
Abstract
Background
Blastocystis species (sp.) are gastrointestinal protozoan parasites with high prevalence rates worldwide. Blastocystis sp. show extensive genetic diversity with 17 different subtypes (STs) described to date. A few studies have investigated the prevalence and STs of Blastocystis sp. in Makkah, Saudi Arabia. Therefore, we aimed in this study to identify and characterize subtypes of Blastocystis sp. in the City of Makkah, Saudi Arabia.
Methods
Stool samples were collected from 140 patients who presented to King Abdulaziz Hospital, Hera General Hospital and Modern Medical Center in Saudi Arabia. Different microscopic examination methods of patients’ stools and molecular analyses (using primers targeting SSU rRNA gene) were performed to identify and characterize STs of Blastocystis sp.
Results
Our microscopic examination of stool samples showed that 96/140 patients (68.6%) had Blastocystis sp. infection. Clinical examination of infected patients revealed that 81 patients were symptomatic, whereas 15 were asymptomatic. Next, we isolated DNA from Blastocystis sp.-positive stool samples followed by PCR amplification of small-subunit ribosomal RNA (SSU rRNA) gene and sequence analysis. Our sequence analysis showed that subtype 3 (ST3) was the most prevalent (53.13%) followed by subtype 1 (ST1) (45.83%), whereas subtype 2 (ST2) was the least prevalent (1.04%). Moreover, our results showed that all three STs resulted in more symptomatic than asymptomatic cases. Finally, we identified novel haplotypes which comprised of 8 ST3, 6 ST1, and one ST2 haplotypes.
Conclusion
Our identification of several haplotypes in patients’ stools confirms the genetic diversity of Blastocystis sp. and may explain the reported low host specificity and differential pathogenicity of Blastocystis sp. We believe that additional molecular epidemiological and genomic studies are needed to understand the prevalence and pathogenicity of different subtypes in humans and animal hosts.
Background
Blastocystis species (sp.) are intestinal protozoan parasites that have several forms in lifecycle including vacuolar, granular, amoeboid, and cyst forms. Human to human transmission is common, through the fecal-oral route, which explains significantly higher rates of infection among food and animal handlersCitation1,Citation2.
According to previous reports, developing countries have higher prevalence rates (55–70%) of Blastocystis sp. compared to developed countries (10–15%). This could be attributed to higher standards of hygiene, lesser contact with animals and lower consumption rates of contaminated food and drink in developed countries.Citation3,Citation4 Recent studies have suggested that Blastocystis sp. exist as part of gut microbiota in healthy individuals (asymptomatic) and in patients presenting with gastrointestinal disease (symptomatic). However, the pathogenicity of the parasite, factors and mechanisms that trigger the progression to disease remain poorly understood.Citation5,Citation6 Symptomatic patients may present with nonspecific gastrointestinal symptoms such as abdominal pain, vomiting, and diarrhea. However, irritable bowel syndrome and urticaria sensitization were also reported.Citation7,Citation8 Several genomic, in vitro, and in vivo studies have identified multiple virulence factors of Blastocystis sp. and demonstrated the parasite’s pathological effects on the intestine.Citation6,Citation9
By 2013, Blastocystis sp. have been classified into 17 subtypes (STs) (ST1-17) in birds and mammals (including humans) based on extensive genetic variations in small-subunit ribosomal RNA (SSU rRNA) gene sequence.Citation10 Eleven additional STs (ST18-28) have been described in literature since then; however, four of these STs (ST18-20, and ST22) are still being validated.Citation11 These different STs are believed to result in differential symptoms and distribution among animal hosts and humans.Citation12 Ten STs have been isolated from human stools (ST1-9 and ST12) and more than 90% of all human cases have been associated with ST1-4.Citation13,Citation14 However, only eight subtypes (ST10-17) were isolated from animal hosts.Citation12,Citation15–17 A study has shown that Blastocystis sp. ST3, isolated primarily from symptomatic patients, produces proteases that play a major role in host protein degradation and immune evasion.Citation4,Citation18
Several molecular methods have been used to identify Blastocystis sp. subtypes that are isolated from humans and animals. These methods include random amplified polymorphic DNA (RAPD) using four different arbitrary polymerase chain reaction (PCR) primers, and restriction fragment length polymorphism (RFLP) followed by PCR to detect SSU rRNA gene variations.Citation19,Citation20 To date, a few studies investigated the prevalence and subtypes of B. hominis in Saudi Arabia. Therefore, the aim of this study was to identify Blastocystis sp. subtypes and their prevalence in stool samples of patients living in City of Makkah, Saudi Arabia.
