
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Background
It has been reported that formononetin (FMN), one of the main ingredients from famous traditional Chinese medicine “Huang-qi” (Astragalus membranaceus [Fisch] Bunge) for Qi-tonifying, exhibits the effects of immunomodulation and tumor growth inhibition via antiangiogenesis. Furthermore, A. membranaceus may alleviate the retinal neovascularization (NV) of diabetic retinopathy. However, the information of FMN on retinal NV is limited so far. In the present study, we investigated the effects of FMN on the hypoxia-induced retinal NV and the possible related mechanisms.
Materials and methods
The VEGF secretion model of acute retinal pigment epithelial-19 (ARPE-19) cells under chemical hypoxia was established by the exposure of cells to 150 μM CoCl2 and then cells were treated with 3-(5′-hydroxymethyl-2′-furyl)-1-benzylindazole (YC-1, a potent HIF-1α inhibitor, 1.0 μg/mL) or different concentrations of FMN (0.2 μg/mL, 1.0 μg/mL, and 5.0 μg/mL). The supernatants of cells were collected 48 hours later to measure the VEGF concentrations, following the manufacturer’s instruction. The mRNA expressions of VEGF, HIF-1α, PHD-2, and β-actin were analyzed by quantitative reverse transcription polymerase chain reaction, and the protein expressions of HIF-1α and PHD-2 were determined by Western blot analysis. Furthermore, the rats with retinopathy were treated by intraperitoneal administration of conbercept injection (1.0 mg/kg) or FMN (5.0 mg/kg and 10.0 mg/kg) in an 80% oxygen atmosphere. The retinal avascular areas were assessed through visualization of the retinal vasculature by adenosine diphosphatase staining and hematoxylin and eosin staining.
Results
FMN can indeed inhibit the VEGF secretion of ARPE-19 cells under hypoxia, downregulate the mRNA expression of VEGFA and PHD-2, and decrease the protein expression of VEGF, HIF-1α, and PHD-2 in vitro. Furthermore, FMN can prevent hypoxia-induced retinal NV in vivo.
Conclusion
FMN can ameliorate retinal NV via the HIF-1α/VEGF signaling pathway, and it may become a potential drug for the prevention and treatment of diabetic retinopathy.
Introduction
Astragalus membranaceus (Fisch) Bunge, known as Huang-qi in Chinese or Radix Astragali in Latin, is one of the most popular herbal medicines worldwide, and it has been widely used as a Qi-tonifying medicine in the People’s Republic of China, Mongolia, and Korea for a long time.Citation1,Citation2 Pharmacological studies have shown that A. membranaceus exhibits many beneficial effects, including immunomodulation,Citation3–Citation5 antihyperglycemic effects and improved insulin sensitivity,Citation6–Citation8 anti-inflammation effects,Citation9,Citation10 antioxidant effects,Citation11,Citation12 antiviral effects,Citation13,Citation14 hepatoprotection effects,Citation15,Citation16 antineoplastic effects, protection of cardiovascular function,Citation17 and so on. Meanwhile, phytochemical studies have displayed >100 compounds from A. membranaceus (root), such as flavonoids, polysaccharides, saponins, sucroses, amino acids, and phenolic acids.Citation1 Among them, formononetin (FMN; 7-hydroxy-4′-methoxyisoflavone), a flavonoid with neuroprotection,Citation18 anti-inflammation,Citation19 antiviral,Citation20 antiangiogenesis and tumor growth inhibition,Citation21 cardioprotection,Citation22 and other pharmacological effects, has been frequently used as the quality control marker of A. membranaceus and its preparations. Recently, it has been reported that some preparations mainly composed of A. membranaceus, such as Huangqi injection and Keluoxin capsule, could alleviate the retinal neovascularization (NV) of diabetic retinopathy (DR).Citation23,Citation24 A previous study has clearly demonstrated that FMN from A. membranaceus has an inhibitory effect on the tumor growth via antiangiogenesis,Citation21 but it is still unknown whether FMN can inhibit hypoxia-induced retinal NV in the pathophysiologic process of DR.
DR is a common microvascular complication of patients with diabetes mellitus.Citation25 Retinal NV can induce vitreous hemorrhage and tractional retinal detachment, resulting in visual deterioration.Citation26 Furthermore, increased vascular permeability leads to macular edema in patients with DR.Citation27 Therefore, DR becomes the leading cause of blindness in the adults. Importantly, VEGF plays a critical role in the retinal NV of DR, which stimulates the proliferation and migration of vascular endothelial cells and increases vascular permeability.Citation28 Hypoxia is one of the most potent triggers of VEGF expression, acting on the processes of DNA transcription, mRNA stabilization, and translation and release of VEGF,Citation29 which is centrally controlled by the HIF-1α, a transcription factor that regulates hypoxia-inducible genes, including VEGFA, and induces an angiogenic response.Citation30 Therefore, HIF-1α is increased to induce the expression of VEGF under hypoxia, resulting in increased vascular permeability and retinal NV. On the other hand, inhibition of HIF-1α can prevent the retinal NV in the condition of hypoxia.Citation31–Citation33 These studies indicate that HIF-1α/VEGF signaling pathway plays the key role in the retinal NV of DR. In the present study, we investigated the preventive effect of FMN on retinal NV from secretion of VEGF in the acute retinal pigment epithelial-19 (ARPE-19) cells induced by CoCl2 in vitro and NV of oxygen-induced retinopathy of a rat model in vivo.
