
Abstract
Background
Bisphosphonates, including zoledronate, target osteoclasts and are widely used in the treatment of osteoporosis and other bone resorption diseases, despite side effects that include damaging the stomach epithelium. Beneficial and adverse effects on other organ systems, including the cardiovascular system, have also been described and could impact on the use of bisphosphonates as therapeutic agents. Vascular smooth muscle cells (VSMCs) are major constituents of the normal vascular wall and have a key role in intimal thickening and atherosclerosis, in part by secreting MMPs that remodel the extracellular matrix and cleave cell surface proteins or secreted mediators. In this study, we investigated the effects of zoledronate on MMP expression.
Methods
Rat VSMCs were stimulated by PDGF (50 ng/mL) plus TNF-α (10 ng/mL) or left unstimulated for a further 24 hours in serum-free medium. In other series of experiments, cells were pre-treated either with SC-514 (50 μM) or with apocynin (20 nM) for 2 hours, then zoledronate (100 μM) was added into 2% fetal calf serum containing medium for 24 hours.
Results and discussion
Using isolated rat VSMCs in culture, zoledronate (100 μM) increased MMP-9 and -13 mRNA expressions but inhibited MMP-2 expression. MMP-9 and MMP-13 up-regulation was shown to depend on the NF-κB pathway; and this was activated by zoledronate. Furthermore, zoledronate elevated the levels of reactive oxygen species detected by either dichlorofluorescein in isolated VSMCs or lucigenin enhanced chemiluminescence in rat aortic rings in vitro. Apocynin, an inhibitor of NADPH oxidase, reversed NF-κB activation and MMP-9 and MMP-13 up-regulation by zoledronate.
Conclusion
We conclude that zoledronate increases MMP-9 and MMP-13 expressions in rat VSMCs dependent upon stimulation of the NF-κB pathway by reactive oxygen species. Effects on MMP expression may contribute to the pharmacologic profile of bisphosphonates.
Introduction
Bisphosphonates are widely used clinically for the treatment of osteoporosis and bone metastasis. Although they primarily target osteoclasts, adverse and beneficial effects on other organ systems have been described. Destruction of the gastric epithelium by high concentrations of bisphosphonates is a recognized off-target effect. On the contrary, beneficial effects on atherosclerosis have also been described,Citation1–Citation3 although the precise mechanisms involved have not been elucidated.
Migration and proliferation of vascular smooth muscle cells (VSMCs) play key roles in the development of vascular proliferative diseases such as atherosclerosis and restenosis.Citation4 Moreover, previous studies showed that bisphosphonates inhibit adhesion of VSMCs to extracellular matrix components in tissue culture, as well as their proliferation and migration.Citation5,Citation6 The end result is decreased neointimal hyperplasia both in preclinical and clinical investigations.Citation7–Citation9 MMPs play a variety of roles in normal vascular physiology and pathology. For example, increased expression of MMP-13 (especially in rodents) can degrade fibrillar type I and III collagen, thereby increasing atherosclerotic plaque’s vulnerability to rupture.Citation10 Conversely, MMP-2 and MMP-9 stimulate migration and proliferation of VSMCs by degrading basement components including type IV collagen. This contributes to restenosis but may prevent plaque rupture thanks to formation of a more stable plaque cap.Citation11
Bisphosphonates have been reported to inhibit expression and activity of some MMPs in different steps of cancer progression.Citation12–Citation14 However, the effects of bisphosphonates on MMPs implicated in the pathogenesis of atherosclerosis and restenosis have not been elucidated. In this study, we therefore investigated the effects of zoledronate, which is a highly potent bisphosphonate, on MMP-2, MMP-9, and MMP-13 expressions in rat VSMCs.
