed from the MODIS albedo and LST analyses.
MODELS FOR SUBSURFACE HEAT FLUX
In the absence of direct measurements, the estimation of subsurface heat flux (G) follows from the continuity equation:
where t is time, T is temperature, z is depth, ρs is soil density, cs is soil volumetric heat capacity, and k is thermal conductivity. Some inference regarding the soil heat flux parameters is required for an estimate of G, because these are difficult to obtain without resorting to destructive measurements.
Campbell and Norman (Citation1998) offered a simplification for the case of deep soils subject to a sinusoidal temperature forcing:
where A is the amplitude of diurnal Tsurf fluctuations, t0 is a phase shift that depends on the time reference, D is damping depth, and ω is the angular frequency which equals 7.3 × 10—5S—1 for diurnal fluctuations. D can be estimated using
where Ax represents the amplitude of diurnal soil temperature fluctuations at depth x, here 1 cm and 5 cm. D represents the depth at which the amplitude of temperature changes is 37%, 1/e, of that at the surface. From the surface and subsurface temperature measurements at Innoko, the only unknown is k, and we explore a range of values between the minimum (ca. 0.05 W m—1 K—1) and maximum (ca. 0.55 W m—1 K—1 for Sphagnum spp. and 0.25 W m-1 K—1 for feather mosses) quantified by O'Donnell et al. (2009) to estimate G. Cladonia rangiferina was assumed to share a similar range of k as feather mosses.
THEORETICAL CONSIDERATIONS
Explanations for the partitioning of energy toward or away from G follow from the net radiation balance:
where Rn is the net radiation, H is sensible heat flux, λE is latent heat exchange, and J is energy storage, assumed to be negligible here. Expanding the turbulent flux terms H and λE results in:
where gH and gv are conductances for heat and water vapor respectively, cp is the specific heat of air at constant pressure, pa is atmospheric pressure, Ts is aerodynamic surface temperature, Ta is air temperature, λ is the latent heat of vaporization, es [Ts ] is the saturation vapor pressure of water, and ea is the partial pressure of water in air. Tsurf is related to G via Equation 1, and to H and λE through its relationship to Ts (CitationNorman et al., 1995).
Equation (4) can be rewritten to include incident ↓ and outgoing ↑ shortwave (SW) and longwave (LW) energy flux:
and expanded to incorporate the shortwave albedo, α:
Systems with higher albedo will have lower Rn , all else being equal. Tsurf itself is related to LW via the Stefan-Boltzmann Law for a gray body:
where A is surface area, σ is the Stefan-Boltzmann constant, and σ is emissivity, often assumed to be ca. 0.97 in natural environments (CitationCampbell and Norman, 1998), but in actuality a function of albedo following Kirchoff's Law of Radiation at equilibrium (see CitationChen et al., 2002). To summarize, albedo and Tsurf enter different terms in the surface energy balance, and are therefore determinants of the subsurface and atmospheric energy budget for which the surface represents a boundary.
Results
INNOKO: SURFACE AND SUBSURFACE TEMPERATURE
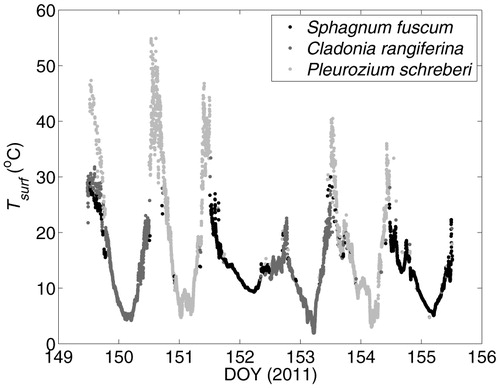
Prevailing weather conditions were warm and dry during the earlier part of the measurement period before DOY 152 at Innoko, which was followed by overcast skies with intermittent rain events. Tsurf reached over 55 °C in P. schreberi, but rarely reached over 30 °C in S. fuscum or C. rangiferina (). Tsurf of P. schreberi exceeded the other bryophytes by nearly 20 °C during warm, sunny periods, but by no more than ca. 5 °C during cool, wet periods ().
FIGURE 2. Radiometric surface temperature (Tsurf ) of representative Sphagnum fuscum, Cladonia rangiferina, and Pleurozium schreberi patches in the Innoko National Wildlife Refuge, Alaska.
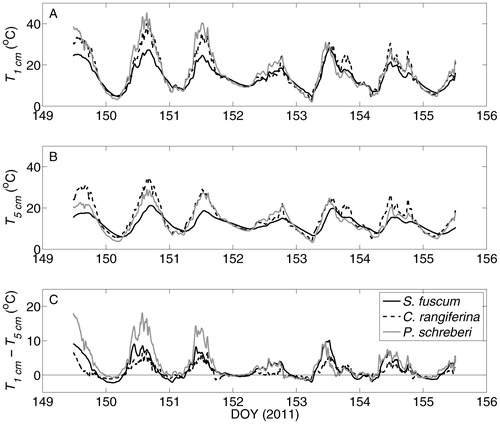
T1 cm in P. schreberi likewise tended to exceed that of C. rangiferina and S. fuscum, the latter commonly by more than 10 °C, during morning and midday periods (, part A). T1 cm among species converged at night and after rain events (, part A). T5 cm was often greatest in C. rangiferina during dry daytime periods (, part B) such that the difference between T1 cm and T5 cm was usually lowest in C. rangiferina during the day (, part C), with implications for G ( and Equation 2). Mean T1 cm and T5 cm during the study period are significantly different among species (two-sided t-test, p < 0.05). The mean and standard deviation of T1cm was 14.2 ± 5.8 °C in S. fuscum, 16.2 ± 8.5 °C in C. rangiferina, and 16.7 ± 9.6 °C in P. schreberi. T5 cm over the measurement period was 12.4 ± 3.6 °C in S. fuscum, 15.2 ± 7.0 °C in C. rangiferina, and 13.6 ± 6.0 °C in P. schreberi.
