
Abstract
c-Kit, a receptor tyrosine kinase, is involved in intracellular signaling, and the mutated form of c-Kit plays a crucial role in occurrence of some cancers. The function of c-Kit has led to the concept that inhibiting c-Kit kinase activity can be a target for cancer therapy. The promising results of inhibition of c-Kit for treatment of cancers have been observed in some cancers such as gastrointestinal stromal tumor, acute myeloid leukemia, melanoma, and other tumors, and these results have encouraged attempts toward improvement of using c-Kit as a capable target for cancer therapy. This paper presents the findings of previous studies regarding c-Kit as a receptor tyrosine kinase and an oncogene, as well as its gene targets and signaling pathways in normal and cancer cells. The c-Kit gene location, protein structure, and the role of c-Kit in normal cell have been discussed. Comprehending the molecular mechanism underlying c-Kit-mediated tumorogenesis is consequently essential and may lead to the identification of future novel drug targets. The potential mechanisms by which c-Kit induces cellular transformation have been described. This study aims to elucidate the function of c-Kit for future cancer therapy. In addition, it has c-Kit inhibitor drug properties and their functions have been listed in tables and demonstrated in schematic pictures. This review also has collected previous studies that targeted c-Kit as a novel strategy for cancer therapy. This paper further emphasizes the advantages of this approach, as well as the limitations that must be addressed in the future. Finally, although c-Kit is an attractive target for cancer therapy, based on the outcomes of treatment of patients with c-Kit inhibitors, it is unlikely that Kit inhibitors alone can lead to cure. It seems that c-Kit mutations alone are not sufficient for tumorogenesis, but do play a crucial role in cancer occurrence.
Keywords:
Introduction
c-Kit, a type III receptor tyrosine kinase (RTK), plays a crucial role in cancer occurrence.Citation1 Currently, c-Kit is mainly considered a stem cell factor (SCF), which participates in vital functions of the human body, such as fertility, homeostasis, and melanogenesis; nevertheless, early studies on c-Kit introduced it as an oncogene.Citation2,Citation3 Deregulation of c-Kit, including overexpression and gain of function mutations, has been detected in several human cancers. Leukemia is the first cancer linked to c-Kit activating mutation.Citation4 Subsequent studies reported that c-Kit activating mutation is found in almost all cases of systemic mastocytosis and other hematopoietic cancers; these findings support the hypothesis that the c-Kit target is possibly located in the stem cell compartment.Citation5 c-Kit has been reported to be mostly correlated with gastrointestinal stromal tumor (GIST), with 80% of all GIST cases involving c-Kit activating mutation. As such, the use of Kit inhibitors has provided novel insights for cancer treatment.Citation6 In addition, Kit mutations have been detected in cancers such as leukemia,Citation7 unilateral ovarian dysgerminoma,Citation8–Citation10 melanoma,Citation11 and others.Citation12–Citation14 Evidence reveals that targeting c-Kit as an oncogene by using kinase inhibitor drugs such as imatinib is a promising approach for cancer treatment. However, several issues have been raised regarding this approach. For instance, resistance to imatinib, a famous c-Kit inhibitor drug, has been observed in several cases and is attributed to changes in c-Kit mutations; moreover, c-Kit is expressed in normal tissues such as breast epithelial, vascular endothelial, sweat glands, and retinal astrocytes.Citation15 In this regard, c-Kit mutations cannot be considered a risk factor for cancer occurrence.Citation16 Therefore, targeting c-Kit for cancer treatment is only feasible in cases where c-Kit is the “driver” of the cancer.
Gene and protein structures of c-Kit
c-Kit, a protooncogene in a region on the long arm of chromosome 4 (4q11–4q13), encodes the SCF receptor (CD117).Citation17,Citation18 c-Kit is the cellular equivalent of the v-kit oncogene, a transforming feline retrovirus, and a 145 kDa transmembrane glycoprotein, which belongs to class III of the RTK family. This family is categorized into three domains: a hydrophobic transmembrane, an extracellular ligand-binding domain, and a cytoplasmic domain with tyrosine kinase activity.Citation19
Four c-Kit isomers caused by alternative RNA splicing have been found in humans.Citation20 The presence of serine residues in the kinase insert region differentiates the two isoforms, though the function of a serine residue is still unknown. A stretch of four acids on the extracellular side also distinguishes the two other isoforms. At the molecular level, these isoforms differ in terms of ability to induce signal transduction and tumorigenic potential.Citation21–Citation26 The isoform without the tetrapeptide sequence is regarded as the strongest inducer and highest transformer.Citation27
Another c-Kit isoform has been detected in murine testis; this isoform is truncated resulting from the controlled promoter element within intron 16, which contains 12 amino acids and a carboxyterminal tail without kinase activity.Citation28 This isoform has also been found to be expressed in human prostate cancers.Citation29 By contrast, one study reported that this isoform is mouse specific and cannot be found in humans.Citation30
c-Kit in normal stem cells
c-Kit, an SCF receptor,Citation1 plays an important role in stem cell maintenance and differentiation.Citation31 c-Kit expression has been detected in various stem cells or cells with self-renewal potency and progenitor cells.Citation32 Studies have also confirmed that c-Kit is expressed in different kinds of stem cells, especially hematopoietic cells.Citation33,Citation34 In several loss-of-function mutations of c-Kit, the mutated site has been linked to a wide range of defects, from minor defects in catalytic activity to critical flaws in the hematopoietic system in mice.Citation35 c-Kit mutations has also been reported to significantly affect other systems such as the reproductive, pigmentation, and nervous system.Citation36,Citation37
Hematopoietic stem cells divide asymmetrically and can self-renew or differentiate into all hematopoietic cell lineages, including myeloid (monocytes and macrophages, neutrophils, basophils, eosinophils, erythrocytes, megakaryocytes/platelets, and dendritic cells) and lymphoid (T-cells, B-cells, and NK-cells) lineages.Citation38 During differentiation, stemness and self-renewal are lessened, whereas cell proliferation is maintained; this phenomenon leads to increased cell numbers.Citation39 Arguably, c-Kit, as an SCF receptor, is expressed in original hematopoietic cells, such as stem and progenitor cells. However, c-Kit expression vanishes during differentiation.Citation40–Citation43 Less than 1% of peripheral blood cells express c-Kit, which indicates the minimal role of c-Kit in differentiated hematopoietic cells.Citation44–Citation46 Furthermore, c-Kit is highly expressed in mast cells; as such, completely differentiated cells depend on c-Kit signal transduction for their survival, function, and growth.Citation47 Finally, among lymphoid lineages, c-Kit is involved in immune system maintenance in adult animals.Citation48
c-Kit affects germ cells in the fertility system, which are classified as progenitor cells.Citation48 Previous studies showed that c-Kit mutations most likely and normally leads to the protection of germ cells from apoptosis and to the induction of cell migration and proliferation.Citation49 The pathway necessary for apoptosis inhibition by c-Kit is the PI3 kinase/AKT pathway, which is essential in cellular processes such as protein synthesis, proliferation, and survival.Citation50 Mutant mice overexpressing the c-Kit receptor cannot activate PI3 kinase, leading to infertility.Citation51 c-Kit is also involved in germ cell biology, particularly during gametogenesis in male and female mice.