Materials and methods
Reagents and antibodies
Conbercept injection (Lot: 20110610B) was provided by Chengdu Kanghong Pharmaceutical Group Co., Ltd. (Chengdu, Sichuan, People’s Republic of China). FMN (Lot: 20100311) was purchased from Jubang Biotechnology Co., Ltd. (Chengdu, Sichuan, People’s Republic of China) and dissolved in dimethyl sulfoxide (DMSO) to make a stock solution of 10.0 mg/mL. Human VEGF enzyme-linked immunosorbent assay (ELISA) kits (Lot: DVE00) were provided by R&D Systems Inc. (Minneapolis, MN, USA). Dulbecco’s Modified Eagle’s Medium (DMEM)/F12 medium, fetal bovine serum (FBS), and trypsin–ethylenediaminetetraacetic acid (EDTA) solution were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Tetramethylethylenediamine, DMSO, adenosine diphosphate (ADP), and ammonium sulfide were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). The 5-[1-(phenylmethyl)-1H-indazol-3-yl]-2-furanmethanol (YC-1), anti-HIF-1α antibody, and anti-PHD-2 antibody were obtained from Abcam (Cambridge, MA, USA). PrimeScript RT reagent kit with gDNA Eraser and SYBR Premix Ex Taq™ were purchased from Takara-Bio (Kusatsu, Shiga, Japan).
Cell culture
A human ARPE-19 cell line (Lot: 60279299) was purchased from the American Type Culture Collection (Manassas, VA, USA), and the cells were cultured in DMEM/F12 medium with 10% FBS, 100 units/mL penicillin, and 100 μg/mL streptomycin in an incubator at 37°C with 5% CO2. After 90% confluence, the cells were digested by 0.25% trypsin–0.02% EDTA for passage.
Determination of VEGF secretion by ARPE-19 cells under chemical hypoxia
According to the previous report from the literature,Citation28 ARPE-19 cells were seeded into 96-well plates at a density of 8×103 cells/well. After being cultured for 24 hours, the culture medium was replaced with fresh FBS-free DMEM/F12 medium. Then, the cells were treated with 1.0 μg/mL YC-1 and different concentrations of FMN (0.2 μg/mL, 1.0 μg/mL, and 5.0 μg/mL). The cells in the control group were treated with the culture medium containing the same concentration of DMSO as in the FMN solution. A total of 150 μM of CoCl2 was added to mimic hypoxic condition after 24 hours. The cells treated with DMSO-containing medium were used as control, and those treated with CoCl2 and YC-1 as positive controls. Conditioned medium was collected 48 hours later and centrifuged at 800× g for 5 minutes, and the supernatants were transferred to vials and stored at −80°C for further analysis. All experiments were performed in triplicate.
The concentrations of VEGF protein in the conditioned media were measured using the human VEGF Quantikine ELISA kits according to the manufacturer’s instruction. Absorbance values (450 nm) were recorded in triplicate with a microplate reader (SpectraMax M5; Molecular Devices LLC, Sunnyvale, CA, USA). The concentrations of VEGF were calculated from the standard curve. The sensitivity of the VEGF kits was 5.0 pg/mL.
RNA isolation and analysis of the mRNA expressions of VEGF, HIF-1α, and PHD-2
ARPE-19 cells were plated at a density of 1×105 cells/well in six-well plates, allowed to adhere overnight, and then treated with YC-1 (1.0 μg/mL) or different concentrations of FMN (0.2 μg/mL, 1.0 μg/mL, and 5.0 μg/mL). After being cultured for 24 hours, the culture media were withdrawn, and the cells were washed with cold phosphate-buffered saline before harvest. The cell pellets were collected for mRNA extraction after microcentrifuging at 800× g for 5 minutes at 4°C. Total RNA from the ARPE-19 cells was isolated with the TRIzol reagent. cDNA was synthesized with a cDNA synthesis kit according to the manufacturer’s protocol. The relative levels of each gene mRNA transcripts to β-actin were determined by quantitative reverse transcription polymerase chain reaction (qRT-PCR) using the SYBR premixed system and specific primers. The primer sequences for VEGFA, HIF-1α, PHD-2, and β-actin were as follows: homo VEGFA 5′-CGA AAC CAT GAA CTT TCT GC-3′ (forward) and 5′-CCT CAG TGG GCA CAC ACT CC-3′ (reverse), homo HIF-1α 5′-ACA AGT CAC CAC AGG ACA G-3′ (forward) and 5′-AGG GAG AAA ATC AAG TCG-3′ (reverse), homo PHD-2 5′-AAA CCA TTG GGC TGC TCA T-3′ (forward) and 5′-CGT ACA TAA CCC GTT CCA TTG-3′ (reverse), and homo β-actin 5′-AGC GGG AAA TCG TGC GTG AC-3′ (forward) and 5′-AGT TTC GTG GAT GCC ACA GGA C-3′ (reverse). qRT-PCR was performed by Opticon 3 continuous fluorescence detector (MJ Research, Inc., Waltham, MA, USA). The comparative cycle of threshold fluorescence (Ct) method was used, and the relative transcript amount of the target gene was normalized to that of β-actin using the
method.