Materials and methods
Cell culture
Primary rat cells
Male Sprague Dawley rats were killed by cervical dislocation in accordance with the Directive 2010/63/EU of the European Parliament. Approval was granted by the University of Bristol ethical review board. VSMCs (four separate preparations from different rats) were obtained by the explant method from thoracic aortas and were cultured in Dulbecco’s Modified Eagle’s Medium (Lonza, Basel, Switzerland) containing 10% fetal calf serum (FCS), penicillin (100 mg/mL), streptomycin (100 U/mL), and L-glutamine (2 mM). VSMCs were used between passage 6 and 8 in the experiments, which is within the limits (passage 3–8) of previous studies.Citation15 Reduced serum (2%) or serum-free conditions were used in some incubations to avoid interfering with the zymographic assays, and were well tolerated by VSMCs.Citation16
Treatment of the cells
Zoledronate (Novartis International AG, Basel, Switzerland) was obtained as a 2.94 mM sterile, neutral aqueous solution for intravenous infusion containing 4 mg of zoledronate, 220 mg mannitol, and 24 mg sodium citrate in 5 mL. Cells were seeded into 12-well plates. When they became confluent, they were pre-treated with zoledronate to a final concentration of 100 μM zoledronate or kept as untreated in 2% FCS containing medium for 24 hours. Given the high dilution and innocuous nature of the vehicle, untreated cells were used as a control. Cells were stimulated by PDGF (50 ng/mL) plus TNF-α (10 ng/mL) or left unstimulated for a further 24 hours in serum-free medium. In other series of experiments, cells were pre-treated either with SC-514 (Cayman Chemicals MA, USA), (50 μM)Citation17 or with apocynin (20 nM) for 2 hours, then zoledronate (100 μM) was added into 2% FCS containing medium for 24 hours.
Real-time polymerase chain reaction (PCR) analysis
Total RNA were extracted from VSMCs using the total RNA extraction kit (Ambion, Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instructions. Extracted RNA was quantified using Nanodrop™ 1000 Spectrophotometer (Thermo Fisher Scientific) and reverse transcribed using a Roche Reverse Transcription Kit (Hoffman-La Roche Ltd., Basel, Switzerland) (100 ng RNA per reaction). A genomic DNA removal step was included, and the resulting cDNA was diluted in 10 mM Tris-HCl, pH 8. Real-time quantitative PCR was performed in Illumina Eco Real-Time PCR with Roche SYBR Green Master Mix and primers for MMP-2, MMP-9, MMP-13, IκB, and 36B4, which is a housekeeping gene. Primers are listed in . To confirm amplification specificity, PCR products from each primer pair were subjected to agarose gel electrophoresis. Relative mRNA expression levels were calculated by using the Delta-Delta Ct method using 36B4 as a normalization control.
Table 1 Primer sequences used for real-time polymerase chain reaction
Western blotting and gelatin zymography
Conditioned media of the cells were collected and concentrated using centrifugal filter units (Merck Millipore, Billerica, MA, USA). Cells were lysed in sodium dodecyl sulfate (SDS) buffer (2% SDS [w/v], 16% glycerol [v/v], and 50 mM Tris [pH 6.8]). Protein concentration in lysates was measured using BCA Protein Assay Kit (Thermo Fisher Scientific) and was used to normalize loading of gels.
MMP-9 activity in conditioned medium was determined by gelatin zymography as described previously.Citation17 Briefly, 7.5% polyacrylamide gels containing 2 mg/mL of gelatin were subjected to electrophoresis. Following electrophoresis, SDS was removed by washing in 2.5% Triton X-100. Gels then were incubated at 37°C for 48 hours in incubation buffer (50 mM Tris-HCl, pH 8.0, 50 mM NaCl, 10 mM CaCl2, and 0.05% Triton X-100). Ethylenediaminetetraacetic acid (EDTA) inhibits MMP activity by chelating Zn2+ on enzymes’ catalytic domain. In order to verify if the detected bands were MMP-2 and -9, another gel was incubated in calcium-free incubation solution containing 10 mM EDTA. After the incubation period, gels were stained in 0.2% Coomassie Brilliant Blue. Images of the gels were captured with a scanner. Gelatinase activity was detected as clear bands on dark backgrounds.
For Western blotting, equal total protein concentrations (see above) were loaded on 8%–10% polyacrylamide gels and transferred onto polyvinylidene difluoride membranes as described previously.Citation18 These were blocked for an hour in 5% (w/v) skimmed milk powder and incubated overnight at 4°C with anti-MMP-13 (catalog no ab39012; Abcam, Cambridge, UK) (1:2,000 in 5% BSA:Tris-buffered saline). Bound primary antibodies were detected by horseradish peroxidase-conjugated antibodies (Sigma-Aldrich Co., St Louis, MO, USA; catalog no A6154) (1:5,000 in 5% BSA:Tris-buffered saline) and enhanced chemiluminescence (Amersham International, Buckinghamshire, UK).