c-Kit exhibits other functions. Mutant mice underexpressing c-Kit present defects in pigmentation, which is related to the ability of SCF (a c-Kit cognate ligand) to induce proliferation and guide the migration of melanocytes from the neuronal crest to the dermis.Citation52–Citation54 Other studies revealed two tyrosine residues, namely, 568 and 570, which are necessary for normal pigmentation.Citation55 The loss-of-function mutations of c-Kit has been detected in majority of humans with piebaldism syndrome, a rare autosomal dominant disorder of melanocyte development. This syndrome is characterized by hypopigmentation in the stomach, hair, and extremities as a result of melanocyte deficiency.Citation56 Furthermore, c-Kit activity influences the digestive and nervous systems. c-kit is suggested to be involved in the interstitial cells of Cajal or (ICC) in sum. Loss of ICC is correlated with diseases such as slow transit constipation.Citation57 Previous studies suggested the vital role of c-Kit in ICC development or function.Citation58–Citation60 In mice with c-Kit loss-of-function mutation, ICC is depleted.Citation57 Several studies reported the role of c-Kit in the development and/or function of nervous system cells. c-Kit expression has also been detected in neuroproliferative cells in adult rat brains, in addition to neural cultures.Citation61 Moreover c-Kit signaling is significant in migration of neural stem cells to injured areas of the brain.Citation62 These studies suggest that c-Kit plays a crucial role in the stem and progenitor cells of different systems of the body and that loss-of-function mutations in c-Kit can lead to several defects.
c-Kit and cancers
In this section, we discuss the previous findings of c-Kit deregulations in several types of cancer. As will be shown, there are different type of deregulation of c-Kit, and each of them can result in tumorogenesis. Deregulation of c-Kit can result in cancer in different ways. This deregulation could occur in different ways such as gain of function, loss of function, overexpression, and point mutations.Citation16 The role of c-Kit deregulation in cancer was first identified as a retroviral oncogene using mice as a reference. The role of c-Kit in cancer has not been completely uncovered. c-Kit is a marker for human acute myeloid leukemia (AML) and normal hemopoietic progenitor cells.Citation1 Studies demonstrated the crucial function of c-Kit and its ligand in hematopoiesis,Citation63 fertility,Citation64,Citation65 and melangenesis.Citation66
In a number of cancers, c-Kit activation was detected through overexpression or mutations. Conversely, in other tumors, such as melanoma,Citation67,Citation68 thyroid carcinoma,Citation69 and breast cancer,Citation69 loss-of-function mutation of c-Kit was observed. Moreover, c-Kit gain-of-function mutation in metastatic melanoma induces apoptosis.Citation70 By contrast, in uveal melanomas, c-Kit expression results in cell proliferation, for which treatment with kinase inhibitor drugs leads to apoptosis induction.Citation71 The activating mutation of c-Kit, namely, L576P, has been reported in a small subset of highly metastatic melanomas.Citation72 Thus, there are various c-Kit mutations involved in melanoma, so melanoma can be used as a model to clarify the complex roles of c-Kit in tumorigenesis.Citation73 In certain cancer types, such as GIST, the main cause of molecular events in tumorogenesis is the activating mutations in c-Kit. In this case, targeting c-Kit with imatinib mesylate increases survival by approximately 70%–80% after 2 years compared with cancer treatment without c-Kit targeting drugs.Citation74 In cancer cases where activating mutation in c-Kit is not the causative event, treatment targeting c-Kit results in poor treatment outcomes.
c-Kit mutations were first reported in human mast cell leukemia.Citation4 These mutations have also been detected in GIST.Citation12 Furthermore, c-Kit mutations have been found in most cases of systemic mastocytosis.Citation5
The activation of c-Kit mutations is rarely detected in some cancer types. For instance, c-Kit activation mutations are uncommon in AMLCitation75 and rarely detected in other cancer types; for example, only 26% of germ cell cancers, or more specifically, testicular seminomas, have been associated with c-Kit mutations.Citation8 These mutations have not been detected in urinary and ovarian cancers and only in 30% cases of urinal ovarian dysgerminoma.Citation8 Moreover, a low frequency of activating c-Kit mutation and proliferation is found in melanoma.Citation76
Different types of c-Kit mutations can cause tumors. The concurrent overexpression of c-Kit and its ligands occur in some types of tumors such as colorectal carcinoma, breast carcinoma, small-cell lung carcinoma, neuroblastoma, and gynecological tumor.Citation77 Tumors resistant to chemotherapy, such as malignant mesothelioma, have been shown to be associated with simultaneous upregulated expression of c-Kit and its ligand.Citation78
In several tumors, c-Kit overexpression is found without mutation and/or mutation in its ligand. In normal physiological circumstances, only a minority of hematopoietic cells express c-Kit. AML cells express c-Kit, which influences the malignant phenotype of this cancer.Citation3,Citation46 A previous study reported that c-Kit expression level is 7.4-fold higher in renal oncocytoma and chromophobe renal carcinoma than that in renal normal tissues.Citation79 c-Kit overexpression has not been observed in other types of renal cancers.Citation80
In cancers involving RTKs, gain-of-function mutations are the main events that lead to cancer progression.Citation81 The first gain-of-function c-Kit mutation was identified in the human mast cell line HMC1. These mutations, known as D816V and V560G, are located in the juxtamembrane region and in the tyrosine kinase domain, respectively.Citation4 Mutations in the juxtamembrane domain are disrupted by the interaction of this region with the kinase domain, which results in inhibited mutation. Furthermore, point mutations in this region can result in induced c-Kit dimerization.Citation82 These events consequently induce the activation of kinase domains.