Western blot analysis of the proteins of HIF-1α and PHD-2
ARPE-19 cells were seeded at a density of 2.0×105 cells/bottle in culture bottles, and the cells were treated with YC-1 (1.0 μg/mL) or different concentrations of FMN (0.2 μg/mL, 1.0 μg/mL, and 5.0 μg/mL). After being cultured for 24 hours, the culture media were withdrawn and the cells were washed with cold phosphate-buffered saline for harvest. The cell pellets were disrupted in cell radioimmunoprecipitation assay buffer (0.5% NP-40, 50 mM Tris-HCl, 120 mM NaCl, 1 mM EDTA, 0.1 mM Na3VO4, 1 mM NaF, 1 mM PMSF, and 1 μg/mL leupeptin, pH 7.5), and then the lysates were centrifuged at 12,000 rpm for 15 minutes at 4°C. The protein concentrations were determined using the BCA method, after an equal amount of protein (30 μg) was electrophoresed on 7.5% density of sodium dodecyl sulfate–acrylamide gels. Following electrophoresis, the proteins were transferred from the gel to a nitrocellulose membrane using an electron transfer system. Nonspecific binding was blocked with 5% skim milk in Tris-buffered saline with Tween 20 (TBST) buffer (5 mM Tris-HCl, 136 mM NaCl, and 0.1% Tween-20, pH 7.6) for 1 hour. The blots were incubated with antibodies against HIF-1α (1:2,000), PHD-2 (1:1,500), or β-actin (1:800) overnight at 4°C and were washed three times with 1× TBST. Then, the blots were incubated for 1 hour at room temperature with a 1:4,000 dilution of horseradish peroxidase-labeled antirabbit or antimouse IgG and washed three times with 1× TBST. The membranes were developed by incubation within the ECL Western blot detection reagents. The specific protein bands (the upper band of HIF-1α and the lower band of PHD-2) were visualized and analyzed with a ChemiDoc image analyzer (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Rat model of oxygen-induced retinopathy
All animal experiments were performed strictly in accordance with the university’s guidelines and were approved by the Committee on Use and Care of Animals of Sichuan University (Chengdu, Sichuan, People’s Republic of China). Ten pregnant (16–20 weeks old and body weight 300–350 g, for oxygen-induced retinopathy [OIR] model study) Sprague Dawley rats (SPF Grade, Certificate No SCXK2013-24) were purchased from the Experimental Animal Centre, Sichuan Provincial Academy of Medical Sciences in the People’s Republic of China (Chengdu, Sichuan, People’s Republic of China). OIR was induced in Sprague Dawley rat pups according to a protocol as previously described.Citation34 In brief, normalized litters of 50 Sprague-Dawley rat pups with their nursing mothers on postnatal day (PD) 7 were placed in an 80% oxygen atmosphere for 5 days. After being returned to normoxia condition for 5 days (on PD 12), the rat pups were intraperitoneally administered with Conbercept injection at 1.0 mg/kg or FMN at 5.0 mg/kg and 10.0 mg/kg, and the same volume of normal saline as control. Five days later (on PD 17), the rat pups were sacrificed (the pups in the control group were bred under normoxia conditions until PD 17), and the eyes were enucleated and fixed in fresh 4% paraformaldehyde for 2 hours. Eyecups were dissected, and the retinal flat mounts were created and stained with ADPase staining as described previously.Citation35 The retinal avascular areas were assessed through visualization of the retinal vasculature using infusion of ADPase staining, examined under fluorescent microscopy. In each whole mount, the total areas of preretinal neovascular areas were measured using Image-Pro Plus System and expressed as the percentage of the respective average in relation to total retinal areas. The retinas were also histologically examined. Serial sections of paraffin-embedded small pieces of retina (6 mm) were stained with hematoxylin and eosin. Images were taken under microscopy, and endothelial nuclei extended beyond the inner limiting membrane into the vitreous were manually counted in a blind manner.
Statistical analysis
All the data were reported as mean ± standard deviation. Statistical significance of the data was analyzed by one-way univariate analysis of variance for comparing data from more than two groups. A difference at P<0.05 was considered to be statistically significant (marked as *). The higher significance level was set at P<0.01 (marked as **). The data were analyzed among the CoCl2-treated groups as one factor and FMN-treated groups as another factor.
Results
FMN inhibits the secretion of VEGF in the ARPE-19 cells under hypoxia condition
FMN is an important compound in A. membranaceus for the protection of DR, and the structure is shown in . In order to study the effect of FMN on VEGF secretion under hypoxia, ELISA was applied to examine the concentrations of VEGF in the cultured medium of the ARPE-19 cells treated with 150 μM CoCl2 alone or in combination with FMN at 0.2 μg/mL, 1.0 μg/mL, or 5.0 μg/mL. As shown in , the level of VEGF in the culture supernatant was significantly increased after the cells were treated with 150 μM CoCl2 compared to the non-CoCl2 cells (P<0.01). However, YC-1 at 1.0 μg/mL and FMN at 0.2 μg/mL, 1.0 μg/mL, and 5.0 μg/mL markedly decreased the secretion of VEGF compared to the cells treated with CoCl2 under hypoxia condition (P<0.01). These results indicate that FMN inhibits the secretion of VEGF in the culture supernatant of the ARPE-19 cells.
Figure 1 Structure of 7-hydroxy-4′-methoxyisoflavone (FMN).
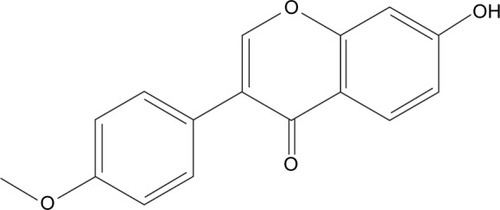
Figure 2 The effects of FMN on VEGF secretion in the ARPE-19 cells at 48 hours under hypoxic condition.