Detection of reactive oxygen species (ROS) in rat thoracic aortic rings
This study was approved by the Local Ethics Committee of Dokuz Eylul University School of Medicine. All animals received care in compliance with the principles of laboratory animal care formulated by the National Society for Medical Research and with the Guide for the Care and Use of Laboratory Animals prepared by the Institute of Laboratory Animal Resources.
Levels of superoxide anion and other ROS in aortic rings were measured as previously described.Citation19 In this study, Wistar rats were euthanized by decapitation under ketamine/xylazine anesthesia. The thoracic aorta segment was dissected. Following careful removal of loose connective tissue, thoracic aorta was cut into 2 mm rings. The rings were then transferred into Dulbecco’s Modified Eagle’s Medium/F12 (Gibco, Thermo Fisher Scientific) cell culture medium containing 2% FCS immediately. The rings were incubated in cell culture medium in the presence or absence of 100 μM zoledronate for 24 hours in a humidified atmosphere containing 5% CO2 at 37°C. Subsequently, aortic rings were transferred to solid white well plates containing 200 mL of HEPES-buffered Krebs-Henseleit solution (pH 7.4). After addition of chemiluminescence enhancers, lucigenin or luminol (final concentration of 5 μmol/L for either), ROS were quantified using a multi-plate reader (Victor III-1420; PerkinElmer Inc., Waltham, MA, USA). Counts were obtained every 10 seconds and normalized per mg of wet tissue weight. Results were expressed as the area under curve for a counting period of 5 minutes of relative light units (rlu)/wet tissue (mg).
Statistical analysis
Statistical analyses were performed using the Prism software v.4 (GraphPad Software, Inc., La Jolla, CA, USA). Multiple comparisons were subjected to one-way analysis of variance followed by post hoc Bonferroni correction. Paired Student’s t-tests were performed to compare means of lucigenin/luminol measurements. When necessary, data were normalized by log transformation. Normalized data were expressed as mean ± standard error of the mean P≤0.05 was considered statistically significant.
Results
Effects of zoledronate on expression of MMPs with and without induction by PDGF and TNF-α
Because of their diverse roles and regulation patterns, we studied constitutively expressed gelatinase, MMP-2, together with another gelatinase, MMP-9, and a collagenase, MMP-13, that could be induced by a combination of PDGF and TNF-α.
Gelatinase A/MMP-2
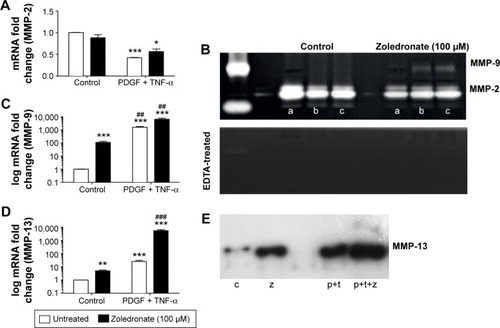
MMP-2 was constitutively expressed in VSMCs (), in agreement with previous studies.Citation16 Zoledronate pre-treatment did not change constitutive MMP-2 mRNA levels (). PDGF plus TNF-α treatment significantly decreased MMP-2 mRNA levels to a similar extent in untreated and zoledronate pre-treated cells (). This was confirmed by zymography in which MMP-2 appeared as a major 72 kDa band () that disappeared on incubation in the presence of EDTA (lower panel).
Figure 1 Effects of PDGF plus TNF-α and zoledronate (48 hours) on MMP-2, -9, and -13.
Abbreviation: EDTA, ethylenediaminetetraacetic acid.
Gelatinase B/MMP-9
MMP-9 was expressed at barely detectable levels in quiescent VSMCs () also in agreement with previous work.Citation16 However, zoledronate pre-treatment significantly increased MMP-9 mRNA levels (). PDGF plus TNF-α induced even higher MMP-9 mRNA levels and this effect was enhanced by zoledronate pre-treatment (). Consistent with this, only after PDGF plus TNF-α induction of zoledronate pre-treated cells was MMP-9 protein detectable by zymography as a 92 kDa band () that disappeared on incubation in the presence of EDTA (lower panel).