Another mutation hotspot is found in codon 816, located in the second part of the kinase domain, and leads to the activation of the domain. This mutation is considered as ligand-independent activation because of the switching of aspartic acid residues to asparagine, tyrosine, valine, or histidine residues.Citation83 Nevertheless, other studies provide contradicting results, in which the kinase domain forms a dimer in the absence of SCF.Citation82,Citation84,Citation85 Furthermore, studies showed that both mutants in the kinase domain (D814Y) and juxtamembrane domain (KΔ27) (in frame deletion at codons 547–555) lead to substrate alteration, which results in significant outcomes based on signaling pathway(s) activated by the mutant c-Kit.Citation86,Citation87
A broad range of juxtamembrane domain mutations have been detected in GIST; these mutations include duplications, point mutations, deletions, or their combinations. Mutations in this region have also been detected in approximately 30% of AML,Citation88 sinonasal lymphoma,Citation89 and rare cases of mastocytosis.Citation90
A wide range of human cancers are caused by mutations in the c-Kit kinase domain; these cancers include different types of leukemia, such as acute myeloid, core-factor binding, and mast cell leukemia;Citation7,Citation91,Citation92 testicular germ cell tumor; intracranial and ovarian dysgerminoma;Citation9,Citation93,Citation94 mastocytosis;Citation14–Citation95 and papillary renal carcinomas.Citation13
Hotspot region of Kit mutation
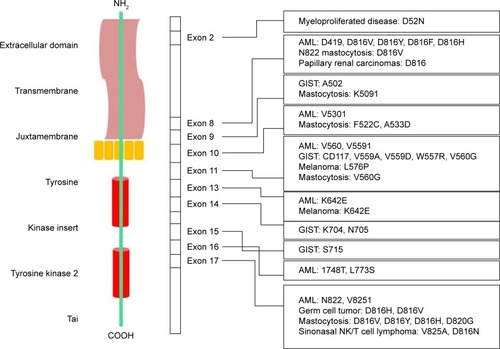
Numerous Kit mutation sites are found and vary in different cancer types,Citation16 reflecting the effect of each mutation on downstream signaling pathways.Citation12 Some “hotspots” in the Kit gene are regular in certain main domain structures.Citation113 Mutations in domains, such as intracellular and extracellular juxtamembranes, located on exons 8, 9, and 11,Citation12 as well as exon 17, which corresponds to the activation loop in the kinase domain,Citation114 disrupt the autoinhibitory mechanisms of Kit.Citation115,Citation116
The importance of these two domains is reflected in their critical role in the Kit structure and function. The first group of extracellular juxtamembrane domains is responsible for the correct binding of the receptor monomer and the stabilization of dimers, which contain dimeric SCF.Citation117 The mutations in this region, particularly in exons 8 and 9, are detected in AML and GIST, respectively.Citation118,Citation119 The most common mutations in c-Kit are summarized in and are classified based on type of cancer, exon location, and the type of mutation.
Table 1 The most common mutations in c-Kit
Kit mutations occur within exon 11 in almost 65% of all GIST cases. This exon encodes a key autoregulatory domain of the RTK, which is the intracellular juxtamembrane domain, and stabilizes the inactive conformation of the kinase domain.Citation121 In addition, mutations in other exons, such as exons 11 and 17, have been identified in GISTCitation122 and hematological cancers,Citation123 respectively.
The most detected c-Kit mutations that lead to melanoma are located within exons 11 and 13, namely, L576P and K642E, respectively.Citation76 Mutations in exon 17, which encodes the activation loop of the kinase domain, result in hemopoietic malignancies in germ cell tumors. After kinase activation, conformational shifts occur in this region.Citation116 The KIT cDNA structure in different cancers and their respective mutations is illustrated in . c-Kit mutation based on their gene location, corresponding cancer, and drug sensitivity are listed in .
Table 2 c-Kit mutation based on their gene location, corresponding cancer, and drug sensitivity
Figure 1 KIT cDNA and protein structure in different cancers and their respective mutations.
c-Kit signal transduction
c-Kit is involved in several signaling pathways, as discussed.
PI3-kinase
PI3-kinase is the most studied pathways involving c-Kit. In this pathway, phosphatidylinositide 3′-kinase phosphorylates the 3′-hydroxyl group of the inositol ring of lipids in the cell membrane.Citation79 As such, the negative electric potential across the lipid increases, which results in the interaction of PIP3 in the cell membrane with proteins comprising pleckstrin homology (PH) domains. Consequently, the PH domain transduces proteins from the cytoplasm to the plasma, thus activating AKT. This activation is crucial for the apoptosis ability of SCF. Serine/threonine kinase AKT is a vital PH domain that contains proteins in c-Kit signaling.Citation34
Recent studies revealed that SCF interacts with the transcription factor FOXO3a, which leads to survival through AKT-mediated phosphorylation; this factor belongs to the O subclass of the forkhead family of transcription factors and is characterized by a diverse forkhead DNA-binding domain.Citation34,Citation81 As a result, the expression of proto-apoptotic protein Bim decreases and Mek-dependent phosphorylation is downregulated.Citation80 In addition, PI3-kinase plays an important role in SCF-induced proliferation and regulation of the actin cytoskeleton and cell migration.Citation83 Earlier studies confirmed the necessity of PI3-kinase in hematopoietic cell growth and tumorigenicity, which involves the active form of c-Kit in the genome.Citation84,Citation85
These studies focused on class I PI3-kinase, despite that class II PI3KC2β is significantly associated with c-Kit.Citation86 Studies have also illustrated the interaction of C2 domains with phosphotyrosine residues,Citation87 which can prove the probability of interaction between PI3K-C and c-Kit activation.
Src family kinase
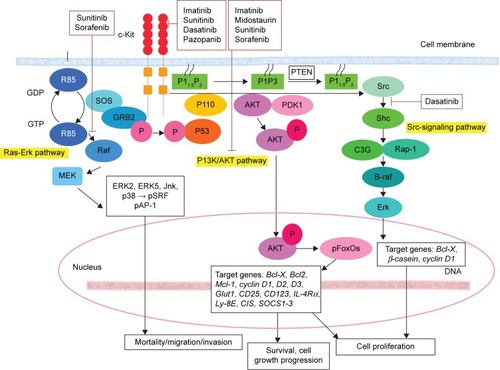
The other signaling pathway that stimulates c-Kit activity is the Src family kinases (SFKs), which are cytoplasmic tyrosine kinases. This pathway is involved in several crucial biological functions such as survival, chemotaxis, and proliferation.Citation88 Although studies prove that SFKs are activated by c-Kit, the function of this activation in the pathway remains unknown.Citation89,Citation90 In addition, the involvement of SFK in c-Kit internalization has been demonstrated at the cell biological level.Citation91
The contribution of Lyn in c-Kit, STAT3, and JNK phosphorylation was determined using Lyn−/− bone marrow mast cells.Citation92 PI3-kinase/AKT signaling is negatively regulated by Lyn, although the underlying mechanism remains unknown. SFK evidently plays a role in SCF-induced chemotaxis and proliferation of primary hematopoietic progenitor cells.Citation91 In the study by Hong et al,Citation93 it was demonstrated that SFK affects the downstream pathways of c-Kit, although other signaling molecules, such as APS, SHP1, and SHP2, also interact with the SFK-binding site.Citation94,Citation95
Phenotypic analysis on mutant c-Kit mice with inability to interact with SFK showed that SFK plays a role(s) in c-Kit signaling in lymphocytes.Citation39,Citation55 In addition to lymphocyte defects, severe problems in pigmentation, splenomegaly, and mast cell development have been observed in double c-Kity567/569f mutations. Thus, PI3-kinase and SFK are two important signaling pathways in most tissues, and any defect in these pathways can interrupt their interaction with c-Kit could lead to severe defects.Citation150
Ras-Erk pathway
The next signaling pathway that stimulates the activity of c-Kit is the Ras-Erk pathway. The activation of MAP-kinases, namely, Erk1/2, plays a vital role in cell proliferation, differentiation, and survival. The main role of c-Kit in this signaling includes the recruitment of the guanine exchange factor Sos to its substrate, namely, the small GTPase Ras, which is located in the plasma membrane. By converting GTP to GDP, Sos provides the guanine nucleotide phosphate which binds to Ras, resulting in Ras activation. Active Ras consequently translocates Raf-1 to the plasma membrane. Mek, another kinase, and Erk are then activated. Finally, the activated Erk results in the alteration of protein activity and gene expression.Citation97
In terms of the relevance of c-Kit to this pathway, a special region is found in c-Kit, namely, tyrosine residues 703 and 936, which directly interacts with the stable complex of Sos with the adaptor protein Grb2.Citation98 Altogether, c-Kit is considered a target for cancer therapy because it mediates the activation of Erk1/2. In addition, in HL60 cells, Erk activation upregulates survivin expression by SCF mediation, which leads to resistance toward apoptosis induced by radiation.Citation102
These signaling pathways are activated by Kit. Other important signaling pathways involving c-Kit include the JAK/STAT pathway and various signaling proteins, such as adaptor proteins (eg, Crk, Gab, APS, Gads, ShcA, Grb2, and Grb7), cytoplasmic tyrosine kinases, and protein tyrosine phosphatases.Citation39,Citation94,Citation101,Citation102,Citation151 The role of c-Kit in the molecular function of the aforementioned signaling proteins is characterized to varied degrees. With respect to the diverse functions of c-Kit, large numbers of these proteins are affected in different ways. In addition to the direct or indirect effect of c-Kit on various signaling pathways or proteins, this kinase often functions in cooperation with other cytokines and growth factors.Citation150,Citation152 The summary of the signaling pathway with involvement of c-Kit in normal and cancer cells is demonstrated in and , respectively.