Abbreviations: FMN, formononetin; ARPE-19, acute retinal pigment epithelial-19; SD, standard deviation; DMSO, dimethyl sulfoxide.
Effects of FMN on the mRNA expressions of VEGFA, HIF-1α, and PHD-2 in the ARPE-19 cells
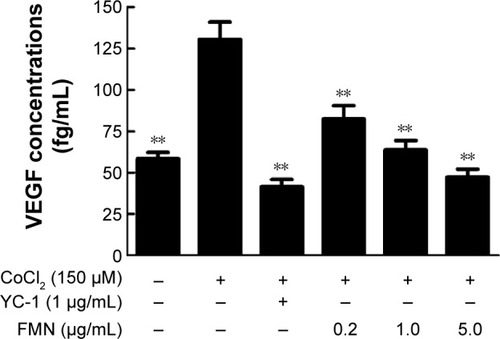
The expression of VEGFA can be induced by posttranscriptional mechanisms directed at the VEGFA mRNA or its relative regulator in the pathophysiological states.Citation36 We hypothesized that FMN may be involved in the mRNA expressions of VEGFA, HIF-1α, and PHD-2. For this purpose, we examined the mRNA expressions of VEGFA, HIF-1α, and PHD-2 in the ARPE-19 cells by qRT-PCR analysis. As shown in , the expression of VEGFA mRNA increased significantly in the ARPE-19 cells treated with 150 μM CoCl2 for 24 hours (P<0.01). FMN at 0.2–5.0 μg/mL significantly decreased the high expression of VEGFA mRNA induced by CoCl2 (P<0.01) in a dose-dependent manner (), indicating that FMN blocked the elevated expression of VEGFA mRNA induced by hypoxia. The data in show that the combination of 150 μM CoCl2 with YC-1 at 1.0 μg/mL or FMN at 0.2–5.0 μg/mL did not affect the mRNA expression of HIF-1α compared to CoCl2 alone. However, the data in show that the mRNA expression of PHD-2 was increased (P<0.01) under CoCl2-induced hypoxia-mimicking conditions and was significantly downregulated by YC-1 (1.0 μg/mL) or FMN (0.2–5.0 μg/mL) compared to CoCl2 alone (P<0.01). These results show that VEGF expression under hypoxia is associated with the mRNA expressions of VEGFA and PHD-2.
Figure 3 The effects of FMN on the mRNA expressions of VEGFA (A), HIF-1α (B), and PHD-2 (C) in the ARPE-19 cells at 24 hours under normoxia or hypoxic condition.
Abbreviations: FMN, formononetin; ARPE-19, acute retinal pigment epithelial-19; SD, standard deviation; DMSO, dimethyl sulfoxide.
Effects of FMN on the intracellular levels of HIF-1α and PHD-2 protein in the ARPE-19 cells
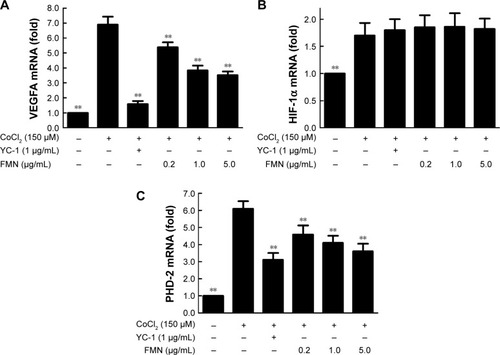
VEGFA is potently induced by hypoxia and regulated by HIF-1α and its degradation regulator PHD-2.Citation37 Here, we used the anti-HIF-1α and anti-PHD-2 antibodies to examine the protein expressions of HIF-1α and PHD-2 in the cultured ARPE-19 cells under hypoxia condition. Western blot analysis showed that the protein expressions of HIF-1α and PHD-2 were significantly upregulated in the ARPE-19 cells treated with 150 μM CoCl2 for 24 hours compared to the cells treated with the medium (P<0.01) and significantly downregulated by YC-1 (1.0 μg/mL) or FMN (0.2–5.0 μg/mL) compared to CoCl2 alone (P<0.01; ). These results indicate that the inhibition of FMN on VEGF secretion of the ARPE-19 cells under hypoxia condition is associated with the protein expressions of HIF-1α and PHD-2, suggesting a possible mechanism involved in hypoxia-induced retinal NV and related signaling pathways.
Figure 4 The effects of FMN on the protein expressions of HIF-1α (A) and PHD-2 (B) in the ARPE-19 cells at 24 hours under normoxia or hypoxic condition.
Abbreviations: FMN, formononetin; ARPE-19, acute retinal pigment epithelial-19; SD, standard deviation; DMSO, dimethyl sulfoxide.
FMN inhibits hypoxia-induced retinal NV in vivo
After we demonstrated the inhibitory effect of FMN on VEGF secretion of the ARPE-19 cells in vitro, next, we investigated the preventive effect of FMN on the hypoxia-induced retinal NV with the superficial vascular plexuses, using ADPase staining in a rat pup model. Rat pups were exposed to 80% oxygen from PD 7 to PD 12 to produce retinal vaso-obliteration and then returned to room air. This condition made the retina relatively hypoxic, resulting in the formation of retinal NV. On the PD 17, the preretinal neovascular area ratio was significantly increased in the OIR rats treated with vehicle (saline) compared to that of the normal rats treated with vehicle (P<0.01). However, Conbercept (1 mg/kg) or FMN (5 mg/kg and 10 mg/kg) by intraperitoneal injection markedly reversed the hypoxia-induced neovascular area ration and inhibited retinal NV (). Furthermore, Conbercept and FMN alleviated NV, vessel tortuosity, and dilated vessels in the OIR rats significantly ().