FIGURE 3. Temperature at 1 cm (T1 cm , A) and 5 cm (T5cm, B) below the surface of Sphagnum fuscum, Cladonia rangiferina, and Pleurozium schreberi patches in the Innoko National Wildlife Refuge, Alaska. Subplot C represents the difference between subplots A and B.
INNOKO: MODELED SUBSURFACE HEAT FLUX
Values of D calculated from observations () and used for the estimation of G are listed in . The ranges of modeled G varied between ca. ± 200 W m—2 for S. fuscum, between ca. ± 100 W m—2 for C. rangiferina, and between ca. ± 150 W m—2 for P. shreberi (). Following O'Donnell et al. (2009), k was likely low when mosses and lichens were dry before DOY 152, and was likely higher after the precipitation events. Consequently, the magnitude of G was likely on the low end of the modeled range for all species during the early part of the measurement period, and on the higher end of the modeled range during the latter part of the measurement period.
FIGURE 4. Estimated subsurface heat flux (G) in representative (A) Sphagnum fuscum, (B) Cladonia rangiferina, and (C) Pleurozium schreberi patches in the Innoko National Wildlife Refuge, Alaska. The line widths reflect uncertainty in thermal conductivity (k), which were taken to represent a range of published values (O'Donnell et al., 2009). Positive values reflect energy flux from surface to subsurface.
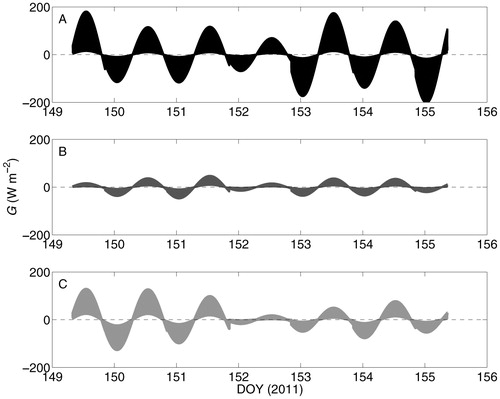
TABLE 1 Modeled damping depth (m) using Equation (3) as a function of day of year (2011) for the Sphagnum fuscum, Cladonia rangiferina, and Pleurozium schreberi study sites at Innoko, Alaska.
ABISKO: SPECTRAL REFLECTANCE
Reflectance in S. fuscum tended to be greater than that of P. piliferum and bare ground across the visible wavebands and the near infrared wavebands up to ca. 1300 nm (). A spike in reflectance in the red section of the visible spectrum (620–730 nm) was observed in S. fuscum. P. piliferum reflected on average more radiation than S. fuscum across longer wavelengths in the near infrared spectrum, taken here to be 1400–1900 nm. Reflectance of the two moss species was typically <10% at wavelengths between 1900 and 2400 nm. Bare ground patches reflected less than ca. 20% of radiation across the entire visible to near-infrared spectrum, but reflected more radiation than the bryophytes at wavelengths longer than 1400 nm. Saturated bare ground reflected less radiation than the moss species across the entire observed spectrum.
FIGURE 5. The mean and standard deviation of the spectral reflectance of four plots of Sphagnum fuscum, three plots of Polytrichum piliferum, two representative patches of bare ground, and a patch of saturated bare ground in a tundra ecosystem near Abisko, Sweden.
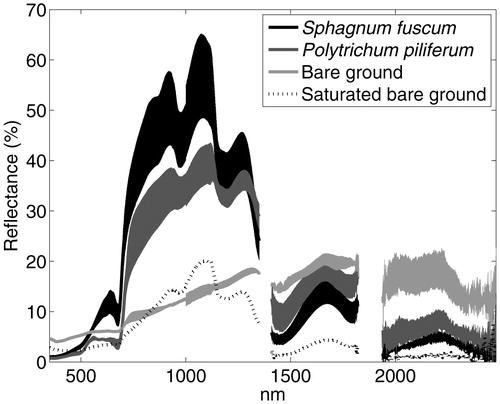
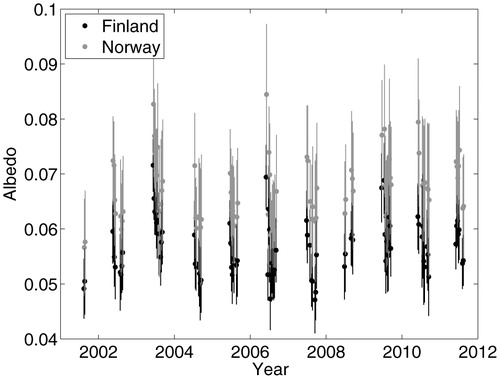
FIGURE 6. The mean and standard deviation of shortwave albedo calculated from the MODIS MCD34A1 product following Schaaf et al. (Citation2002) for Finnish and Norwegian sides of a Cladonia disturbance gradient along a border fence.