Figure 2 Signal transducer and activator of transcription tyrosine kinase domain c-Kit in normal cell.
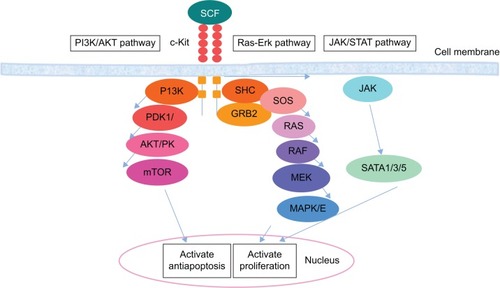
Figure 3 Signal transducer and activator of transcription tyrosine kinase domain c-Kit in cancer cell.
c-Kit: a potential target for cancer therapy
Kinase superfamily proteins are considered a main target for molecular cancer therapy. Numerous studies have uncovered the molecular chronicles that occur during cancer development. The gist of most researchers is that kinases are an essential factor for cancer progression and are overexpressed by tumors. As such, kinases should be targeted as a new method for cancer treatment. Different approaches have been considered for this strategy. The first approach uses antibodies against kinase proteins. For instance, trastuzumab (Herceptin®; Genentech Inc, South San Francisco, CA, USA) targets the extracellular domain of HER2. Other antibodies, such as low-molecular-weight kinase inhibitors, eg, gefitinib (Iressa®; AstraZeneca, London, UK), aim at enzymatic activity. The first group can only affect proteins with an extracellular domain, whereas the second group can target both transmembrane and intracellular proteins. The first group of proteins is very specific, whereas the specificity of the second group of proteins may be limited. Most kinase inhibitors target and bind to the enzymatic domain and compete with ATP; however, the specificity of these inhibitors are conserved because of unique binding patterns.Citation101
The major concern in cancer management is the occurrence of resistance toward drugs. This resistance could be due to mutations in the target protein, which result in the reduction of the binding between the drug and the kinase. In addition, overexpression of transport proteins by cells may occur, which leads to decreased intracellular concentration of drugs. Additionally, some other oncogenes may substitute for or reimburse the inhibition of the drug target. For instance, for the drug used in chronic myelogenous leukemia (imatinib mesylate), drug resistance has been observed as a result of both the overexpression of Bcr-Abl and mutation in the kinase domain, which is specific for drug binding.Citation104 c-Kit mutation based on their gene location, corresponding cancer, and drug sensitivity are listed in .
Low-molecular- weight inhibitors have been utilized for c-Kit targeting. For instance, imatinib mesylate is one of the c-Kit targeting drugs that inhibit both Abl and PDGFRs. List of the drugs that target c-Kit have been listed in , and are classified based on drug name, molecular formula, common and specific targets, structural formula, and general function. The list of c-Kit inhibitors with their complete list of their targets, besides c-Kit, with emphasize of their affectivity on c-Kit is listed in . In addition, c-Kit inhibitors classification based on their targets, chemical and structure formulae, and diseases they are tested on, have been listed in .
Table 3 List of the drugs that target c-Kit
Table 4 List of c-Kit inhibitors
Table 5 c-Kit inhibitors classification based on their targets, chemical and structure formulae, and diseases they are tested on
Conclusion
Overall, particular mutations in c-Kit are accountable for cancer occurrence, such as GIST and SM (Systemic Mastocytosis) cases, as well as subsets of AML and melanoma. As the c-Kit mutations are the “drivers” in these cases, the use of kinase inhibitors, such as imatinib, could significantly improve cancer treatment. Nevertheless, the second mutation in c-Kit, which disturbs the biding region of kinase inhibitor, results in drug resistance. Thus far, no alternative solution for the current problem is established. This limitation could be addressed by elucidating the c-Kit pathway and its targeting genes. Cotargeting these pathways may also lead to tumor control. Finally, inhibition of cancer stem cells would be vital when considering the crucial role of cancer stem cells in drug resistance and cancer recurrence in cancer therapy.
Acknowledgments
The authors would like to express their utmost gratitude and appreciation to the University of Malaya Research Grant (RG084-13BIO), RP024A/14HTM UMRP grant, IPPP grant (PG082-2013B), the BKP grant (BK020-2012), and the Malaysian Ministry of Higher Education HIR Grants (UM.C/625/1/MOHE/MED/17 and UM.C/625/1/MOHE/MED/33) for providing financial support to conduct this study. The authors are thankful to Selleckchem.com for the reuse of their data.
Disclosure
The authors report no conflicts of interest in this work.