Figure 5 The effects of FMN on quantification of preretinal neovascular area ratio in the OIR rats.
Abbreviations: FMN, formononetin; OIR, oxygen-induced retinopathy; NV, neovascularization; SD, standard deviation; CBC, conbercept ophthalmic injection.
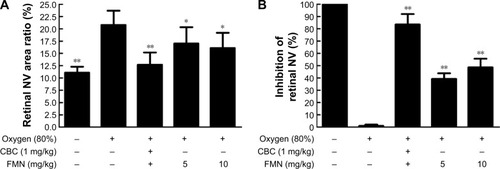
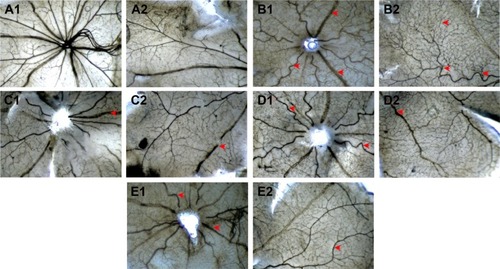
Figure 6 The effects of Conbercept and FMN on retinal NV of the OIR rats in the ADPase-stained retinal sections (40×).
Abbreviations: FMN, formononetin; NV, neovascularization; OIR, oxygen-induced retinopathy; ADP, adenosine diphosphate; ip, intraperitoneal.
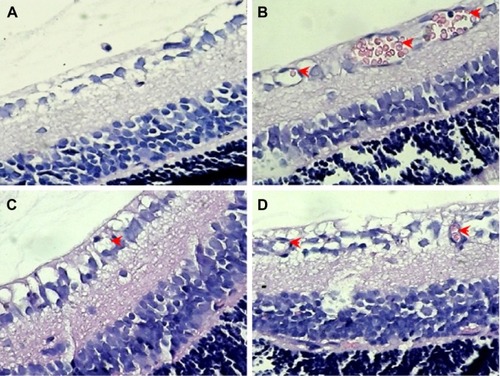
In order to further confirm the inhibitory effect of FMN on retinal NV, we counted the vascular cell nuclei extended beyond the internal limiting membrane in the retinal tissue sections of rats, with hematoxylin and eosin stain. Nuclei anterior to the internal limiting membrane were not found in the retinal tissue sections of the control rats (), but a lot of neovascular nuclei and vessels were found in the retinal tissue sections of the OIR rats (). Interestingly, the neovascular nuclei and vessels in the retinal tissue sections were remarkably reduced by Conbercept (1 mg/kg) or FMN (5 mg/kg and 10 mg/kg) treatment ().
Figure 7 The effects of FMN on retinal NV in H&E-stained retinal tissue sections (×400).
Abbreviations: FMN, formononetin; NV, neovascularization; H&E, hematoxylin and eosin; OIR, oxygen-induced retinopathy; ip, intraperitoneal.
Discussion
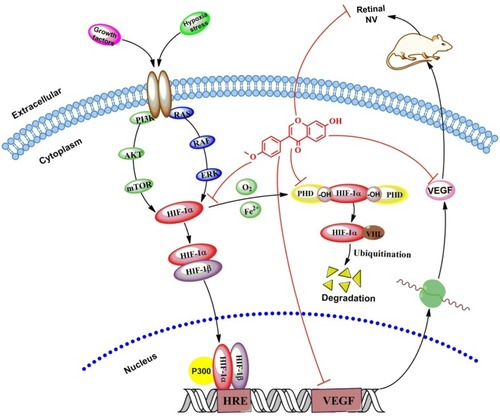
The present study aimed to investigate whether FMN can prevent hypoxia-induced retinal NV, and the role of HIF-1α/VEGF signaling pathway. We applied the VEGF secretion model of ARPE-19 cells induced by CoCl2 in vitro, and NV rat model of oxygen-induced retinopathy in vivo. Our results reveal that FMN can indeed inhibit VEGF secretion of the ARPE-19 cells under hypoxia condition, downregulate mRNA expressions of VEGFA and PHD-2, and decrease the protein expressions of VEGF, HIF-1α, and PHD-2 in vitro. Furthermore, the results also show that FMN can prevent hypoxia-induced retinal NV in vivo. The results show that FMN can alleviate hypoxia-induced retinal angiogenesis through suppression of the HIF-1α/VEGF signaling pathway (). Therefore, our findings suggest that FMN is effective against DR and may potentially be developed as a natural drug for the treatment of DR. However, the pharmacological effects on other animal models and the associated mechanism(s) against DR need to be further investigated.
Figure 8 FMN alleviates hypoxia-induced retinal angiogenesis through suppression of the HIF-1α/VEGF signaling pathway.