Methods
Study Population and Sample Collection
The present study involved 140 patients who presented, in 2017–2018, to King Abdulaziz Hospital, Hera General Hospital, and Modern Medical Center in the City of Makkah, Saudi Arabia. Some of the participants were complaining of GIT symptoms such as diarrhea or abdominal pain, while others were asymptomatic. Stool samples were collected from each patient in a clean container and divided into 2 parts. The first part was examined microscopically for the presence of Blastocystis sp. using direct mount (saline and iodine) and concentration technique; as previously described.Citation21 The second part was stored at −20°C for further molecular analysis.
PCR Amplification and Sequencing
Stool samples that tested positive for Blastocystis sp. were subjected to molecular analysis. DNA was extracted from stool samples using QIAamp DNA Stool Mini Kit (QIAGEN) according to the manufacturer’s instructions. The specific primers, forward primer Blast 505–532 (5′ GGA GGT AGT GAC AAT AAATC 3′) and reverse primer Blast 998–1017 (5′ TGC TTT CGC ACT TGT TCATC 3′),Citation22 were used to amplify the SSU rRNA gene using AccuPower® PCR PreMix (Bioneer). The PCR conditions were as follows: initial denaturation at 95°C for 4 min then 35 cycles of {denaturation at 95°C for 30 seconds, annealing at 54°C for 30 seconds, extension at 72°C for 30 seconds}, and a final extension step at 72°C for 5 minutes. Next, PCR products were separated by electrophoresis on 1.5% agarose gels and visualized by UV following ethidium bromide staining. Positive Blastocystis sp. samples generated PCR products of 500bp. For characterization of Blastocystis sp. genetic variations, all PCR products as well as positive and negative controls were sent to Macrogen company in South Korea for sequencing (https://dna.macrogen.com/).
Sequence Analysis
The DNA sequences of PCR products generated from Blastocystis sp.-positive samples were analyzed. The BLASTN software was used to compare the SSU rRNA sequences obtained with those in GenBank. MEGA X software was used to build a phylogenetic tree using the neighbor-joining method and the Kimura 2-parameter model.Citation23 The bootstrap method with 1000 replicates was used to determine the support of monophyletic communities. All sequences were aligned by MUSCLE software and phylogenetic analysis was performed using MEGA X the maximum likelihood (ML), and Neighbor Joining (NJ) method. The maximum parsimony (MP) methods with 1000 bootstrap replicates were then used to construct the phylogenetic tree. The evolutionary history was concluded using the NJ method.Citation24 The Maximum Composite Likelihood method was used to calculate the evolutionary distancesCitation25 and are in the units of the number of base substitutions per site. The analysis included 46 nucleotide sequences and any equivocal position was deleted for each sequence pair (pairwise deletion option). There was a total of 48,775 positions in the final dataset. MEGA X software was used for evolutionary analyses.Citation23
Statistical Analysis
All collected data were entered into the SPSS program (version 22) and chi-square test was used to analyze categorical variables. P-values of <0.05 were considered statistically significant.
Results
Microscopic Examination of Patients’ Stools Identified 96 Blastocystis sp.-Infected Patients with Significantly Higher Number of Symptomatic Than Asymptomatic Patients
The present study included 140 patients aging between 7 and 80 years (mean ± SD = 35.5±12.4) of which 105 (75%) were males (). Microscopic examination of stool samples of all patients showed that 96 patients (96/140; 68.6%) were infected with Blastocystis sp. Our results showed that there was a significantly higher percentage of males infected with Blastocystis sp. (N=77/105; 73.3%) compared to females (N=19/35; 54.3%) (; p<0.05). Interestingly, age group 21–40 years (yrs) had the highest percentage of infected patients (70/94; 74.5%) compared to age group <20 yrs (1/4; 25%), age group 41–60 yrs (23/37; 62.2%), and age group >60 yrs (2/5; 40%), . However, the differences were not statistically significant (p=0.3).
Table 1 Gender and Age Group of Blastocystis sp.-Infected and Uninfected Patients
Physical examination of Blastocystis sp.-infected patients concluded that a significantly higher number were symptomatic (diarrhea) than asymptomatic (81 vs 15; p< 0.05).
Sequence Analysis of PCR Products Amplified from DNA of Blastocystis sp., Isolated from Stools of Infected Patients, Showed Prevalence of Blastocystis sp. Subtype 3 (ST3)
Our PCR on the DNA extracted from the 96 Blastocystis sp.-positive stool samples generated PCR products of the right size (500 bp) from all samples, which confirmed Blastocystis sp. infection.