References
- AshmanLKThe biology of stem cell factor and its receptor c-kitInt J Biochem Cell Biol199931101037105110582338
- FuritsuTTsujimuraTTonoTIdentification of mutations in the coding sequence of the proto-oncogene c-kit in a human mast cell leukemia cell line causing ligand-independent activation of c-kit productJ Clin Invest199392417367691885
- YavuzASLipskyPEYavuzSMetcalfeDDAkinCEvidence for the involvement of a hematopoietic progenitor cell in systemic mastocytosis from single-cell analysis of mutations in the c-kit geneBlood2002100266166512091362
- CorlessCLFletcherJAHeinrichMCBiology of gastrointestinal stromal tumorsJ Clin Oncol200422183813382515365079
- YardenYKuangWJYang-FengTHuman proto-oncogene c-kit: a new cell surface receptor tyrosine kinase for an unidentified ligandEMBO J198761133412448137
- GiebelLStrunkKHolmesSSpritzROrganization and nucleotide sequence of the human KIT (mast/stem cell growth factor receptor) proto-oncogeneOncogene1992711220722171279499
- UllrichASchlessingerJSignal transduction by receptors with tyrosine kinase activityCell19906122032122158859
- ReithAEllisCLymanSSignal transduction by normal isoforms and W mutant variants of the Kit receptor tyrosine kinaseEMBO J199110924511714377
- VoytyukOLennartssonJMogiASrc family kinases are involved in the differential signaling from two splice forms of c-KitJ Biol Chem2003278119159916612511554
- MaulikGBhartiAKhanEBroderickRJKijimaTSalgiaRModulation of c-Kit/SCF pathway leads to alterations in topoisomerase-I activity in small cell lung cancerJ Environ Pathol Toxicol Oncol200423423725115511212
- PawsonTProtein modules and signalling networksNature199537365155735807531822
- Blume-JensenPSiegbahnAStabelSHeldinCHRönnstrandLIncreased Kit/SCF receptor induced mitogenicity but abolished cell motility after inhibition of protein kinase CEMBO J1993121141997693453
- ZhuWMDongWFMindenMAlternate splicing creates two forms of the human kit proteinLeuk Lymphoma1994125–64414477514064
- CrosierPRicciardiSTHallLRVitasMRClarkSCCrosierKEExpression of isoforms of the human receptor tyrosine kinase c-kit in leukemic cell lines and acute myeloid leukemiaBlood1993824115111587688988
- CaruanaGCambareriACAshmanLKIsoforms of c-KIT differ in activation of signalling pathways and transformation of NIH3T3 fibroblastsOncogene1999185573558110523834
- HuangENockaKBuckJBesmerPDifferential expression and processing of two cell associated forms of the kit-ligand: KL-1 and KL-2Mol Biol Cell1992333493621378327
- AlbanesiCGeremiaRGiorgioMDolciSSetteCRossiPA cell- and developmental stage-specific promoter drives the expression of a truncated c-kit protein during mouse spermatid elongationDevelopment19961224129113028620856
- ParonettoMPFariniDSammarcoIExpression of a truncated form of the c-Kit tyrosine kinase receptor and activation of Src kinase in human prostatic cancerAm J Pathol200416441243125115039213
- SakamotoAYonedaATeradaKA functional truncated form of c-kit tyrosine kinase is produced specifically in the testis of the mouse but not the rat, pig, or humanBiochem Genet20044211–1244145115587987
- DaSilvaNHuZMaWRosnetOBirnbaumDDrexlerHExpression of the FLT3 gene in human leukemia-lymphoma cell linesLeukemia1994858858888182945
- HuZBMaWUphoffCQuentmeierHDrexlerHc-kit expression in human megakaryoblastic leukemia cell linesBlood1994838213321447512841
- RussellESHereditary anemias of the mouse: a review for geneticistsAdv Genet197920357390999
- KeshetELymanSWilliamsDEmbryonic RNA expression patterns of the c-kit receptor and its cognate ligand suggest multiple functional roles in mouse developmentEMBO J199110924251714375
- LevSBlechmanJMGivolDYardenYSteel factor and c-kit protooncogene: genetic lessons in signal transductionCrit Rev Oncog199452–31411687531500
- OkadaSNakauchiHNagayoshiKNishikawaSMiuraYSudaTEnrichment and characterization of murine hematopoietic stem cells that express c-kit moleculeBlood1991787170617121717068
- BroudyVCStem cell factor and hematopoiesisBlood1997904134513649269751
- LymanSDJacobsenSEWc-Kit ligand and Flt3 ligand: stem/progenitor cell factors with overlapping yet distinct activitiesBlood1998914110111349454740
- AshmanLKCambareriACToLBLevinskyRJJuttnerCAExpression of the YB5. B8 antigen (c-kit proto-oncogene product) in normal human bone marrowBlood199178130371712644
- PapayannopoulouTBriceMBroudyVCZseboKMIsolation of c-kit receptor-expressing cells from bone marrow, peripheral blood, and fetal liver: functional properties and composite antigenic profileBlood1991786140314121715789
- BroudyVCSmithFLinNZseboKEgrieJBernsteinIBlasts from patients with acute myelogenous leukemia express functional receptors for stem cell factorBlood199280160671377054
- OliveiraSLukacsNStem cell factor: a hemopoietic cytokine with important targets in asthmaCurr Drug Targets Inflamm Allergy20032431331814561150
- WaskowCPaulSHallerCGassmannMRodewaldHRViable c-Kit W/W mutants reveal pivotal role for c-kit in the maintenance of lymphopoiesisImmunity200217327728812354381
- LovelandKSchlattSStem cell factor and c-kit in the mammalian testis: lessons originating from Mother Nature’s gene knockoutsJ Endocrinol199715333373449203987
- Blume-JensenPJanknechtRHunterTThe kit receptor promotes cell survival via activation of PI 3-kinase and subsequent Akt-mediated phosphorylation of Bad on Ser136Curr Biol19988137797859651683
- KisselHTimokhinaIHardyMPPoint mutation in kit receptor tyrosine kinase reveals essential roles for kit signaling in spermatogenesis and oogenesis without affecting other kit responsesEMBO J20001961312132610716931
- ScottGEwingJRyanDAbboudCStem cell factor regulates human melanocyte-matrix interactionsPigment Cell Res19947144517521051
- MackenzieMAJordanSABuddPSJacksonIJActivation of the receptor tyrosine kinase Kit is required for the proliferation of melanoblasts in the mouse embryoDev Biol19971921991079405100
- Wehrle-HallerBThe role of Kit-ligand in melanocyte development and epidermal homeostasisPigment Cell Res200316328729612753403
- KimuraYJonesNKlüppelMTargeted mutations of the juxtamembrane tyrosines in the Kit receptor tyrosine kinase selectively affect multiple cell lineagesProc Natl Acad Sci U S A2004101166015602015067126
- LyfordGHeCSofferEPan-colonic decrease in interstitial cells of Cajal in patients with slow transit constipationGut200251449650112235070