VEGF plays a key role in the retinal NV in the pathological process of DR, including angiogenesis, vasculogenesis, endothelial cell growth, and so on. Meanwhile, retinal pigment epithelium cells located between the photoreceptors and choriocapillaris (capillaries forming the inner vascular layer of the choroid) play an important role in maintaining retinal homeostasis and VEGF production within the area of blood–ocular barrier.Citation30,Citation38,Citation39 In the present study, we employed CoCl2, a widely used chemical compound, to mimic hypoxia condition in the cultured ARPE-19 cells to study the regulation of VEGF expression. Our data revealed that exposure of 150 mM CoCl2 to the cells significantly increased mRNA expression at 24 hours and markedly increased the VEGF secretion at 48 hours. The different response times with CoCl2 exposure may result because the mRNA transcription and translation and secretion of VEGF occurred at different times. Furthermore, our data demonstrate that FMN can simultaneously inhibit the mRNA transcription and protein expression of VEGF. This finding is consistent with the previous study of antiangiogenesis on the tumor with high expression of VEGF.Citation21 However, FMN also can contrarily promote angiogenesis through estrogen receptor or MAPK pathway in human pancreatic tumor cells with low expression of VEGF.Citation40 The possible reasons for the difference may be due to the diverse pathophysiologic foundation with different VEGF expression levels, or FMN as a natural selective estrogen receptor modulator and/or ligand for the GR and estrogen receptor (ER), to exhibit the angiogenic or antiangiogenic effect in the different pathophysiologic states as a bidirectionally regulative effect of A. membranaceus.Citation41
HIF is a heterodimeric transcriptional factor that is activated and stabilized under hypoxic condition. HIF promotes the expressions of gene products, including angiogenesis factors (eg, VEGF and TGF-β3), erythropoiesis factors (eg, EPO), cell survival, and proliferation factors (eg, IGF-2, ID2, and NOS).Citation37,Citation42–Citation46 Meanwhile, HIF-1 is composed of two subunits of HIF-1α and HIF-1β. HIF-1α is an oxygen-sensitive subunit, and its expression is induced under hypoxic conditions. In contrast, HIF-1β is constitutively expressed. HIF-1β is also known as aryl hydrocarbon nuclear translocator, because it needs to bind with HIF-1α and AhR, facilitating its translocation to the nucleus.Citation37 However, HIF-1α can be hydroxylated by HIF PHDs (especially PHD-2) in normoxia; the hydroxylated HIF-1α is multiubiquitinated and degraded in the proteasome. Conversely, the mRNA expression of PHD-2 is upregulated by hypoxia through an HIF-1α-dependent signaling pathway.Citation47 In addition, the PHD activity is inhibited under low oxygen tension.Citation48 This counter action could explain why the expression of PHD-2 is upregulated by CoCl2 and downregulated by FMN through inhibition of HIF-1α protein expression, as previously reported.Citation49 In the present study, we found the mRNA expressions of VEGF and PHD-2 and the protein expressions of VEGF, HIF-1α, and PHD-2 can be induced by CoCl2 under hypoxia conditions. Meanwhile, FMN can inhibit the increase induced by CoCl2 in a dose-dependent manner. FMN inhibited the secretion of VEGF via the HIF-1α/VEGF signaling pathway. In order to demonstrate the inhibitory effect of FMN on retinal NV in vivo, the rat model of oxygen-induced retinopathy was used to investigate the pharmacological effect of FMN on ischemic retinopathy induced by hyperoxia and followed by returning to normoxia as the literature reported.Citation34 Increased retinal NV was found in the OIR rats, and FMN significantly alleviated the NV, vessel tortuosity, and dilated vessels in the OIR rats, suggesting that FMN may be a potent inhibitor of retinal NV via the HIF-1α/VEGF signaling pathway.
Actually, HIF-1α undergoes quick degradation under normoxic condition and normally has a very short half-life (~5 minutes).Citation50 In contrast, in hypoxic conditions, several pathways have been shown to control HIF-1α stability and transcriptional activity via posttransnational modifications involving hydroxylation, acetylation, ubiquitination, and phosphorylation reactions, including hypoxic regulation pathway (pVHL dependent or pVHL independent),Citation51 growth factor signaling pathway,Citation52 Mdm2 pathway,Citation53 Hsp90 pathway, and so on.Citation54 Therefore, our further investigation into the molecular mechanisms associated with the effects of FMN on anti-retinal NV should include the study of upstream regulation, degradation of HIF-1α, and HRE transcription.
Conclusion
In the present study, we reveal that FMN plays an important role in the inhibition of VEGF secretion, through downregulating the mRNA expressions of VEGFA and PHD-2 and decreasing the protein expressions of VEGF, HIF-1α, and PHD-2 in the cultured ARPE-19 cells. Furthermore, we also demonstrate the inhibitory effect of FMN on retinal NV in a rat model of oxygen-induced retinopathy in vivo. These results may provide important insights into potential discovery and development of FMN as a novel drug for the treatment of DR clinically. However, more studies are needed to further investigate the pharmacological effects in other animal models and the possible associated mechanism(s) against DR.
Authors contributions
Jianming Wu, Xiao Ke, Xiaoping Gao, and Xiaofeng Hao conceived and designed the experiments; Jianming Wu, Wei Fu, Hongcheng Zhang, Wei Wang, Na Ma, and Manxi Zhao performed the experiments; Jianming Wu, Manxi Zhao, and Xiaoping Gao analyzed the data; Xiao Ke, Xiaofeng Hao, and Zhirong Zhang contributed new reagents and analysis tools; and Jianming Wu and Zhirong Zhang wrote the article. All authors contributed toward data analysis, drafting and critically revising the paper and agree to be accountable for all aspects of the work.