Next, we sequenced PCR products to identify the genetic subtype of Blastocystis sp. in each sample. The obtained sequences were analyzed using BLAST software and aligned with Blastocystis sp. sequences deposited in the gene bank. Sequence analysis showed that ST3 existed in significantly higher percentage of stool samples compared to ST1 (53.13% vs 45.83%). ST2 was detected in only 1.04% of samples (p<0.05).
Our results showed that 100% of ST2-infected patients were symptomatic followed by ST1 (88.6%), and ST3 (80.4%).
Novel Blastocystis sp. Haplotypes Were Identified in Subtypes with Inter- and Intra-Subtype Genetic Variations
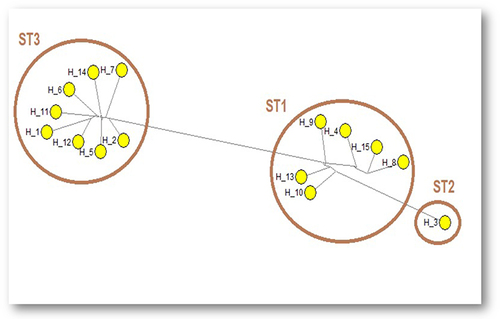
Median joining network analysis, of sequences of PCR products, demonstrated genetic variations between the different subtypes, and identified intra-subtype genetic variations within ST1 and ST3 (). The median joining network analysis identified 15 haplotypes; H1-15, in Blastocystis sp. subtypes. ST3 comprised 8 haplotypes, ST1 comprised 6 haplotypes, whereas ST2 comprised only one haplotype.
Figure 1 Median joining network sequence analysis of SSU rRNA gene-PCR products demonstrated different haplotypes (H1-15) within Blastocystis sp. subtypes. Maximum parsimony (MP) analysis was used as the optional post-processing calculation using Network 4.6.1.3 software. Each haplotype represents a distinct subtype. The haplotypes are numbered from H1 to H15.
Further sequence analysis of PCR products revealed inter-subtype genetic variations with genetic differentiation values ranging from a minimum of JC = 0.0991, between ST1 and ST2 subtypes, to higher values between ST1 and ST3 (JC = 0.248) (). Moreover, high intra-subtype genetic variations were found between ST1 sequences (JC = 0.0834) followed by ST3 (JC = 0.0356) ().
Table 2 Inter- and Intra-Subtype Genetic Variations Characterize Blastocystis sp. Subtypes/Haplotypes
Higher Genetic Variations Were Detected Between ST1 Haplotypes Compared to ST3 Haplotypes
Next, we performed an overall analysis of sequences of SSU rRNA gene PCR products, obtained from all Blastocystis sp.-positive stool samples. We identified 113 sites that were polymorphic and 91 sites that were parsimony informative, which led to the distinction of 15 haplotypes. The total number of non-InDel and InDel sites were 431 and 25, respectively. The 91 parsimony informative sites included 59 sites that had two variants, 24 sites that had three variants, and 8 sites with four variants. The average number of nucleotide differences was 52.076. We found that the haplotype and nucleotide diversity were 1.000 and 0.12083, respectively.
Consistent with the data shown in , the analysis of haplotypes of each subtype confirmed that ST1 has higher genetic variations than ST3, with higher nucleotide diversity, higher numbers of polymorphic sites and higher number of nucleotide differences (). All subtypes showed negative neutrality tests (Tajima’s test and Fu’s Fs) which means that DNA sequence is evolving naturally and not under directional selection.
Table 3 Genetic Variability of Blastocystis sp. Haplotypes
Next, we constructed the phylogenetic tree and included all our identified haplotypes. We determined the relation of the identified haplotypes to other Blastocystis sp. isolates (). We used thirty reference sequences of Blastocystis sp. subtypes ST1-ST17 () and Proteromonas lacertae (U37108) as an outgroup sequence.
Table 4 GenBank References for the Blastocystis sp. Subtypes Used to Construct the Phylogenetic Tree
Figure 2 Phylogenetic relationship of Blastocystis sp. haplotypes identified in the current study to other Blastocystis sp. isolates, based on SSU rRNA gene sequences. The reference GenBank sequence accession numbers of all isolates are shown, including our haplotypes accession numbers in bold. Proteromonas lacertae served as the outgroup sequence. *Indicates the haplotypes identified in the current study.