- HuizingaJDThunebergLKluppelMMalyszJMikkelsenHBBernsteinAW/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activityNature199537365123473497530333
- WardSMBurnsAJTorihashiSHarneySCSandersKMImpaired development of interstitial cells and intestinal electrical rhythmicity in steel mutantsAm J Physiol19952696C1577C15858572188
- WardSMBurnsAJTorihashiSSandersKMMutation of the proto-oncogene c-kit blocks development of interstitial cells and electrical rhythmicity in murine intestineJ Physiol1994480191977853230
- JinKMaoXOSunYXieLGreenbergDAStem cell factor stimulates neurogenesis in vitro and in vivoJ Clin Invest2002110331131912163450
- SunLLeeJFineHANeuronally expressed stem cell factor induces neural stem cell migration to areas of brain injuryJ Clin Invest20041139136415124028
- AshmanLKGriffithRTherapeutic targeting of c-KIT in cancerExpert Opin Investig Drugs2013221103115
- LennartssonJJelacicTLinnekinDShivakrupaRNormal and oncogenic forms of the receptor tyrosine kinase kitStem Cells2005231164315625120
- HuangSLucaMGutmanMEnforced c-KIT expression renders highly metastatic human melanoma cells susceptible to stem cell factor-induced apoptosis and inhibits their tumorigenic and metastatic potentialOncogene19961311233923478957075
- Willmore-PayneCHoldenJATrippSLayfieldLJHuman malignant melanoma: detection of BRAF-and c-kit–activating mutations by high-resolution amplicon melting analysisHum Pathol200536548649315948115
- D’AmatoGSteinertDMMcAuliffeJCTrentJCUpdate on the biology and therapy of gastrointestinal stromal tumorsCancer Control2005121445615668652
- HirotaSIsozakiKMoriyamaYGain-of-function mutations of c-kit in human gastrointestinal stromal tumorsScience199827953505775809438854
- KellyLMGillilandDGGenetics of myeloid leukemiasAnnu Rev Genomics Hum Genet20023117919812194988
- Hoei-HansenCEKraggerudSMAbelerVMKærnJRajpert-De MeytsELotheRAOvarian dysgerminomas are characterised by frequent KIT mutations and abundant expression of pluripotency markersMol Cancer2007611217274819
- BeadlingCJacobson-DunlopEHodiFSKIT gene mutations and copy number in melanoma subtypesClin Cancer Res200814216821682818980976
- HeinrichMCBlankeCDDrukerBJCorlessCLInhibition of KIT tyrosine kinase activity: a novel molecular approach to the treatment of KIT-positive malignanciesJ Clin Oncol20022061692170311896121
- CatalanoARodilossiSRippoMRCaprariPProcopioAInduction of stem cell factor/c-Kit/slug signal transduction in multidrug-resistant malignant mesothelioma cellsJ Biol Chem200427945467064671415337769
- IkedaHKanakuraYTamakiTExpression and functional role of the proto-oncogene c-kit in acute myeloblastic leukemia cellsBlood19917811296229681720040
- HuoLSugimuraJTretiakovaMSc-kit expression in renal oncocytomas and chromophobe renal cell carcinomasHum Pathol200536326226815791570
- RodriguesGAParkMOncogenic activation of tyrosine kinasesCurr Opin Genet Devel19944115248193535
- KitayamaHKanakuraYFuritsuTConstitutively activating mutations of c-kit receptor tyrosine kinase confer factor-independent growth and tumorigenicity of factor-dependent hematopoietic cell linesBlood19958537907987530509
- MoriyamaYTsujimuraTHashimotoKRole of aspartic acid 814 in the function and expression of c-kit receptor tyrosine kinaseJ Biol Chem19962717334733508631931
- LamLChowRBergerSA transforming mutation enhances the activity of the c-Kit soluble tyrosine kinase domainBiochem J19993381311389931308
- TsujimuraTHashimotoKKitayamaHActivating mutation in the catalytic domain of c-kit elicits hematopoietic transformation by receptor self-association not at the ligand-induced dimerization siteBlood1999934131913299949175
- CasteranNDe SepulvedaPBesluNSignal transduction by several KIT juxtamembrane domain mutationsOncogene200322304710472212879016
- PiaoXPaulsonRVan Der GeerPPawsonTBernsteinAOncogenic mutation in the Kit receptor tyrosine kinase alters substrate specificity and induces degradation of the protein tyrosine phosphatase SHP-1Proc Natl Acad Sci U S A1996932514665146698962111
- NaoeTKiyoiHNormal and oncogenic FLT3Cell Mol Life Sci200461232932293815583855
- HongyoTLiTSyaifudinMSpecific c-kit mutations in sinonasal natural killer/T-cell lymphoma in China and JapanCancer Res20006092345234710811105
- BüttnerCHenzBMWelkerPSeppNTGrabbeJIdentification of activating c-kit mutations in adult-, but not in childhood-onset indolent mastocytosis: a possible explanation for divergent clinical behaviorJ Investig Dermatol19981116122712319856847
- AshmanLKFerraoPColeSRCambareriACEffects of mutant c-kit in early myeloid cellsLeuk Lymphoma1999345–645146110492068
- BeghiniAPeterlongoPRipamontiCBc-kit mutations in core binding factor leukemiasBlood200095272672810660321
- TianQFriersonHFKrystalGWMoskalukCAActivating c-kit gene mutations in human germ cell tumorsAm J Pathol199915461643164710362788
- PaulsKWardelmannEMerkelbach-BruseSBüttnerRZhouHc-KIT codon 816 mutation in a recurrent and metastatic dysgerminoma of a 14-year-old girl: case studyVirchows Arch2004445665165415455230
- SakumaYSakuraiSOguniSSatohMHironakaMSaitoKc-Kit gene mutations in intracranial germinomasCancer Sci200495971672015471556
- NagataHWorobecASOhCKIdentification of a point mutation in the catalytic domain of the protooncogene c-kit in peripheral blood mononuclear cells of patients who have mastocytosis with an associated hematologic disorderProc Natl Acad Sci U S A1995922310560105647479840
- LongleyBJTyrrellLLuSZSomatic c-KIT activating mutation in urticaria pigmentosa and aggressive mastocytosis: establishment of clonality in a human mast cell neoplasmNature Genet19961233123148589724
- LinZHHanEMLeeESA distinct expression pattern and point mutation of c-kit in papillary renal cell carcinomasMod Pathol200417661161615073597
- HubbardSRJuxtamembrane autoinhibition in receptor tyrosine kinasesNat Rev Mol Cell Biol20045646447115173825
- DiNittoJPDeshmukhGDZhangYFunction of activation loop tyrosine phosphorylation in the mechanism of c-Kit auto-activation and its implication in sunitinib resistanceJ Biochem2010147460160920147452
- MorganSJSmithADParkerPJPurification and characterization of bovine brain type I phosphatidylinositol kinaseEur J Biochem199019137617672167854
- EngströmMKarlssonRJönssonJIInactivation of the forkhead transcription factor FoxO3 is essential for PKB-mediated survival of hematopoietic progenitor cells by kit ligandExp Hematol200331431632312691919