Acknowledgments
The authors thank Professor Shousong Cao (Southwest Medical University) for proof reading and editing of the manuscript. This work was supported by Technology Support Program of Science and Technology, Department of Sichuan Province, People’s Republic of China (2012SZ0038).
Disclosure
The authors report no conflicts of interest in this work.
References
- FuJWangZHuangLReview of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi)Phytother Res20142891275128325087616
- ZhangJXieXLiCFuPSystematic review of the renal protective effect of Astragalus membranaceus (root) on diabetic nephropathy in animal modelsJ Ethnopharmacol2009126218919619735713
- ChoWCLeungKNIn vitro and in vivo immunomodulating and immunorestorative effects of Astragalus membranaceusJ Ethnopharmacol2007113113214117611061
- YangBXiaoBSunTAntitumor and immunomodulatory activity of Astragalus membranaceus polysaccharides in H22 tumor-bearing miceInt J Biol Macromol20136228729024060282
- ChenSMTsaiYSLeeSWAstragalus membranaceus modulates Th1/2 immune balance and activates PPARgamma in a murine asthma modelBiochem Cell Biol201492539740525264079
- AgyemangKHanLLiuEZhangYWangTGaoXRecent advances in Astragalus membranaceus anti-diabetic research: pharmacological effects of its phytochemical constituentsEvid Based Complement Alternat Med2013201365464324348714
- MaoXQYuFWangNHypoglycemic effect of polysaccharide enriched extract of Astragalus membranaceus in diet induced insulin resistant C57BL/6J mice and its potential mechanismPhytomedicine200916541642519201177
- ZouFMaoXQWangNLiuJOu-YangJPAstragalus polysaccharides alleviates glucose toxicity and restores glucose homeostasis in diabetic states via activation of AMPKActa Pharmacol Sin200930121607161519960007
- KoJKChikCWThe protective action of radix Astragalus membranaceus against hapten-induced colitis through modulation of cytokinesCytokine2009472859019539495
- LaiPKChanJYWuSBAnti-inflammatory activities of an active fraction isolated from the root of Astragalus membranaceus in RAW 264.7 macrophagesPhytother Res201428339540423640962
- JiaoJGaiQYWangWUltraviolet radiation-elicited enhancement of isoflavonoid accumulation, biosynthetic gene expression, and antioxidant activity in Astragalus membranaceus hairy root culturesJ Agric Food Chem201563378216822426370303
- ShahzadMShabbirAWojcikowskiKWohlmuthHGobeGCThe antioxidant effects of Radix Astragali (Astragalus membranaceus and related species) in protecting tissues from injury and diseaseCurr Drug Targets Epub201596
- JinMZhaoKHuangQShangPStructural features and biological activities of the polysaccharides from Astragalus membranaceusInt J Biol Macromol20146425726624325861
- SunYYangJExperimental study of the effect of Astragalus membranaceus against herpes simplex virus type 1Di Yi Jun Yi Da Xue Xue Bao2004241575814724098
- SunWYWeiWGuiSYProtective effect of extract from Paeonia lactiflora and Astragalus membranaceus against liver injury induced by bacillus Calmette-Guerin and lipopolysaccharide in miceBasic Clin Pharmacol Toxicol2008103214314918816297
- SunWYWangLLiuHLiXWeiWA standardized extract from Paeonia lactiflora and Astragalus membranaceus attenuates liver fibrosis induced by porcine serum in ratsInt J Mol Med201229349149822109673
- LuoZZhongLHanXWangHZhongJXuanZAstragalus membranaceus prevents daunorubicin-induced apoptosis of cultured neonatal cardiomyocytes: role of free radical effect of Astragalus membranaceus on daunorubicin cardiotoxicityPhytother Res200923676176719172580
- SunMZhouTZhouLFormononetin protects neurons against hypoxia-induced cytotoxicity through upregulation of ADAM10 and sAβPPαJ Alzheimers Dis201228479580822085862
- MaZJiWFuQMaSFormononetin inhibited the inflammation of LPS-induced acute lung injury in mice associated with induction of PPAR gamma expressionInflammation20133661560156623907652
- WangHZhangDGeMFormononetin inhibits enterovirus 71 replication by regulating COX-2/PGE(2) expressionVirol J2015123525890183
- WuXYXuHWuZFFormononetin, a novel FGFR2 inhibitor, potently inhibits angiogenesis and tumor growth in preclinical modelsOncotarget2015642445634457826575424
- ZhangSTangXTianJCardioprotective effect of sulphonated formononetin on acute myocardial infarction in ratsBasic Clin Pharmacol Toxicol2011108639039521232020
- LiCZhouWWangZHuangqi injection attenuates the pancreatic β-cells damage in streptozotocin-induced diabetic ratsInt Med J2014212211214
- HuJFWangWWuJMInhibitory effects of Keluoxin capsule on hypoxia-induced angiogenesis in retinaChin J Exp Tradit Med Form20142010156160
- VaziriKSchwartzSGRelhanNKishorKSFlynnHWJrNew therapeutic approaches in diabetic retinopathyRev Diabet Stud2015121–219621026676668
- JoDHAnHChangDJHypoxia-mediated retinal neovascularization and vascular leakage in diabetic retina is suppressed by HIF-1alpha destabilization by SH-1242 and SH-1280, novel hsp90 inhibitorsJ Mol Med (Berl)201492101083109224875598