Discussion
Blastocystis sp. infections are prevalent in both developing and developed countries. Multiple studies have described several gastrointestinal manifestations following Blastocystis sp. infections.Citation26 However, several studies have shown that only a few subtypes of Blastocystis sp. are associated with diseases in humans, whereas other subtypes cause asymptomatic infections.
In the present study, we aimed to identify and characterize Blastocystis sp. subtypes in 140 patients who attended King Abdulaziz Hospital, Hera General Hospital and Modern Medical Center in City of Makkah, Saudi Arabia. We detected Blastocystis sp. infection in 68.6% (96/140) patients who presented with gastrointestinal symptoms (symptomatic) or remained asymptomatic. Other studies from different countries have shown variable prevalence rates of 0.54% in Iran,Citation27 2.5% in Japan,Citation28 3.3% in Singapore,Citation29 19% in Lebanon,Citation17 22.1% in Libya,Citation30 33.3% in Egypt,Citation31 40.7% in the Philippines,Citation32 and 46.9% in Venezuela.Citation33 This could be attributed to variations in sanitation standards in different countries. Our small number of patients could be the reason for the higher prevalence rates of Blastocystis sp. in our study compared to other studies.
We report higher prevalence rate of Blastocystis sp. in males compared to females, which is consistent with the findings of previous studies in Iran, Libya and Turkey.Citation34–36 The higher prevalence rate of Blastocystis sp. in males compared to females may be attributed to their more frequent outdoor activities. There are several contradictory studies regarding Blastocystis sp. pathogenicity.Citation37 In the present study, we report that the number of symptomatic Blastocystis sp. -infected patients were significantly higher than the number of asymptomatic patients (81 symptomatic patients vs 15 asymptomatic).
Nine subtypes of Blastocystis sp. (ST1 to ST9) were isolated from human stoolsCitation38 of which ST1, ST2, ST3, ST4, and ST5 were the most common. It has been shown that ST4 is the prevalent subtype in European countries, while ST1, ST2, ST3, and ST5 are the common subtypes in other countries all over the world.Citation39 However, there is still debate about the relationship between Blastocystis sp. subtypes and clinical symptoms.Citation40–42
Our molecular analysis, of patients’ stool samples, showed that ST3 was detected in significantly higher percentage of samples than ST1 (53.13% vs 45.83%). We identified ST2 in only one sample. This finding indicates that ST3 is the predominant subtype of Blastocystis sp. in our patients. Predominance of ST3, in our study, is consistent with a previous reportCitation43 which characterized Blastocystis sp. subtypes in symptomatic (diarrhea) and asymptomatic patients in City of Makkah, Saudi Arabia. Moreover, ST3 was found to be the prevalent subtype (41–92%) in a comparative study between Japan, Bangladesh, Pakistan and Germany,Citation28 whereas the prevalence rates in other studies were reported at 78%, 75.9%, 54.5%, 53.5% and 33.3% in Singapore, Turkey, Egypt, France and Lebanon, respectively.Citation17,Citation29,Citation36,Citation44,Citation45
We additionally showed that a significantly higher percentage of ST3-infected patients were symptomatic than asymptomatic (80.4% vs 19.6%). This finding is in agreement with findings of another study where most ST3 patients showed gastrointestinal symptoms.Citation5 Tan et al also showed similar results and reported that all symptomatic patients in his study were ST3-infected.Citation46 Symptoms other than gastrointestinal manifestations were also reported in Blastocystis sp. ST3-infected patients. A patient infected with ST3 presented with acute urticaria and gastrointestinal symptoms.Citation47 We also showed that most ST1-infected patients (88.6%) were symptomatic which is in agreement with other studies which reported the ability of ST1 to cause symptomatic infections.Citation28,Citation48 This indicates that Blastocystis sp. STs, that were identified in our study, tend to cause symptomatic than asymptomatic infections.
Our sequence analysis, of PCR products obtained from all samples, identified three parsimony informative sites and three mutations (not parsimony informative) across the entire obtained sequences, resulting in the identification of 15 Blastocystis sp. haplotypes. (8 different ST3 haplotypes, 6 ST1 haplotypes, and one ST2 haplotype). The sequence analysis of ST1 and ST3 samples revealed high genetic variations. This high level of genetic variability in ST1 and ST3 appears to support t’s low host specificity.Citation49 Despite our findings, we believe that large-scale molecular epidemiological studies, both in humans and in animals, will be useful to further understand the transmission of Blastocystis sp. Moreover, immunological and pathogenicity studies should be performed for better understanding of the Blastocystis sp. pathogenicity and immune responses in humans.