NORWAY/FINLAND BORDER FENCE: MODIS
Mean MODIS albedo was significantly higher on the Norwegian side of the border fence (0.071) than the Finnish side of the border fence (0.061) during all snow-free periods in the measurement record (p < 0.05; ).
Across all quality-controlled MODIS pixels (2001–2011), snow-free nighttime LST on the Norwegian side of the border fence (LSTNorway 3.2 ± 4.5 °C) was significantly higher than all values from the Finnish side of the fence (LST Finland .5 ± 4.3 °C). Mean nighttime LST Norway was significantly higher than nighttime LST Finland during 25 of the 63 MODIS observations with sufficient number of pixels that passed quality control criteria to compute statistics, and mean nighttime LST finland was significantly higher than mean nighttime LST Norway during 7 of the 63 MODIS observations.
A sufficient number of daytime LST MODIS pixels to support statistical analyses were available on only 14 instances during snow-free periods. The mean of all acceptable daytime LST observations were not significantly different between the Norwegian (6.4 ± 6.4 °C) and Finnish (6.9 ± 7.1 °C) sides of the fence. Mean daytime LST Norway (LST finland ) was significantly higher than daytime LST Finland on 2 of 14 occasions, and mean daytime LST Finland was significantly higher than daytime LST Norway on 2 of 14 occasions with sufficient number of pixels to compute statistics.
Discussion
SPHAGNUM FUSCUM
S. fuscum maintained cooler maximum Tsurf than P. schreberi during most of the measurement record at Innoko, and maintained slightly cooler maximum Tsurf than C. rangiferina during the dry early period (). The extremes of modeled G in S. fuscum were often larger than those of C. rangiferina and P. schreberi (), reflecting high potential values of k (O'Donnell et al., 2009). Although Sphagnum spp. associated with wetland ecosystems demonstrate large latent heat losses (CitationKim and Verma, 1996), latent heat flux is often lower than that of vascular plant species despite trivial resistance to latent heat losses (CitationVogelmann and Moss, 1993; CitationKim and Verma, 1996) due to the lack of active water transport from deeper layers to the surface (CitationOechel and van Cleve, 1986; CitationLiljedahl et al., 2011).
Near infrared reflectance of Sphagnum fuscum measured in Abisko () exceeded laboratory observations of other Sphagnum species made by Vogelmann and Moss (Citation1993), which did not exceed 40% reflectance across the infrared wavelengths. Vogelmann and Moss (Citation1993) demonstrated that Sphagnum spp. albedo is a function of moisture status by observing a strong increase in reflectance across the visible to near-infrared spectrum in Sphagnum cuspidatum during a drying experiment. Field-based spectroscopy in Abisko was made during favorable, well-watered periods during the growing season, and reflectance exhibited considerable variability among patches, which may have resulted from landscape position and moisture status. Measuring moisture status nondestructively in mosses and organic soils across both space and time is an ongoing technical challenge.
CLADONIA RANGIFERINA
C. rangiferina maintained a small gradient between T1 cm and T5cm below the surface during the measurement period at Innoko compared to other study species (). The extremes of modeled subsurface heat flux were accordingly lower than that of both S. fuscum and P. schreberi (), noting that representative k values from P. schreberi were used here (O'Donnell et al., 2009). In actuality, the magnitude of G may be even lower if stagnant air layers, with a k of ca. 0.025 W m—1 K—1, persist in the open areas underneath the C. rangiferina surface.
Cladonia species maintain a relatively high albedo when wet or dry (ca. 0.22 versus 0.155 for a mesic Polytrichum juniperinum—dominated meadow; CitationPetzold and Rencz, 1975), such that Rn of Cladonia-dominated surfaces will be lower than that of surfaces with lower albedo, all else being equal. During dry periods, the C. rangiferina canopy was crisp to the touch, suggesting that latent heat losses were minimal. The relatively cool radiometric surface temperature of C. rangiferina versus that of P. schreberi suggests that H was likely lower if one assumes that radiometric and aerodynamic surface temperatures are approximately equal in uniform canopies and that boundary layer and atmospheric resistances to H in the low-statured mats are similar among species (CitationNorman et al., 1995). In summary, Rn in C. rangiferina is lower than many other bryophyte and lichen species due to relatively large albedo, but less of this available energy is allocated to G than in the other species studied here. This leaves more energy available for surface heating than may have been expected given its high albedo.
PLEUROZIUM SCHREBERI
Tsurf of P. schreberi often approached temperatures in excess of 50 °C during the dry period (), which meets or exceeds that thought to be damaging to the photosynthetic machinery in vascular plants in subarctic regions (CitationLarcher, 1995) and lies above the long-recognized temperature optima for bryophytes and lichen photosynthesis (e.g. CitationLange, 1962; CitationKershaw, 1985). Optimum temperatures for net photosynthesis at light saturation in Sphagnum and feather moss species of interior Alaska is on the order of 10–15°C during late June and on the order of 20–26 °C during midAugust (CitationSkre and Oechel, 1981); the surface temperature of all species measured here meet or exceed these values during a dry period in late May (), noting the difference between leaf internal and surface temperature. Tsurf , T1 cm , and T5 cm in P. schreberi were often greater than that of S. fuscum ( and ), suggesting that bryophyte species composition, rather than mere presence/ absence, plays an important role in the thermal budget of boreal and tundra soils. It is important to note that feather moss species are commonly associated with forest understories (CitationNilsson and Wardle, 2005) and avoid high radiation loads when shaded in these habitats.