Collagenase 3/MMP-13
The effects of zoledronate on MMP-13 expression were similar to those on MMP-9. Zoledronate pre-treatment significantly increased MMP-13 mRNA and protein levels in control cells (). PDGF plus TNF-α induction increased MMP-13 mRNA and protein levels even further, and these were additively increased in zoledronate treated cells ().
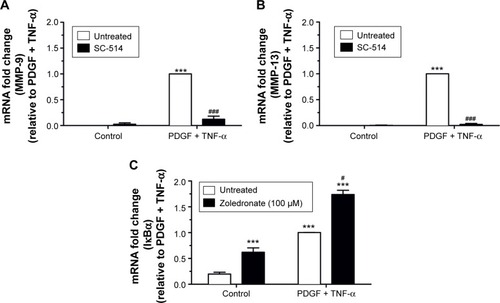
Role of NF-κB in MMP-9 and MMP-13 induction
The NF-κB pathway was previously shown to play an essential role in up-regulation of MMP-1, MMP-3, and MMP-9 in rabbit and human VSMCs using gene transfer of a dominant negative form of the inhibitor of κB, IκBα.Citation16 We also showedCitation17 that induction of several MMPs in human monocytes can be reversed by using SC-514, which is a selective inhibitor of IκB kinase 2.Citation20 We therefore used SC-514 to investigate whether NF-κB also mediates MMP-9 and MMP-13 up-regulation in rat VSMCs. SC-514 had no effect on the barely detectable levels of MMP-9 () or MMP-13 () mRNA in dimethyl sulfoxide treated cells. However, SC-514 strongly inhibited induction of MMP-9 and MMP-13 by PDGF plus TNF-α, thereby demonstrating the importance of NF-κB in up-regulation of both these MMPs. To investigate whether zoledronate activated the NF-κB pathway in VSMCs, we measured expression of IκBα mRNA because its promoter is a direct target of NF-κB activation in a process designed to cause feedback inhibition.Citation21 Detectable levels of IκBα mRNA were present in quiescent VSMCs () but, similarly to MMP-9 and MMP-13, these were significantly augmented by pretreatment with zoledronate (). PDGF plus TNF-α increased IκBα mRNA levels even further and this was also enhanced by zoledronate pre-treatment ().
Figure 2 Evidence for involvement of the NF-κB pathway.
ROS production in zoledronate treated isolated VSMCs and aortas
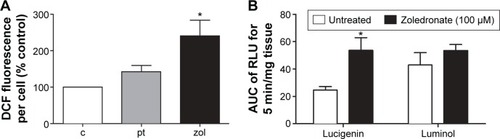
The NF-κB pathway is known to be sensitive to ROS production in VSMCs, independently of the effects of inflammatory mediators.Citation22 We therefore investigated whether ROS production in response to zoledronate might be responsible for activation of the NF-κB pathway, and hence MMP-9 and MMP-13 induction. Firstly, to confirm the activation of ROS, a dichlorofluorescein assay was performed to quantify cellular oxidative stress in isolated rat VSMCs. The cells were treated with zoledronate or PDGF plus TNF-α, with untreated cells as control. The results showed clearly that zoledronate treatment increased ROS levels, whereas no significant effect was observed with PDGF plus TNF-α (). For further confirmation, ROS levels were measured by using lucigenin or luminol in intact rat aortic tissue studied ex vivo. Incubation of rat aortic rings with zoledronate (100 μM) for 24 hours significantly elevated lucigenin enhanced chemiluminescence by approximately 2.5 times compared to control, untreated aortic rings (). The data for luminol enhanced chemiluminescence showed the same trend but was not significant (P=0.116, ).
Figure 3 Effects of zoledronate on ROS levels in rat VSMCs and aortic rings.
Abbreviations: ROS, reactive oxygen species; VSMCs, vascular smooth muscle cells; AUC, area under curve; RLU, relative light units.