Several studies have isolated ST1 and ST3 from a variety of species including monkeys, goats, pigs, dogs, and non-human primates. As a result, it has been proposed that these subtypes are zoonotic subtypes, capable of infecting humans at varying rates.Citation48,Citation50–52 A study analyzed both the nuclear SSU rRNA and the MLST of the mitochondrion-like organelle genome and discovered significant intra-subtype genetic variation in ST3. The fact that ST3 has high intra-subtype variation means that this subtype may have co-evolved with human hosts over a long time.Citation53
Conclusions
A few studies investigated the prevalence and subtypes of Blastocystis sp. STs in patients from Saudi Arabia. Our study showed that 81/96 Blastocystis sp.-infected patients from Saudi Arabia were symptomatic, whereas stool analysis identified 15 genetically distinct haplotypes that are distributed in three STs (8 ST3, 6 ST1, and 1 ST2). We believe that future genomic analysis could shed light on potential coevolutionary aspects, which could help researchers better understand Blastocystis sp. Biology, interaction with the host, and pathogenicity. Furthermore, proteomic studies of the various subtypes of Blastocystis sp. could enhance our understanding of the parasite’s pathogenic mechanisms.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the ethics and research committee, Faculty of Applied Medical Sciences, King Abdulaziz University (Protocol number: FAMS-ERC2017-12). An informed consent was obtained from each patient involved in this study. The Saudi Ministry of Health, Saudi Arabia cooperated in this study.
Data Sharing Statement
All genetic sequences are available at [https://www.ncbi.nlm.nih.gov/genbank/]. GenBank accession numbers are: MZ396562, MZ396563, MZ396564, MZ396565, MZ396566, MZ396567, MZ396568, MZ396569, MZ396570, MZ396571, MZ396572, MZ396573, MZ396574, MZ396575, MZ396576.
Disclosure
The authors declare no competing interests in this work.
References
- Wakid M, Azhar E, Zafar T. Intestinal parasitic infection among food handlers in the holy city of Makkah During Hajj season1428 Hegira (2007G). J King Abdulaziz Univ Sci. 2009;16(1):39–52. doi:10.4197/med.16-1.4
- Aldahhasi W, Toulah F, Wakid M. Evaluation of common microscopic techniques for detection of Blastocystis hominis. J Egypt Soc Parasitol. 2020;50(1):33–40. doi:10.21608/jesp.2020.88748
- Belkhair J, Karrati I, Tarmidi M, El Mezouari M, Moutaj R. Blastocystis hominis microbiota: study of 13255 patients and review of the literature. J Microbiol Exp. 2021;9(2):29–32. doi:10.15406/jmen.2021.09.00319
- Palasuwan A, Palasuwan D, Mahittikorn A, Chiabchalard R, Combes V, Popruk S. Subtype distribution of Blastocystis in communities along the Chao Phraya River, Thailand. Korean J Parasitol. 2016;54(4):455–460. doi:10.3347/kjp.2016.54.4.455
- Khademvatan S, Masjedizadeh R, Yousefi-Razin E, et al. PCR-based molecular characterization of Blastocystis hominis subtypes in southwest of Iran. J Infect Public Health. 2018;11(1):43–47. doi:10.1016/j.jiph.2017.03.009
- Ajjampur SSR, Tan KSW. Pathogenic mechanisms in Blastocystis spp. - Interpreting results from in vitro and in vivo studies. Parasitol Int. 2016;65(6Pt B):772–779. doi:10.1016/j.parint.2016.05.007
- Kantardjiev V, Galev A, Broshtilova V. Urticaria associated with amoeboid forms of Blastocystis spp. Asian J Res Infect Dis. 2019;2(3):1–4. doi:10.9734/ajrid/2019/v2i330105
- Lepczyńska M, Dzika E, Kubiak K, Korycińska J. The role of Blastocystis sp. as an etiology of irritable bowel syndrome. Polish Ann Med. 2016;23:57–60. doi:10.1016/j.poamed.2015.04.001
- Wu Z, Mirza H, Tan KSW. Intra-subtype variation in enteroadhesion accounts for differences in epithelial barrier disruption and is associated with metronidazole resistance in Blastocystis subtype-7. PLoS Negl Trop Dis. 2014;8(5):e2885. doi:10.1371/journal.pntd.0002885
- Stensvold CR, Lebbad M, Hansen A, et al. Differentiation of Blastocystis and parasitic archamoebids encountered in untreated wastewater samples by amplicon-based next-generation sequencing. Parasite Epidemiol Control. 2020;9:e00131. doi:10.1016/j.parepi.2019.e00131
- Hublin JSY, Maloney JG, Santin M. Blastocystis in domesticated and wild mammals and birds. Res Vet Sci. 2021;135:260–282. doi:10.1016/j.rvsc.2020.09.031
- Alfellani MA, Taner-Mulla D, Jacob AS, et al. Genetic diversity of Blastocystis in livestock and zoo animals. Protist. 2013;164(4):497–509. doi:10.1016/j.protis.2013.05.003
- Stensvold CR, Tan KSW, Clark CG. Blastocystis. Trends Parasitol. 2020;36(3):315–316. doi:10.1016/j.pt.2019.12.008
- Stensvold CR, Clark CG. Molecular identification and subtype analysis of Blastocystis. Curr Protoc Microbiol. 2016;43(1):20A- 2.