NORWAY/FINLAND BORDER FENCE
Tsurf (), G (), and albedo (; CitationPetzold and Rencz, 1975) differ among bryophyte and lichen types. Bryophyte and lichen community composition thus may have implications for all energy flux terms in Equation 4, but it is unclear if community composition alone has a discernable impact on land surface characteristics of importance to climate. The Norway/Finland border fence offers a test of the role of bryophyte and lichen community structure in surface-atmosphere energy flux; C. rangiferina was for all intents and purposes removed by reindeer from the Finnish side of the fence by grazing management practices (CitationKayhko and Pellikka, 1994). Cladonia spp. and other lichen species are on the decline across the Arctic and subarctic (CitationJoly et al., 2009), due in part to R. tarandus grazing pressure (CitationJoly et al., 2007).
Albedo was consistently higher on the Norwegian side of the border fence during the snow-free season () by ca. 0.01 on average. This figure may appear small, but a 1% difference is substantial given representative ranges of variability in forest albedo, which rarely exceed 0.2 or fall below 0.1 for deciduous forests during the growing season (CitationHollinger et al., 2010). From this point of view, a 1% change in albedo is approximately a 10% change over the possible ranges of albedo found in deciduous forests during the growing season. Obvious changes in reflectance across the visible bands (CitationKayhko and Pellikka, 1994) and significant changes to shortwave albedo () from Cladonia spp. presence is measurable from space.
From the Earth system perspective, changes to snow extent and duration or tree and shrub cover (CitationBetts, 2000; CitationChapin et al., 2005; CitationHollinger et al., 2010) are likely to play a larger role in land surface albedo than changes to bryophyte and lichen community composition in subarctic ecosystems. For example, Euskirchen et al. (Citation2009) found that changes in summer albedo of 0.6% over a 100 year post-fire successional trajectory in Alaskan boreal forests is associated with a -0.9 W m—2 decade-1 climate forcing, but a 4.3 W m—2 decade—1 forcing from projected changes in the length of the snow-free season to the year 2100 is predicted. Our observed 1% decrease in surface albedo with Cladonia spp. removal corresponds to a > 1 W m—2 decade—1 decrease in climate forcing if these changes occur on the same time scales as those studied by Euskirchen et al. (Citation2009). Bryophyte and lichen species composition may also play an important role in snow capture and melt; Street et al. (Citation2011a) observed that patches of Polytrichum spp. at Abisko were the first to thaw in the spring on account of their microtopographical position on the tops of small ridges and their low albedo (). The small-scale patterns of snowmelt and albedo change, on the order of centimeters to meters, lie far below the spatial resolution of MODIS and most other remote sensing products.
Increases in vegetation greenness and shrub abundance have contributed to a net warming of the land surface in the Arctic (CitationJia et al., 2003; CitationChapin et al., 2005), but studies to date have only focused on vascular vegetation. Along the Norway/Finland border fence, LST Norway was often higher than LST Finland , especially at night, suggesting a slight surface cooling from C. rangiferina overgrazing. These observations are consistent with the observation that Tsurf of C. rangiferina was often higher than in S. fuscum, which can be explained in part by the reduced transport of heat to the subsurface in C. rangiferina ( and ). In other words, the counterintuitive result that a decrease in albedo was accompanied by a decrease in LST can be understood by considering the entire surface energy balance (Equation 4) and differences in G between Cladonia spp. and other surface types. The finding that a decrease in albedo led to a decrease in LST is not unprecedented; studies of temperate forest management have found that darker forests maintain lower surface temperatures than brighter fields due to increases in latent and sensible heat flux (CitationJuang et al., 2007). An early study by Kershaw (Citation1978) found that the Cladonia stellaris phase of a post-wildfire lichen recovery time series had high albedo and large longwave radiation losses (related to Tsurf via Equation 8) compared to post-burn surfaces and the climax Stereocaulon paschale, although both lichen species are characteristically white/ off white. The energy balance, cool subsurface, and relatively warm surface of Cladonia spp. should be further investigated, especially with respect to its unique morphology, miniature white canopy, and important interaction with wildlife and reindeer herd management (CitationJoly et al., 2007, 2009).
MODELING IMPLICATIONS
Our results demonstrate that the biophysical properties of different bryophyte and lichen species can impact surface-atmosphere and surface-subsurface energy flux to a degree that is relevant for climate forcing scenarios. Moss layers are commonly included in process-based models of arctic ecosystem function (e.g. BIOME-BGC; CitationEngstrom et al., 2006; Engstrom and Hope, 2011) and are required for an accurate description of energy and water flux (CitationBeringer et al., 2001). Correctly accounting for moss impacts on subsurface energy flux is critical for models of permafrost thaw; empirical evidence demonstrates that the presence of a moss layer can delay permafrost thaw by about a month in the high Arctic (CitationGornall et al., 2007), but the role of individual species on permafrost thaw has yet to be examined. Improving existing maps of plant communities in boreal, subarctic, and arctic ecosystems (e.g. CitationWalker et al., 2005) to include estimates of moss and lichen species composition is a logical starting point for building species-level effects into ecosystem models.