Effects of NADPH oxidase inhibitor, apocynin, on induction of MMP-9, MMP-13, and IκB mRNAs
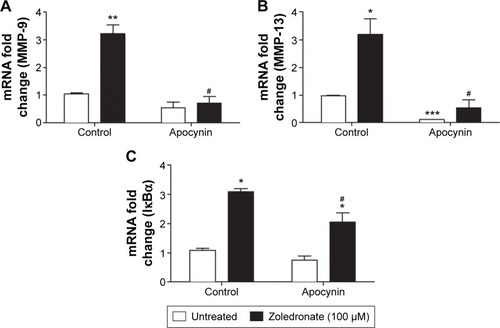
NADPH oxidase is a prominent source of ROS in VSMCs.Citation23 We therefore investigated the effects of apocynin, which is an inhibitor of NADPH oxidase. Apocynin reversed the elevation of MMP-9 mRNA levels in zoledronate pre-treated cells but had no significant effect on untreated cells (). Apocynin significantly decreased MMP-13 mRNA levels in untreated cells, and also normalized the high levels of MMP-13 mRNA expression in zoledronate pre-treated cells (). Furthermore, apocynin significantly inhibited NF-κB activation measured by the increase in mRNA levels of IκB-α in zoledronate pre-treated cells, but had no effect on untreated cells ().
Figure 4 Effects of apocynin with and without zoledronate on MMP-9, -13, and IκBα.
Discussion
In this study, we investigated the effects of zoledronate in primary rat VSMCs on expression of three abundant MMPs that are known to influence their migration, proliferation, and apoptosis. We chose MMP-2, MMP-9, and MMP-13 since they show diverse substrate specificity and response to induction by PDGF plus TNF-α. Our data, for the first time, demonstrated that zoledronate induced expression of MMP-9 and MMP-13 but not MMP-2. Then, we investigated the possible underlying mechanisms and revealed that zoledronate may have a role in ROS production and activation of the NF-κB pathway.
As mentioned in the Introduction, bisphosphonates have been reported to inhibit expression and activity of some MMPs during cancer progression.Citation12–Citation14 However, we found that zoledronate increased expressions of MMP-9 and MMP-13 in primary rat VSMCs, consonant with other published studies. For example, in a published study zoledronate administration before tooth extraction was stated to augment MMP-9 mRNA levels, protein expression of MMP-9, and enzymatic activity of MMP-9 during wound healing. The same study also reported that RANKL was at least partially responsible for the increased MMP-9 mRNA transcription.Citation24 Similarly, pamidronate, another nitrogen containing bisphosphonate similar to zoledronate, at concentrations of 30 μM to 300 μM, caused a 5- to 10-fold increase in MMP-9 mRNA levels in human monocytes.Citation25 Nitrogen containing bisphosphonate, alendronate markedly stimulated mRNA levels of MMP-13 in primary rat osteoblasts at concentrations higher than 10 μM.Citation26 While these results support our basic findings, none of the published studies identified the underlying mechanisms.
We showed here that NF-κB activity is essential for up-regulation of MMP-9 and MMP-13 in rat VSMCs using the competitive inhibitor of κB kinase 2, SC-514.Citation20 SC-514 can target other unrelated kinases PIM3, PIM1, DYRK1A, DYRK3, and Aurora B with similar affinity.Citation27 However, MMP-1, MMP-3, and MMP-9 transcription in rabbit and human VSMCsCitation16 was also inhibited by adenovirus-mediated gene transfer of dominant negative IκBα, which reinforces the specific role of NF-κB in MMP-9 induction. Similarly, NF-κB activation was shown to be essential for MMP-13 induction in chondrocytes.Citation28
We hypothesized, furthermore, that zoledronate can activate the NF-κB pathway. We found from preliminary experiments, that the effects of zoledronate required a 24-hour pre-incubation (results not shown). This implied that any activation of NF-κB would be slow and the method to detect it would need to be sensitive to prolonged activation. Acute activation is triggered by phosphorylation of IκBα which is then targeted for degradation by the 26S proteasome, allowing NF-κB dimers to translocate to the nucleus, usually within minutes, and activate transcription.Citation21 Subsequently, one of the primary targets for active NF-κB is the IκBα gene, which drives re-synthesis of the IκBα protein and feeds back on the activation cascade.Citation21 As a consequence, IκBα mRNA accumulates over an extended time period and we argued that this might provide a suitable measurement to investigate the effects of zoledronate. In fact, we found clear evidence that zoledronate pre-treatment increased IκBα mRNA and therefore slowly induced NF-κB activation. However, the findings from this study regarding NF-κB require further investigation, since the current study did not examine the protein expressions of NF-κB.