- SugimotoMCutlerAShenBInhibition of EGF signaling protects the diabetic retina from insulin-induced vascular leakageAm J Pathol2013183398799523831329
- RosenRVagagginiTChenYZeaxanthin inhibits hypoxia-induced VEGF secretion by RPE cells through decreased protein levels of hypoxia-inducible factors-1alphaBiomed Res Int2015201568738625688362
- PagesGPouyssegurJTranscriptional regulation of the vascular endothelial growth factor gene – a concert of activating factorsCardiovasc Res200565356457315664382
- KuriharaTWestenskowPDFriedlanderMHypoxia-inducible factor (HIF)/vascular endothelial growth factor (VEGF) signaling in the retinaAdv Exp Med Biol201480127528124664708
- KimJHKimJHYuYSShinJYLeeHYKimKWDeguelin inhibits retinal neovascularization by down-regulation of HIF-1alpha in oxygen-induced retinopathyJ Cell Mol Med20081262407241518208562
- DeNiroMAl-HalafiAAl-MohannaFHAlsmadiOAl-MohannaFAPleiotropic effects of YC-1 selectively inhibit pathological retinal neovascularization and promote physiological revascularization in a mouse model of oxygen-induced retinopathyMol Pharmacol201077334836720008515
- IwaseTFuJYoshidaTSustained delivery of a HIF-1 antagonist for ocular neovascularizationJ Control Release2013172362563324126220
- RicciBOxygen-induced retinopathy in the rat modelDoc Ophthalmol19907431711772209374
- Winners-MendizabalOGOrgeFHDi FioreJMMartinRJKcPHypoxia-hyperoxia paradigms in the development of oxygen-induced retinopathy in a rat pup modelJ Neonatal Perinatal Med20147211311725104123
- OseraCMartindaleJLAmadioMInduction of VEGFA mRNA translation by CoCl2 mediated by HuRRNA Biol201512101121113026325091
- MasoudGNLiWHIF-1alpha pathway: role, regulation and intervention for cancer therapyActa Pharm Sin B20155537838926579469
- MarnerosAGFanJYokoyamaYVascular endothelial growth factor expression in the retinal pigment epithelium is essential for choriocapillaris development and visual functionAm J Pathol200516751451145916251428
- LeYZBaiYZhuMZhengLTemporal requirement of RPE-derived VEGF in the development of choroidal vasculatureJ Neurochem201011261584159220067573
- Lyn-CookBDStottmanHLYanYBlannEKadlubarFFHammonsGJThe effects of phytoestrogens on human pancreatic tumor cells in vitroCancer Lett1999142111111910424789
- Kaczmarczyk-SedlakIWojnarWZychMOzimina-KamińskaETaranowiczJSiwekAEffect of formononetin on mechanical properties and chemical composition of bones in rats with ovariectomy-induced osteoporosisEvid Based Complement Alternat Med2013201345705223762138
- ZhangWPetrovicJMCallaghanDEvidence that hypoxia-inducible factor-1 (HIF-1) mediates transcriptional activation of interleukin-1beta (IL-1beta) in astrocyte culturesJ Neuroimmunol20061741–2637316504308
- FilippiICarrarelliPLuisiSDifferent expression of hypoxic and angiogenic factors in human endometriotic lesionsReprod Sci201623449249726408396
- JelkmannWRegulation of erythropoietin productionJ Physiol201158961251125821078592
- ThomasRKimMHHIF-1 alpha: a key survival factor for serum-deprived prostate cancer cellsProstate200868131405141518563715
- KrotovaKPatelJMBlockERZharikovSHypoxic upregulation of arginase II in human lung endothelial cellsAm J Physiol Cell Physiol20102996C1541C154820861464
- BerraEBenizriEGinouvèsAVolmatVRouxDPouysségurJHIF prolyl-hydroxylase 2 is the key oxygen sensor setting low steady-state levels of HIF-1alpha in normoxiaEMBO J200322164082409012912907
- BritoLGSchiavonVFAndradeJMExpression of hypoxia-inducible factor 1-alpha and vascular endothelial growth factor-C in locally advanced breast cancer patientsClinics (Sao Paulo)20116681313132021915477
- CarrollVAAshcroftMRole of hypoxia-inducible factor (HIF)-1alpha versus HIF-2alpha in the regulation of HIF target genes in response to hypoxia, insulin-like growth factor-I, or loss of von Hippel-Lindau function: implications for targeting the HIF pathwayCancer Res200666126264627016778202
- SalcedaSCaroJHypoxia-inducible factor 1alpha (HIF-1alpha) protein is rapidly degraded by the ubiquitin-proteasome system under normoxic conditions. Its stabilization by hypoxia depends on redox-induced changesJ Biol Chem19972723622642226479278421
- TugSDelos ReyesBFandreyJBerchner-PfannschmidtUNon-hypoxic activation of the negative regulatory feedback loop of prolyl-hydroxylase oxygen sensorsBiochem Biophys Res Commun2009384451952319427832
- KaidiAQualtroughDWilliamsACParaskevaCDirect transcriptional up-regulation of cyclooxygenase-2 by hypoxia-inducible factor (HIF)-1 promotes colorectal tumor cell survival and enhances HIF-1 transcriptional activity during hypoxiaCancer Res200666136683669116818642
- ZhouCHZhangXPLiuFWangWModeling the interplay between the HIF-1 and p53 pathways in hypoxiaSci Rep201551383426346319
- EschrichtSJarrKUKuhnCHeat-shock-protein 90 protects from downregulation of HIF-1alpha in calcineurin-induced myocardial hypertrophyJ Mol Cell Cardiol20158511712626031702