- Zhu W, Tao W, Gong B, et al. First report of Blastocystis infections in cattle in China. Vet Parasitol. 2017;246:38–42. doi:10.1016/j.vetpar.2017.09.001
- Roberts T, Stark D, Harkness J, Ellis J. Subtype distribution of Blastocystis isolates identified in a Sydney population and pathogenic potential of Blastocystis. Eur J Clin Microbiol Infect Dis. 2013;32(3):335–343. doi:10.1007/s10096-012-1746-z
- El Safadi D, Meloni D, Poirier P, et al. Molecular epidemiology of Blastocystis in Lebanon and correlation between subtype 1 and gastrointestinal symptoms. Am J Trop Med Hyg. 2013;88(6):1203–1206. doi:10.4269/ajtmh.12-0777
- Abdel-Hameed DM, Hassanin OM. Proteaese activity of Blastocystis hominis subtype3 in symptomatic and asymptomatic patients. Parasitol Res. 2011;109(2):321–327. doi:10.1007/s00436-011-2259-x
- Motazedian H, Ghasemi H, Sadjjadi SM. Genomic diversity of Blastocystis hominis from patients in southern Iran. Ann Trop Med Parasitol. 2008;102(1):85–88. doi:10.1179/136485908X252197
- Sardarian K, Hajilooi M, Maghsood A, Moghimbeigi A, Alikhani M. A study of the genetic variability of Blastocystis hominis isolates in Hamadan, west of Iran. 2012.
- Manser MM, Saez ACS, Chiodini PL. Faecal parasitology: concentration methodology needs to be better standardised. PLoS Negl Trop Dis. 2016;10(4):e0004579. doi:10.1371/journal.pntd.0004579
- Böhm-Gloning B, Knobloch J, Walderich B. Five subgroups of Blastocystis hominis from symptomatic and asymptomatic patients revealed by restriction site analysis of PCR-amplified 16S-like rDNA. Trop Med Int Health. 1997;2(8):771–778. doi:10.1046/j.1365-3156.1997.d01-383.x
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35(6):1547–1549. doi:10.1093/molbev/msy096
- Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406–425. doi:10.1093/oxfordjournals.molbev.a040454
- Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci U S A. 2004;101(30):11030–11035. doi:10.1073/pnas.0404206101
- Duda A, Kosik-Bogacka D, Lanocha-Arendarczyk N, Kołodziejczyk L, Lanocha A. The prevalence of Blastocystis hominis and other protozoan parasites in soldiers returning from peacekeeping missions. Am J Trop Med Hyg. 2015;92(4):805–806. doi:10.4269/ajtmh.14-0344
- Riabi TR, Haghighi A, Mirjalali H, et al. Study of prevalence, distribution and clinical significance of Blastocystis isolated from two medical centers in Iran. Gastroenterol Hepatol Bed Bench. 2017;10:S102–S107. doi:10.22037/ghfbb.v0i0.1292
- Yoshikawa H, Wu Z, Kimata I, et al. Polymerase chain reaction-based genotype classification among human Blastocystis hominis populations isolated from different countries. Parasitol Res. 2004;92(1):22–29. doi:10.1007/s00436-003-0995-2
- Wong KHS, Ng GC, Lin RTP, Yoshikawa H, Taylor MB, Tan KSW. Predominance of subtype 3 among Blastocystis isolates from a major hospital in Singapore. Parasitol Res. 2008;102(4):663–670. doi:10.1007/s00436-007-0808-0
- Abdulsalam AM, Ithoi I, Al-Mekhlafi HM, Al-Mekhlafi AM, Ahmed A, Surin J. Subtype distribution of Blastocystis isolates in Sebha, Libya. PLoS One. 2013;8(12):e84372. doi:10.1371/journal.pone.0084372
- Rayan HZE, Ismail OA, El Gayar EK. Prevalence and clinical features of Dientamoeba fragilis infections in patients suspected to have intestinal parasitic infection. J Egypt Soc Parasitol. 2007;37(2):599–608.