To answer the question regarding how zoledronate activated NF-κB, we set up additional experiment series in rat VSMCs. Our results from dichlorofluorescein experiments showed that zoledronate treatment increased ROS levels; and this finding was confirmed with an experiment on rat thoracic aortic rings by lucigenin measurements. Our evidences are consistent with previous experiments demonstrating bisphosphonate induced superoxide production in gastric and small intestinal epithelial cells.Citation29 In addition, zoledronate administration (100 μg/kg subcutaneously for 28 days) caused increased oxidative stress and decreased antioxidant levels in rabbit livers.Citation30 To confirm the role of oxidative stress in NF-κB activation and up-regulation of MMP-9 and MMP-13, we showed that apocynin, an inhibitor of NADPH oxidase, significantly inhibited these effects of zoledronate in VSMCs.
Our results provide new understanding regarding the pharmacological effects of bisphosphonates in the cardiovascular system. They are of particular relevance to pathologies, such as restenosis and vein graft intimal thickening, where there is a predominant role of VSMCs.Citation4 Other sources of MMPs, such as macrophages, become relevant in the context of atherosclerosis,Citation4 and it would be interesting, in future studies, to investigate how they respond to zoledronate treatment. Moreover, the NF-κB pathway has an established role in the induction of several MMPs in diverse cell types including: human epithelial cells,Citation31 fibroblasts,Citation32 and macrophages.Citation17 It is therefore highly likely that the effects of zoledronate that we have identified here in rat VSMCs, apply more widely and could have a major impact on tissue remodeling and inflammatory activation caused by zoledronate. Further studies will be needed to address this interesting possibility.
In conclusion, our results conclusively demonstrate that zoledronate can increase expression of some MMPs in rat VSMCs. Furthermore, this is associated with stimulation of ROS production and consequent activation of the NF-κB pathway.
Acknowledgments
The authors would like to thank Dr Goksel Gokce, Ege University Faculty of Pharmacy and Dr Steve White, University of Bristol for valuable help and expertise on oxidative stress measurements. MZA would also like to thank Prof Levent Ustunes for kind help and encouragement. This study was supported by the British Heart Foundation grant CH95/001.
Disclosure
The authors have no conflicts of interest to disclose.
References
- YlitaloRSyvalaHTuohimaaPYlitaloPSuppression of immunoreactive macrophages in atheromatous lesions of rabbits by clodronatePharmacol Toxicol200290313914312071335
- Ylitalo-HeikkalaRBisphosphonates and atherosclerosis: why?Lupus2006151290317212001
- BevilacquaMDominguezLJRosiniSBarbagalloMBisphosphonates and atherosclerosis: why?Lupus200514977377916218486
- NewbyACZaltsmanABMolecular mechanisms in intimal hyperplasiaJ Pathol2000190330030910685064
- WuLZhuLShiWHZhangJMaDYuBZoledronate inhibits the proliferation, adhesion and migration of vascular smooth muscle cellsEur J Pharmacol2009602112413119000670
- AlbadawiHHauraniMJOkluRDifferential effect of zoledronic acid on human vascular smooth muscle cellsJ Surg Res2013182233934623164362
- CelilogluMAydinYBalciPKolamazTThe effect of alendronate sodium on carotid artery intima-media thickness and lipid profile in women with postmenopausal osteoporosisMenopause200916468969319240658
- GuzelogluMGulMReelBYurekliIAykutKHazanEThe effects of zoledronic acid on neointimal hyperplasia: a rabbit carotid anastomosis modelAnadolu Kardiyol Derg20111129310021285021
- SantosLLCavalcantiTBBandeiraFAVascular effects of bisphosphonates-a systematic reviewClin Med Insights Endocrinol Diabetes20125475423133318
- LibbyPMechanisms of acute coronary syndromes and their implications for therapyN Engl J Med2013368212004201323697515