Modeling the role of bryophyte and lichen species on ecosystem processes must also take into account multiple feedback processes that dictate high-latitude ecosystem change (CitationWookey et al., 2009). Bryophytes and lichens are generally expected to respond negatively to climate warming at subarctic latitudes, often due to shading by dwarf shrubs (CitationCornelissen et al., 2001; Citationvan Wijk et al., 2004; CitationWalker et al., 2006; CitationWookey et al., 2009). Bryophyte cover has also increased in response to experimental warming and nutrient addition at high arctic latitudes (CitationGordon et al., 2001; CitationHudson and Henry, 2010). Improving the representation of vegetation, including non-vascular vegetation in climate and ecosystem models, is a complex and ongoing challenge that is critical for understanding vegetation-climate interactions in an era of global change.
CONCLUSIONS AND FUTURE RESEARCH DIRECTIONS
Albedo, Tsurf and G are important components of the surface energy balance (Equations 4–7) and can differ significantly among bryophyte and lichen species both temporally (e.g. and ) and spatially (e.g. ). Future work should investigate the full surface energy balance across different bryophyte and lichen species, noting that partitioned energy flux measurements using radiometers (Equation 6) and G measurements using heat flux plates are likely to be more defensible than eddy covariance or Bowen ratio methodologies in most cases (CitationKim and Verma, 1996; CitationLiljedahl et al., 2011), owing to the complex spatial structure of bryophyte and lichen communities (CitationAsner et al., 2003). Furthermore, the open stature of Cladonia canopies may challenge conventional measurements of subsurface heat flux using soil heat flux plates if the instrument receives a direct solar radiation load under the miniature canopy. Regardless of methodological difficulties, understanding surface-subsurface and surface-atmosphere energy exchange in Arctic and subarctic ecosystems is a priority to better understand feedbacks between vegetation change, permafrost dynamics, and climate (CitationZimov et al., 2005; CitationChapin et al., 2008; CitationSchuur et al., 2008).
Despite methodological challenges, careful remote sensing applications and detailed radiometric and micrometeorological measurements can offer insights into the role played by different bryophyte species in the energy budget of the land surface. We suggest that future research should explicitly consider species composition when studying the thermal budget of boreal and arctic ecosystems and corresponding heat flux into permafrost-dominated soils.
Acknowledgments
We acknowledge funding from the National Science Foundation (‘Scaling Ecosystem Function: Novel Approaches from Max-Ent and Multiresolution,’ DBI #1021095; Collaborative Research: Impact of Permafrost Degradation on Carbon and Water in Boreal Ecosystems (EAR-0630319, January 2007—ongoing; Q. Zhuang, J. Harden, M. T. Jorgenson, R. Striegl, Y. Shur, Co-PIs); the Marie Curie Incoming International Fellowship Programme; the Natural Environmental Resource Council (U.K.); and the State of Montana. We would like to thank Carmel Johnston, Claire Treat, Torre Jorgenson, and Jennifer Harden for logistical support at Innoko. We would like to thank Mathew Disney, Phil Lewis, Tristan Quaife, and Mathew Williams for research support at Abisko, and Amy Trowbridge for valuable comments on the manuscript.
Related Research Data
References Cited
- Ahlenius, H. , 2010: Reindeer herding—Vegetation impacts (Norway and Finland),< http://maps.grida.no/graphicslib/detail/reindeerherding-vegetation-impacts-norway-and-finland_c996 >.
- Asner, G. P. , Scurlock, J. M. O. , and Hicke, J. A. , 2003: Global synthesis of leaf area index observations: implications for ecological and remote sensing studies. Global Ecology and Biogeography, 12: 191–205.
- Beringer, J. , Lynch, A. H. , Chapin, F. S., III , Mack, M. C. , and Bonan, G. B. , 2001: The representation of arctic soils in the land surface model: the importance of mosses. Journal of Climate, 14: 3324–3335.
- Beringer, J. , Chapin, F. S., III , Thompson, C. C. , and McGuire, A. D. , 2005: Surface energy exchanges along a tundra-forest transition and feedbacks to climate. Agricultural and Forest Meteorology, 131: 143–161.
- Betts, R. A. , 2000: Offset of the potential carbon sink from boreal forestation by decreases in surface albedo. Nature, 408: 187–190.
- Blok, D. , Heijmans, M. , Schaepman-Strub, G. , van Ruijven, J. , Parmentier, F. , Maximov, T. , and Berendse, F. , 2011: The cooling capacity of mosses: Controls on water and energy fluxes in a Siberian tundra site. Ecosystems, 14: 1055–1065.
- Bonan, G. , 2008: Ecological Climatology: Concepts and Applications. 2nd edition. Cambridge: Cambridge University Press, 568 pp.
- Bonan, G. B. , Shugart, H. H , and Urban, D. L. , 1990: Sensitivity of some high-latitude boreal forests to climatic parameters. Climatic Change, 16: 9–29.
- Campbell, G. S. , and Norman, J. M. , 1998: An Introduction to Environmental Biophysics. 2nd edition. New York: Springer.