- Baldo ET, Belizario VY, De Leon WU, Kong HH, Chung D-I. Infection status of intestinal parasites in children living in residential institutions in Metro Manila, the Philippines. Korean J Parasitol. 2004;42(2):67–70. doi:10.3347/kjp.2004.42.2.67
- Velásquez V, Caldera R, Wong W, et al. [Blastocystosis: a high prevalence of cases found in patients from Health Center of Soledad, Anzoategui State, Venezuela]. Rev Soc Bras Med Trop. 2005;38(4):356–357. Portuguese. doi:10.1590/s0037-86822005000400017
- Al-Fellani MA, Khan AH, Al-Gazoui RM, Zaid MK, Al-Ferjani MA. Prevalence and clinical features of Blastocystis hominis infection among patients in Sebha, Libya. Sultan Qaboos Univ Med J. 2007;7(1):35–40.
- Khoshnood S, Rafiei A, Saki J, Alizadeh K. Prevalence and genotype characterization of Blastocystis hominis among the Baghmalek people in southwestern Iran in 2013–2014. Jundishapur J Microbiol. 2015;8(10):4–8. doi:10.5812/jjm.23930
- Ozyurt M, Kurt O, Mølbak K, Nielsen HV, Haznedaroglu T, Stensvold CR. Molecular epidemiology of Blastocystis infections in Turkey. Parasitol Int. 2008;57(3):300–306. doi:10.1016/j.parint.2008.01.004
- Roberts T, Stark D, Harkness J, Ellis J. Update on the pathogenic potential and treatment options for Blastocystis sp. Gut Pathog. 2014;6:17. doi:10.1186/1757-4749-6-17
- Rene BA, Stensvold CR, Badsberg JH, Nielsen HV. Subtype analysis of Blastocystis isolates from Blastocystis cyst excreting patients. Am J Trop Med Hyg. 2009;80(4):588–592. doi:10.4269/ajtmh.2009.80.588
- Alfellani MA, Stensvold CR, Vidal-Lapiedra A, Onuoha ESU, Fagbenro-Beyioku AF, Clark CG. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013;126(1):11–18. doi:10.1016/j.actatropica.2012.12.011
- Rezaei Riabi T, Mirjalali H, Haghighi A, et al. Genetic diversity analysis of Blastocystis subtypes from both symptomatic and asymptomatic subjects using a barcoding region from the 18S rRNA gene. Infect Genet Evol. 2018;61:119–126. doi:10.1016/j.meegid.2018.03.026
- Moosavi A, Haghighi A, Mojarad EN, et al. Genetic variability of Blastocystis sp. isolated from symptomatic and asymptomatic individuals in Iran. Parasitol Res. 2012;111(6):2311–2315. doi:10.1007/s00436-012-3085-5
- Alinaghizade A, Mirjalali H, Mohebali M, Stensvold CR, Rezaeian M. Inter- and intra-subtype variation of Blastocystis subtypes isolated from diarrheic and non-diarrheic patients in Iran. Infect Genet Evol. 2017;50:77–82. doi:10.1016/j.meegid.2017.02.016
- Mohamed RT, El-Bali MA, Mohamed AA, et al. Subtyping of Blastocystis sp. isolated from symptomatic and asymptomatic individuals in Makkah, Saudi Arabia. Parasit Vectors. 2017;10(1):1–7. doi:10.1186/s13071-017-2114-8
- Souppart L, Sanciu G, Cian A, et al. Molecular epidemiology of human Blastocystis isolates in France. Parasitol Res. 2009;105(2):413–421. doi:10.1007/s00436-009-1398-9
- Hussein EM, Hussein AM, Eida MM, Atwa MM. Pathophysiological variability of different genotypes of human Blastocystis hominis Egyptian isolates in experimentally infected rats. Parasitol Res. 2008;102(5):853–860. doi:10.1007/s00436-007-0833-z
- Tan TC, Suresh KG, Smith HV. Phenotypic and genotypic characterisation of Blastocystis hominis isolates implicates subtype 3 as a subtype with pathogenic potential. Parasitol Res. 2008;104(1):85–93. doi:10.1007/s00436-008-1163-5
- Katsarou-Katsari A, Vassalos CM, Tzanetou K, Spanakos G, Papadopoulou C, Vakalis N. Acute urticaria associated with amoeboid forms of Blastocystis sp. subtype 3. Acta Derm Venereol. 2008;88(1):80–81. doi:10.2340/00015555-0338