- MöllerCAlfredssonJEngströmMStem cell factor promotes mast cell survival via inactivation of FOXO3a-mediated transcriptional induction and MEK-regulated phosphorylation of the proapoptotic protein BimBlood200510641330133615855272
- MöllerCAlfredssonJEngströmMStem cell factor promotes mast cell survival via inactivation of FOXO3a-mediated transcriptional induction and MEK-regulated phosphorylation of the proapoptotic protein BimBlood200510641330133615855272
- VossellerKStellaGYeeNBesmerPc-Kit receptor signaling through its phosphatidylinositide-3′-kinase-binding site and protein kinase C: role in mast cell enhancement of degranulation, adhesion, and membrane rufflingMol Biol Cell1997859099229168474
- HashimotoKMatsumuraITsujimuraTNecessity of tyrosine 719 and phosphatidylinositol 3′-kinase–mediated signal pathway in constitutive activation and oncogenic potential of c-kit receptor tyrosine kinase with the Asp814Val mutationBlood200310131094110212393643
- ShivakrupaRBernsteinAWatringNLinnekinDPhosphatidylinositol 3′-kinase is required for growth of mast cells expressing the kit catalytic domain mutantCancer Res200363154412441912907613
- ArcaroAKhanzadaUKVanhaesebroeckBTetleyTDWaterfieldMDSecklMJTwo distinct phosphoinositide 3-kinases mediate polypeptide growth factor-stimulated PKB activationEMBO J200221195097510812356726
- BenesCHWuNEliaAEDhariaTCantleyLCSoltoffSPThe C2 domain of PKCδ is a phosphotyrosine binding domainCell2005121227128015851033
- BromannPAKorkayaHCourtneidgeSAThe interplay between Src family kinases and receptor tyrosine kinasesOncogene200423487957796815489913
- LinnekinDDeBerryCSMouSLyn associates with the juxtamembrane region of c-Kit and is activated by stem cell factor in hematopoietic cell lines and normal progenitor cellsJ Biol Chem19972724327450274559341198
- KrystalGWDeBerryCSLinnekinDLitzJLck associates with and is activated by Kit in a small cell lung cancer cell line: inhibition of SCF-mediated growth by the Src family kinase inhibitor PP1Cancer Res19985820466046669788619
- BroudyVCLinNLLilesWCSignaling via Src family kinases is required for normal internalization of the receptor c-KitBlood19999461979198610477727
- ShivakrupaRLinnekinDLyn contributes to regulation of multiple Kit-dependent signaling pathways in murine bone marrow mast cellsCell Signal200517110310915451030
- HongLMunugalavadlaVKapurRc-Kit-mediated overlapping and unique functional and biochemical outcomes via diverse signaling pathwaysMol Cell Biol20042431401141014729982
- WollbergPLennartssonJGottfridssonEYoshimuraARonnstrandLThe adapter protein APS associates with the multifunctional docking sites Tyr-568 and Tyr-936 in c-KitBiochem J20033701033103812444928
- KozlowskiMLaroseLLeeFLeDMRottapelRSiminovitchKASHP-1 binds and negatively modulates the c-Kit receptor by interaction with tyrosine 569 in the c-Kit juxtamembrane domainMol Cell Biol1998184208920999528781
- AgostiVCorbaciogluSEhlersICritical role for Kit-mediated Src kinase but not PI 3-kinase signaling in pro T and pro B cell developmentJ Exp Med2004199686787815024050
- MurphyLOSmithSChenRHFingarDCBlenisJMolecular interpretation of ERK signal duration by immediate early gene productsNat Cell Biol20024855656412134156
- ThommesKLennartssonJCarlbergMRonnstrandLIdentification of Tyr-703 and Tyr-936 as the primary association sites for Grb2 and Grb7 in the c-Kit/stem cell factor receptorBiochem J199934121121610377264
- HosseinimehrSJInanamiOHamasuTActivation of c-kit by stem cell factor induces radioresistance to apoptosis through ERK-dependent expression of survivin in HL60 cellsJ Radiat Res200445455756115635266
- LennartssonJWernstedtCEngströmUHellmanURönnstrandLIdentification of Tyr900 in the kinase domain of c-Kit as a Src-dependent phosphorylation site mediating interaction with c-CrkExp Cell Res2003288111011812878163
- BrizziMFZiniMGAronicaMGBlechmanJMYardenYPegoraroLConvergence of signaling by interleukin-3, granulocyte-macrophage colony-stimulating factor, and mast cell growth factor on JAK2 tyrosine kinaseJ Biol Chem19942695031680316847527392
- Pérez-LosadaJSánchez-MartıńMRodríguez-GarcıáAZinc-finger transcription factor Slug contributes to the function of the stem cell factor c-kit signaling pathwayBlood200210041274128612149208
- LennartssonJRonnstrandLThe stem cell factor receptor/c-Kit as a drug target in cancerCurr Cancer Drug Targets200661657516475976
- DeiningerMBuchdungerEDrukerBJThe development of imatinib as a therapeutic agent for chronic myeloid leukemiaBlood200510572640265315618470
- WakitaSYamaguchiHMiyakeKImportance of c-kit mutation detection method sensitivity in prognostic analyses of t (8; 21)(q22; q22) acute myeloid leukemiaLeukemia20112591423143221606963
- Willmore-PayneCLayfieldLJHoldenJAc-Kit mutation analysis for diagnosis of gastrointestinal stromal tumors in fine needle aspiration specimensCancer Cytopathol20051053165170
- FrostMJFerraoPTHughesTPAshmanLKJuxtamembrane mutant V560GKit is more sensitive to Imatinib (STI571) compared with wild-type c-Kit whereas the kinase domain mutant D816VKit is resistantMol Cancer Ther20021121115112412481435
- RobertsKGOdellAFByrnesEMResistance to c-KIT kinase inhibitors conferred by V654A mutationMol Cancer Ther2007631159116617363509
- KrügerSSotlarKKauschIHornyHPExpression of KIT (CD117) in renal cell carcinoma and renal oncocytomaOncology2005682–326927516015044
- CorbinASDemehriSGriswoldIJIn vitro and in vivo activity of ATP-based kinase inhibitors AP23464 and AP23848 against activation-loop mutants of KitBlood2005106122723415746079
- WangYYZhouGBYinTAML1-ETO and c-KIT mutation/overexpression in t (8; 21) leukemia: implication in stepwise leukemogenesis and response to GleevecProc Natl Acad Sci U S A200510241104110915650049
- ChanIKasprowiczSTharpMDistinct signalling pathways for mutated KIT (V560G) and KIT (D816V) in mastocytosisClin Exp Dermatol201338553854423777495
- LanternierFCohen-AkenineAPalmeriniFPhenotypic and genotypic characteristics of mastocytosis according to the age of onsetPLoS One200834e190618404201
- SchnittgerSKohlTMHaferlachTKIT-D816 mutations in AML1-ETO-positive AML are associated with impaired event-free and overall survivalBlood200610751791179916254134
- SakumaYSakuraiSOguniSHironakaMSaltoKAlterations of the c-kit gene in testicular germ cell tumorsCancer Sci200394648649112824871