- Chapin, F. S. , Sturm, M. , Serreze, M. C. , McFadden, J. P. , Key, J. R. , Lloyd, A. H. , McGuire, A. D. , Rupp, T. S. , Lynch, A. H. , Schimel, J. P. , Beringer, J. , Chapman, W. L. , Epstein, H. E. , Euskirchen, E. S. , Hinzman, L. D. , Jia, G. , Ping, C.-L. , Tape, K. D. , Thompson, C. D. C. , Walker, D. A. , and Welker, J. M. , 2005: Role of land-surface changes in Arctic summer warming. Science, 310: 657–660.
- Chapin, F. S. , Randerson, J. T. , McGuire, A. D. , Foley, J. A. , and Field, C. B. , 2008: Changing feedbacks in the climate-biosphere system. Frontiers in Ecology and the Environment, 6: 313–320.
- Chen, Y. , Sun-Mack, S. , Minnis, P. , Young, D. F. , and Smith, W. L., Jr ., 2002: Surface spectral emissivity derived from MODIS data. Hangzhou, China: SPIE 3rd International Asia-Pacific Environmental Remote Sensing Symposium 2002: Remote Sensing of the Atmosphere, Ocean, Environment, and Space.
- Cornelissen, J. H. C. , Callaghan, T. V. , Alatalo, J. M. , Michelsen, A. , Graglia, E. , Hartley, A. E. , Hik, D. S. , Hobbie, S. E. , Press, M. C. , Robinson, C. H. , Henry, G. H. R. , Shaver, G. R. , Phoenix, G. K. , Gwynn Jones, D. , Jonasson, S. , Chapin, F. S. , Molau, U. , Neill, C. , Lee, J. A. , Melillo, J. M. , Sveinbjörnsson, B. , and Aerts, R. , 2001: Global change and arctic ecosystems: Is lichen decline a function of increases in vascular plant biomass? Journal of Ecology, 89: 984–994.
- Cornelissen, J. H. C. , Lang, S. I. , Soudzilovskaia, N. A. , and During, H. J. , 2007: Comparative cryptogam ecology: a review of bryophyte and lichen traits that drive biogeochemistry. Annals of Botany, 99: 987–1001.
- During, H. J. , and Tooren, B. F. V. , 1990: Bryophyte interactions with other plants. Botanical Journal of the Linnean Society, 104: 79–98.
- Elumeeva, T. G. , Soudzilovskaia, N. A. , During, H. J. , and Cornelissen, J. H. C. , 2011: The importance of colony structure versus shoot morphology for the water balance of 22 subarctic bryophyte species. Journal of Vegetation Science, 22: 152–164.
- Engstrom, R. , and Hope, A. , 2011: Parameter sensitivity of the Arctic Biome—BGC model for estimating evapotranspiration in the Arctic Coastal Plain. Arctic, Antarctic, and Alpine Research, 43: 380–388.
- Engstrom, R. , Hope, A. , Kwon, H. , Harazono, Y. , Mano, M. , and Oechel, W. , 2006: Modeling evapotranspiration in Arctic Coastal Plain ecosystems using a modified BIOME—BGC model. Journal of Geophysical Research, 111: G02021, 20 pp., http://dx.doi.org/ 10.1029/2005JG000102.
- Esseen, P.-A. , Ehnström, B. , Ericson, L. , and Sjöberg, K. , 1997: Boreal forests. Ecological Bulletins, 46: 16–47, http://www.jstor.org/stable/ 20113207 .
- Euskirchen, E. S. , McGuire, A. D. , Rupp, T. S. , Chapin, F. S., III , and Walsh, J. E. , 2009: Projected changes in atmospheric heating due to changes in fire disturbance and the snow season in the western Arctic, 2003–2100. Journal of Geophysical Research, 114: G04022, 15 pp., http://dx.doi.org/10.1029/2009JG001095.
- Fox, A. M. , Huntley, B. , Lloyd, C. R. , Williams, M. , and Baxter, R. , 2008: Net ecosystem exchange over heterogeneous Arctic tundra: scaling between chamber and eddy covariance measurements. Journal of Geophysical Research, 22: GB2027, 15 pp., http://dx.doi.org/10.1029/2007GB003027.
- Gordon, C. , Wynn, J. M. , and Woodin, S. J. , 2001: Impacts of increased nitrogen supply on high Arctic heath: the importance of bryophytes and phosphorus availability. New Phytologist, 149: 461–471.
- Gornall, J. L. , Jónsdóttir, I. S. , Woodin, S. J. , and Van der Wal, R. , 2007: Arctic mosses govern below-ground environment and ecosystem processes. Oecologia, 153: 931–941.
- Henry, G. H. R. , and Gunn, A. , 1991: Recovery of tundra vegetation after overgrazing by caribou in arctic Canada. Arctic, 44: 38–42.
- Hollinger, D. Y. , Ollinger, S. V. , Richardson, A. D. , Meyers, T. P. , Dail, D. B. , Martin, M. E. , Scott, N. A. , Arkebauer, T. J. , Baldocchi, D. D. , Clark, K. L. , Curtis, P. S. , Davis, K. J. , Desai, A. R. , Dragoni, D. , Goulden, M. L. , Gu, L. , Katul, G. G. , Pallardy, S. G. , Paw U, K. T. , Schmid, H. P. , Stoy, P. C. , Suyker, A. E. , and Verma, S. B. , 2010: Albedo estimates for land surface models and support for a new paradigm based on foliage nitrogen concentration. Global Change Biology, 16: 696–710.