- NewbyACMatrix metalloproteinases regulate migration, proliferation, and death of vascular smooth muscle cells by degrading matrix and non-matrix substratesCardiovasc Res200669361462416266693
- GreenJRBisphosphonates: preclinical reviewOncologist20049Suppl 431315459425
- MelaniCSangalettiSBarazzettaFMWerbZColomboMPAmino-biphosphonate-mediated MMP-9 inhibition breaks the tumor-bone marrow axis responsible for myeloid-derived suppressor cell expansion and macrophage infiltration in tumor stromaCancer Res20076723114381144618056472
- GreenJRGuentherAThe backbone of progress – preclinical studies and innovations with zoledronic acidCrit Rev Oncol Hematol201177Suppl 1S3S1221353178
- HewerRCSala-NewbyGBWuYJNewbyACBondMPKA and Epac synergistically inhibit smooth muscle cell proliferationJ Mol Cell Cardiol2011501879820971121
- BondMChaseAJBakerAHNewbyACInhibition of transcription factor NF-kappaB reduces matrix metalloproteinase-1, -3 and -9 production by vascular smooth muscle cellsCardiovasc Res200150355656511376631
- ReelBSala-NewbyGBHuangWCNewbyACDiverse patterns of cyclooxygenase-independent metalloproteinase gene regulation in human monocytesBr J Pharmacol201116381679169021371008
- TsaousiAWilliamsHLyonCAWnt4/β-catenin signaling induces VSMC Proliferation and is associated with intimal thickeningCirc Res2011108442743621193738
- GokceGArunMZErgothioneine produces relaxation in isolated rat aorta by inactivating superoxide anionEur Rev Med Pharmacol Sci201418213339334525487948
- KishoreNSommersCMathialaganSA selective IKK-2 inhibitor blocks NF-kappa B-dependent gene expression in interleukin-1 beta-stimulated synovial fibroblastsJ Biol Chem200327835328613287112813046
- BaldwinASJrThe NF-kappa B and I kappa B proteins: new discoveries and insightsAnnu Rev Immunol1996146496838717528
- RajagopalanSMengXPRamasamySHarrisonDGGalisZSReactive oxygen species produced by macrophage-derived foam cells regulate the activity of vascular matrix metalloproteinases in vitro. Implications for atherosclerotic plaque stabilityJ Clin Invest19969811257225798958220
- AmansoAMGriendlingKKDifferential roles of NADPH oxidases in vascular physiology and pathophysiologyFront Biosci (Schol Ed)201241044106422202108
- BasiDLHughesPJThumbigere-MathVMatrix metalloproteinase-9 expression in alveolar extraction sockets of Zoledronic acid-treated ratsJ Oral Maxillofac Surg201169112698270721752506
- VallealaHHanemaaijerRMandelinJRegulation of MMP-9 (gelatinase B) in activated human monocyte/macrophages by two different types of bisphosphonatesLife Sci200373192413242012954450
- VargheseSCanalisEAlendronate stimulates collagenase 3 expression in osteoblasts by posttranscriptional mechanismsJ Bone Miner Res200015122345235111127199
- BainJPlaterLElliottMThe selectivity of protein kinase inhibitors: a further updateBiochem J2007408329731517850214
- MengsholJAVincentiMPCoonCIBarchowskyABrinckerhoffCEInterleukin-1 induction of collagenase 3 (matrix metalloproteinase 13) gene expression in chondrocytes requires p38, c-Jun N-terminal kinase, and nuclear factor kappaB: differential regulation of collagenase 1 and collagenase 3Arthritis Rheum200043480181110765924
- NaganoYMatsuiHShimokawaOBisphosphonate-induced gastrointestinal mucosal injury is mediated by mitochondrial superoxide production and lipid peroxidationJ Clin Biochem Nutr201251319620323170047
- KarabulutABGulMKarabulutEKiranTROcakSGOtluOOxidant and antioxidant activity in rabbit livers treated with zoledronic acidTransplant Proc20104293820382221094863
- ChengCYHsiehHLHsiaoLDYangCMPI3-K/Akt/JNK/NF-κ is essential for MMP-9 expression and outgrowth in human limbal epithelial cells on intact amniotic membraneStem Cell Res20129192322459175
- BondMFabunmiRPBakerAHNewbyACSynergistic upregulation of metalloproteinase-9 by growth factors and inflammatory cytokines: an absolute requirement for transcription factor NF-kappa BFEBS Lett1998435129349755853