- Yan Y, Su S, Ye J, et al. Blastocystis sp. subtype 5: a possibly zoonotic genotype. Parasitol Res. 2007;101(6):1527–1532. doi:10.1007/s00436-007-0672-y
- Alfellani MA, Jacob AS, Perea NO, et al. Diversity and distribution of Blastocystis sp. subtypes in non-human primates. Parasitology. 2013;140(8):966–971. doi:10.1017/S0031182013000255
- Lee IL, Tan TC, Tan PC, et al. Predominance of Blastocystis sp. subtype 4 in rural communities, Nepal. Parasitol Res. 2012;110(4):1553–1562. doi:10.1007/s00436-011-2665-0
- Stensvold CR, Lewis HC, Hammerum AM, et al. Blastocystis: unravelling potential risk factors and clinical significance of a common but neglected parasite. Epidemiol Infect. 2009;137(11):1655–1663. doi:10.1017/S0950268809002672
- Noël C, Dufernez F, Gerbod D, et al. Molecular phylogenies of Blastocystis isolates from different hosts: implications for genetic diversity, identification of species, and zoonosis. J Clin Microbiol. 2005;43(1):348–355. doi:10.1128/JCM.43.1.348-355.2005
- Scanlan PD, Stensvold CR. Blastocystis: getting to grips with our guileful guest. Trends Parasitol. 2013;29(11):523–529. doi:10.1016/j.pt.2013.08.006
- Pintong A, Sunyanusin S, Prasertbun R, et al. Blastocystis subtype 5: predominant subtype on pig farms, Thailand. Parasitol Int. 2018;67(6):824–828. doi:10.1016/j.parint.2018.08.009
- Arisue N, Hashimoto T, Yoshikawa H. Sequence heterogeneity of the small subunit ribosomal RNA genes among blastocystis isolates. Parasitology. 2003;126(Pt 1):1–9. doi:10.1017/s0031182002002640
- Rivera WL. Phylogenetic analysis of Blastocystis isolates from animal and human hosts in the Philippines. Vet Parasitol. 2008;156(3):178–182. doi:10.1016/j.vetpar.2008.06.001
- Thathaisong U, Siripattanapipong S, Mungthin M, et al. Identification of Blastocystis subtype 1 variants in the home for girls, Bangkok, Thailand. Am J Trop Med Hyg. 2013;88(2):352–358. doi:10.4269/ajtmh.2012.12-0237
- Leelayoova S, Siripattanapipong S, Thathaisong U, et al. Drinking water: a possible source of blastocystis spp. subtype 1 infection in schoolchildren of a rural community in central Thailand. Am J Trop Med Hyg. 2008;79(3):401–406. doi:10.4269/ajtmh.2008.79.401
- Abe N. Molecular and phylogenetic analysis of Blastocystis isolates from various hosts. Vet Parasitol. 2004;120(3):235–242. doi:10.1016/j.vetpar.2004.01.003
- Noël C, Peyronnet C, Gerbod D, et al. Phylogenetic analysis of Blastocystis isolates from different hosts based on the comparison of small-subunit rRNA gene sequences. Mol Biochem Parasitol. 2003;126(1):119–123. doi:10.1016/S0166-6851(02)00246-3
- Villegas-Gómez I, Martínez-Hernández F, Urrea-Quezada A, et al. Comparison of the genetic variability of Blastocystis subtypes between human carriers from two contrasting climatic regions of México. Infect Genet Evol. 2016;44:334–340. doi:10.1016/j.meegid.2016.07.036
- Stensvold CR, Traub RJ, von Samson-himmelstjerna G, Jespersgaard C, Nielsen HV, Thompson RCA. Blastocystis: subtyping isolates using pyrosequencing technology. Exp Parasitol. 2007;116(2):111–119. doi:10.1016/j.exppara.2006.12.002
- Bart A, Wentink-Bonnema EMS, Gilis H, et al. Diagnosis and subtype analysis of Blastocystis sp. in 442 patients in a hospital setting in the Netherlands. BMC Infect Dis. 2013;13(1):2–7. doi:10.1186/1471-2334-13-389
- Sharifi Y, Abbasi F, Shahabi S, Zaraei A, Mikaeili F, Sarkari B. Comparative genotyping of Blastocystis infecting cattle and human in the south of Iran. Comp Immunol Microbiol Infect Dis. 2020;72:101529. doi:10.1016/j.cimid.2020.101529