- Hudson, J. M. G. , and Henry, G. H. R. , 2010: High Arctic plant community resists 15 years of experimental warming. Journal of Ecology, 98(5): 1035–1041, http://dx.doi.org/10.1111/j.1365-2745.2010.01690.x .
- Jia, G. J. , Epstein, H. E. , and Walker, D. A. , 2003: Greening of arctic Alaska, 1981–2001. Geophysical Research Letters, 30: 4 pp., http://dx.doi.org/10.1029/2003GL018268.
- Joly, K. , Jandt, R. R. , Meyers, C. R. , and Cole, M. J. , 2007: Changes in vegetative cover on Western Arctic Herd winter range from 1981–2005: potential effects of grazing and climate change. Rangifer, Special Issue 17: 199–207.
- Joly, K. , Jandt, R. R. , and Klein, D. R. , 2009: Decrease of lichens in Arctic ecosystems: the role of wildfire, caribou, reindeer, competition and climate in north-western Alaska. Polar Research, 28: 433–442.
- Juang, J.-Y. , Katul, G. G. , Siqueira, M. B. S. , Stoy, P. C. , and Novick, K. A. , 2007: Separating the effects of albedo from eco-physiological changes on surface temperature along a successional chronosequence in the southeastern US. Geophysical Research Letters, 34: L21408, http://dx.doi.org/10.1029/2007GL031296 .
- Kayhko, J. , and Pellikka, P. , 1994: Remote sensing of the impact of reindeer grazing on vegetation in northern Fennoscandia using SPOT XS data. Polar Research, 13: 115–124.
- Kershaw, K. A. , 1978: The role of lichens in boreal tundra transition areas. The Bryologist, 81: 294–306.
- Kershaw, K. A. , 1985: Physiological Ecology of Lichens : Cambridge, New York: Cambridge University Press.
- Kim, J. , and Verma, S. B. , 1996: Surface exchange of water vapour between an open Sphagnum, fen and the atmosphere. Boundary-Layer Meteorology, 79: 243–264.
- Lange, O. L. , 1962: Die Photosynthese der Flechten bei tiefen Temperaturen und nach Frostperioden. Berichte der Deutchen Botanischen Gesellschaft, 75: 441–454.
- Larcher, W. , 1995: Phsiological Plant Ecology. 3rd edition. Berlin: Springer, 506 pp.
- Liljedahl, A. K. , Hinzman, L. D. , Harazono, Y. , Zona, D. , Tweedie, C. E. , Hollister, R. D. , Engstrom, R. , and Oechel, W. C. , 2011: Nonlinear controls on evapotranspiration in arctic coastal wetlands. Biogeosciences, 8: 3375–3389.
- Nichols, D. S. , and Brown, J. M. , 1980: Evaporation from a sphagnum moss surface. Journal of Hydrology, 48: 289–302.
- Nilsson, M.-C , and Wardle, D. A. , 2005: Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Frontiers in Ecology and the Environment, 3: 421–428.
- Norman, J. M. , Kustas, W. P. , and Humes, K. S. , 1995: Source approach for estimating soil and vegetation energy fluxes in observations of directional radiometric surface temperature. Agricultural and Forest Meteorology, 77: 263–293.
- O'Donnell, J. A. , Romanovsky, V. E. , Harden, J. W. , and McGuire, A. D. , 2009: The effect of moisture content on the thermal conductivity of moss and organic soil horizons from black spruce ecosystems in interior Alaska. Soil Science, 174: 646–651.
- Oechel, W. C. , and van Cleve, K. , 1986: The role of bryophytes in nutrient cycling in the taiga. In van Cleve, K. , Chapin, F. S., III , Flanagan, P. W. , Viereck, L. A. , and Dyrness, C. T. (eds.), Forest Ecosystems in the Alaskan Taiga: a Synthesis of Structure and Function. New York: Springer-Verlag, 121–137.
- Petzold, D.E. , and Rencz, A. N. , 1975: The albedo of selected subarctic surfaces. Arctic and Alpine Research, 7: 393–398.
- Rees, W. G. , 1993: Infrared emissivities of Arctic land cover types. International Journal of Remote Sensing, 14: 1013–1017.
- Schaaf, C. B. , Gao, F. , Strahler, A. H. , Lucht, W. , Li, X. , Tsang, T. , Strugnell, N. C. , Zhang, X. , Jin, Y. , Muller, J.-P. , Lewis, P. , Barnsley, M. , Hobson, P. , Disney, M. , Roberts, G. , Dunderdale, M. , Doll, C. , d'Entremont, R. , Hu, B. , Liang, S. , Privette, J. L. , and Roy, D. P. , 2002: First operational BRDF, albedo and nadir reflectance products from MODIS. Remote Sensing of Environment, 83: 135–148.
- Schuur, E. A. G. , Bockheim, J. , Canadell, J. G. , Euskirchen, E. S. , Field, C. B. , Goryachkin, S. V. , Hagemann, S. , Kuhry, P. , Lafleur, P. M. , Lee, H. , Mazhitova, G. , Nelson, F. E. , Rinke, A. , Romanovsky, V. E. , Shiklomanov, N. , Tarnocai, C. , Venevsky, S. , Vogel, J. G. , and Zimov, S. A. , 2008: Vulnerability of permafrost carbon to climate change: implications for the global carbon cycle. BioScience, 58: 701–714.