- OrfaoAGarcia-MonteroACSanchezLEscribanoLRecent advances in the understanding of mastocytosis: the role of KIT mutationsBr J Haematol20071381123017555444
- NakataYKimuraAKatohOc-Kit point mutation of extracellular domain in patients with myeloproliferative disordersBr J Haematol19959136616638555071
- KimuraANakataYKatohOHyodoHc-Kit point mutation in patients with myeloproliferative disordersLeuk Lymphoma1997253–42812879168438
- ScheijenBGriffinJDTyrosine kinase oncogenes in normal hematopoiesis and hematological diseaseOncogene200221213314333312032772
- MakiRKeedyV2012KIT Exon 14 Mutation in GIST [webpage on the Internet]My Cancer Genome Available from: https://www.mycancergenome.org/content/disease/gist/kit/50/Accessed April 21, 2016
- HeinrichMCGriffithDJDrukerBJWaitCLOttKAZiglerAJInhibition of c-kit receptor tyrosine kinase activity by STI 571, a selective tyrosine kinase inhibitorBlood200096392593210910906
- TuvesonDAWillisNAJacksTSTI 571 inactivation of the gastrointestinal stromal tumor c-KIT oncoprotein: biological and clinical implicationsOncogene200120365054505811526490
- AttoubSRivatCRodriguesSThe c-kit tyrosine kinase inhibitor STI571 for colorectal cancer therapyCancer Res200262174879488312208734
- WeisbergEBoultonCKellyLMInhibition of mutant FLT3 receptors in leukemia cells by the small molecule tyrosine kinase inhibitor PKC412Cancer Cell20021543344312124173
- WheelerSSeegmillerAVnencak-JonesA2013KIT c.2447A>T (D816V) Mutation in Acute Myeloid Leukemia [webpage on the Internet]My Cancer Genome Available from: https://www.mycancergenome.org/content/disease/acute-myeloid-leukemia/kit/275/Accessed April 21, 2016
- YeeKWO’FarrellAMSmolichBDSU5416 and SU5614 inhibit kinase activity of wild-type and mutant FLT3 receptor tyrosine kinaseBlood200210082941294912351406
- SmolichBDYuenHAWestKAGilesFJAlbitarMCherringtonJMThe antiangiogenic protein kinase inhibitors SU5416 and SU6668 inhibit the SCF receptor (c-kit) in a human myeloid leukemia cell line and in acute myeloid leukemia blastsBlood20019751413142111222388
- KimDWJoYSJungHSAn orally administered multitarget tyrosine kinase inhibitor, SU11248, is a novel potent inhibitor of thyroid oncogenic RET/papillary thyroid cancer kinasesJ Clin Endocrinol Metabol2006911040704076
- Von BubnoffNGorantlaSKanchaRLordickFPeschelCDuysterJSpotlight correspondenceLeukemia2005191670167116015383
- LovlyCPaoWSosmanJ2015KIT c.2446G>C (D816H) Mutation in Melanoma [webpage on the Internet]My Cancer Genome Available from: https://www.mycancergenome.org/content/disease/melanoma/kit/84/Accessed April 21, 2016
- SambolEBAmbrosiniGGehaRCFlavopiridol targets c-KIT transcription and induces apoptosis in gastrointestinal stromal tumor cellsCancer Res200666115858586616740725
- BelloneGSilvestriSArtusioEGrowth stimulation of colorectal carcinoma cells via the c-kit receptor is inhibited by TGF-β1J Cell Physiol199717211119207920
- CurtinJABusamKPinkelDBastianBCSomatic activation of KIT in distinct subtypes of melanomaJ Clin Oncol200624264340434616908931
- TerheydenPHoubenRPajouhPThornsCZillikensDBeckerJCResponse to imatinib mesylate depends on the presence of the V559A-mutated KIT oncogeneJ Investig Dermatol2010130131431619812602
- WoodmanSETrentJCStemke-HaleKActivity of dasatinib against L576P KIT mutant melanoma: molecular, cellular, and clinical correlatesMol Cancer Ther2009882079208519671763
- Quintás-CardamaALazarAJWoodmanSEKimKRossMHwuPComplete response of stage IV anal mucosal melanoma expressing KIT Val560Asp to the multikinase inhibitor sorafenibNat Clin Pract Oncol200851273774018936790
- CarvajalRDAntonescuCRWolchokJDKIT as a therapeutic target in metastatic melanomaJAMA2011305222327233421642685
- HaradaHHaradaYNiimiHKyoTKimuraAInabaTHigh incidence of somatic mutations in the AML1/RUNX1 gene in myelodysplastic syndrome and low blast percentage myeloid leukemia with myelodysplasiaBlood200410362316232414615365
- GuoJSiLKongYPhase II, open-label, single-arm trial of imatinib mesylate in patients with metastatic melanoma harboring c-Kit mutation or amplificationJ Clin Oncol201129212904290921690468
- GirardN2015KIT c.2460T>A (D820E) Mutation in Thymic Carcinoma [webpage on the Internet]My Cancer Genome Available from: https://www.mycancergenome.org/content/disease/thymic-carcinoma/kit/120/Accessed April 21, 2016
- ButiSDoniniMSergioPImpressive response with imatinib in a heavily pretreated patient with metastatic c-KIT mutated thymic carcinomaJ Clin Oncol20112933e803e80521969494
- SchirosiLNanniniNNicoliDActivating c-KIT mutations in a subset of thymic carcinoma and response to different c-KIT inhibitorsAnn Oncol20122392409241422357254
- AntonescuCRSommerGSarranLAssociation of KIT exon 9 mutations with nongastric primary site and aggressive behavior KIT mutation analysis and clinical correlates of 120 gastrointestinal stromal tumorsClin Cancer Res2003993329333712960119
- RossiFYozgatYde StanchinaEImatinib upregulates compensatory integrin signaling in a mouse model of gastrointestinal stromal tumor and is more effective when combined with dasatinibMol Cancer Res2010891271128320736294
- StröbelPHartmannMJakobAThymic carcinoma with overexpression of mutated KIT and the response to imatinibN Engl J Med2004350252625262615201427
- AntonescuCRBesmerPGuoTAcquired resistance to imatinib in gastrointestinal stromal tumor occurs through secondary gene mutationClin Cancer Res200511114182419015930355
- YohKNishiwakiYIshiiGMutational status of EGFR and KIT in thymoma and thymic carcinomaLung Cancer200862331632018448188
- DişelUÖztuzcuSBeşenAAPromising efficacy of sorafenib in a relapsed thymic carcinoma with c-KIT exon 11 deletion mutationLung Cancer201171110911220970876
- GuoTAgaramNPWongGCSorafenib inhibits the imatinib-resistant KITT670I gatekeeper mutation in gastrointestinal stromal tumorClin Cancer Res200713164874488117699867
- TimokhinaIKisselHStellaGBesmerPKit signaling through PI 3-kinase and Src kinase pathways: an essential role for Rac1 and JNK activation in mast cell proliferationEMBO J19981721625062629799234
- GoddardNMcIntyreASummersgillBGilbertDKitazawaSShipleyJKIT and RAS signalling pathways in testicular germ cell tumours: new data and a review of the literatureInt J Androl200730433734917573850
- WandziochEEdlingCEPalmerRHCarlssonLHallbergBActivation of the MAP kinase pathway by c-Kit is PI-3 kinase dependent in hematopoietic progenitor/stem cell linesBlood20041041515714996702