- Shaver, G. R. , Street, L. E. , Rastetter, E. B. , van Wijk, M. T. , and Williams, M. , 2007: Functional convergence in regulation of net CO2 flux in heterogeneous tundra landscapes in Alaska and Sweden. Journal of Ecology, 95: 802–817.
- Skre, O. , and Oechel, W. C. , 1981: Moss functioning in different taiga ecosystems in interior Alaska I. Seasonal, phenotypic, and drought effects on photosynthesis and response patterns. Oecologia, 48: 50–59.
- Skre, O. , Oechel, W. C. , and Miller, P. M. , 1983: Moss leaf water content and solar radiation at the moss surface in a mature black spruce forest in central Alaska. Canadian Journal of Forest Research—Revue Canadienne De Recherche Forestiere, 13: 860–868.
- Spadavecchia, L. , Williams, M. , Bell, R. , Stoy, P. C. , Huntley, B. , and van Wijk, M. T. , 2008: Topographic controls on the leaf area index of a Fennoscandian tundra ecosystem. Journal of Ecology, 96: 1238–1251.
- Street, L. E. , 2011: Carbon Dynamics in Arctic Vegetation. PhD thesis. School of GeoSciences, University of Edinburgh, Edinburgh, U.K., 236 pp.
- Street, L. E. , Shaver, G. R. , Williams, M. , and van Wijk, M. T. , 2007: What is the relationship between changes in canopy leaf area and changes in photosynthetic CO2 flux in arctic ecosystems? Journal of Ecology, 95: 139–150.
- Street, L. E. , Stoy, P. C. , Sommerkorn, M. , Fletcher, B. J. , Sloan, V. , Hill, T. C. , and Williams, M. , 2011a: Seasonal bryophyte productivity in the sub-arctic: a comparison with vascular plants. Functional Ecology, 26: 365–378, http://dx.doi.ord/10.1111/j.13652435.2011.01954.x .
- Street, L. E. , Subke, J.-A. , Sommerkorn, M. , Heinemeyer, A. , and Williams, M. , 2011b: Turnover of recently assimilated carbon in arctic bryophytes. Oecologia, 167: 325–337.
- Tarnocai, C. , Canadell, J. G. , Schuur, E. A. G. , Kuhry, P. , Mazhitova, G. , and Zimov, S. , 2009: Soil organic carbon pools in the northern circumpolar permafrost region. Global Biogeochemical Cycles, 23: GB2023, 11 pp., http://dx.doi.org/10.1029/2008GB003327 .
- van Wijk, M. T. , Clemmensen, K. E. , Shaver, G. R. , Williams, M. , Callaghan, T. V. , Chapin, F. S. I. , Cornelissen, J. H. C. , Gough, L. , Hobbie, S. E. , Jonasson, S. , Lee, J. A. , Michelsen, A. , Press, M. C. , Richardson, S. J. , and Reuth, H. , 2004: Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, northern Sweden: generalizations and differences in ecosystem and plant type responses to global change. Global Change Biology, 10: 105–123.
- Vogelmann, J. E. , and Moss, D. M. , 1993: Spectral reflectance measurements in the genus Sphagnum. Remote Sensing of Environment, 45: 273–279.
- Walker, D. A. , Raynolds, M. K. Daniels, F. J. A. , Einarsson, E. , Elvebakk, A. , Gould, W. A. , Katenin, A. E. , Kholod, S. S. , Markon, C. J. , Melnikov, E. S. , Moskalenko, N. G. , Talbot, S. S. , Yurtsev, B. A. , and Team, C. , 2005: The circumpolar arctic vegetation map. Journal of Vegetation Science, 16: 267–282.
- Walker, M. D. , Wahren, C. H. , Hollister, R. D. , Henry, G. H. R. , Ahlquist, L. E. , Alatalo, J. M. , Bret-Harte, M. S. , Calef, M. P. , Callaghan, T. V. , Carroll, A. B. , Epstein, H. E. , Jonsdottir, I. S. , Klein, J. A. , Magnuʃsson, B. , Molau, U. , Oberbauer, S. F. , Rewa, S. P. , Robinson, C. H. , Shaver, G. R. , Suding, K. N. , Thompson, C. C. , Tolvanen, A. , Totland, O. , Turner, P. L. , Tweedie, C. E. , Webber, P. J. , and Wookey, P. A. , 2006: Plant community responses to experimental warming across the tundra biome. Proceedings of the National Academy of Sciences of the United States of America, 103: 1342–1346.
- Williams, M. , Bell, R. , Spadavecchia, L. , Street, L. E. , and van Wijk, M. T. , 2008: Upscaling leaf area index in an Arctic landscape through multi-scale observations. Global Change Biology, 14: 1517–1530.
- Wookey, P. A. , Aerts, R. , Bardgett, R. D. , Baptist, F. , Brathen, K. A. , Cornelissen, J. H. C. , Gough, L. , Hartley, I. P. , Hopkins, D. W. , Lavorel, S. , and Shaver, G. R. , 2009: Ecosystem feedbacks and cascade processes: understanding their role in the responses of Arctic and alpine ecosystems to environmental change. Global Change Biology , 15: 1153–1172.
- Zimov, S. A. , Schuur, E. A. G , and Chapin, F. S., III, 2005: Permafrost and the global carbon budget. Science, 312: 1612–1613.