
Abstract
Background
Adoptive T cell therapy has been proven to be a promising modality for the treatment of cancer patients in recent years. However, the increased expression of inhibitory receptors could negatively regulate the function and persistence of transferred T cells which mediates T cell anergy, exhaustion, and tumor regression. In this study, we investigated increased cytotoxic activity after the blockade of PD-1 for effective immunotherapy.
Methods
The cytotoxic function of expanded CD8+ CTLs and interactions with tumor cells investigated after blocking of PD-1. Ex vivo expanded CD8+ CTLs were co-cultured with mismatch repair (MMR) stable or deficient (high microsatellite instability [MSI-H]) EpCAM+ tumor cells. The levels of IFN-γ and GrB were detected by enzyme-linked immunosorbent spot assay. Flow cytometry and confocal microscopy were used to assess CD107a mobilization, cytosolic uptake, and cell migration.
Results
A dramatic increase in PD-1 expression on the surface of CD8+ CTLs during ex vivo expansion was observed. PD-1 level was downregulated by approximately 40% after incubation of the CD8+ CTLs with monoclonal antibody which enhanced the secretion of IFN-γ, GrB, and CD107a. Additionally, PD-1 blockade enhanced cell migration and cytosolic exchange between CD8+ CTLs and MMR deficient (MSI-H) EpCAM+PD-L1+ tumor cells.
Conclusion
The blockade of PD-1 enhanced the cytotoxic efficacy of CD8+ CTLs toward MMR deficient tumor cells. In conclusion, we propose that blocking of PD-1 during the expansion of CD8+ CTLs may improve the clinical efficacy of cell-based adoptive immunotherapy.
Introduction
Following the success and approval of anti-CTLA-4 with ipilimumab and anti-PD-1 immunotherapy with nivolumab to treat both metastatic melanomaCitation1,Citation2 and pre-treated metastatic squamous non-small-cell lung cancer (NSCLC),Citation3,Citation4 there has been interest in shifting the focus to controlling immunosuppressive mechanisms.Citation5–Citation7 One of the most important mechanisms underlying the significant clinical benefits of immune checkpoint blockade is associated with activating anti-tumor T cell immunity and attenuating T cell exhaustion.Citation8,Citation9 The expression of inhibitory receptors (iRs) by CD8+ T cells is generally considered a hallmark of impaired T cell function, especially during chronic viral infection and cancer.Citation10,Citation11 Over the last decade, the family of iRs has been identified in T cell exhaustion, including PD-1, CTLA-4, mucin domain protein-3, LAG3, BTLA, and TIGIT.Citation8,Citation12
Although the mechanism of underlying T cell exhaustion is complex, an increasing amount of evidence indicates that blocking PD-1 is thus far the most effective means of restoring CD8+ T cell function. Recently, the results of a study of 46 patients with advanced melanoma who received a single agent, pembrolizumab, a PD-1 monoclonal antibody (mAb), showed that the presence of CD8+ T cells at the invasive tumor margin was associated with the expression of the PD-1/PD-L1 immune inhibitory axis, and these cells are predictive biomarkers for PD-1 mAb treatment.Citation13 Moreover, treatment with PD-1 mAb successfully attenuates T cell exhaustion, resulting in the proliferation of intra-tumoral CD8+ T cells and restoring potent CTL-mediated responses in patients who experienced tumor regression. The presence and localization of CD8+ T cells within the tumor microenvironment has been associated with prognostic significance in a number of solid tumors including melanoma, NSCLC,Citation14 colorectal cancer,Citation15,Citation16 renal cell carcinoma, triple-negative breast cancer,Citation17 ovarian cancer,Citation18 glioma,Citation19 and urothelial carcinoma,Citation20 in addition, the successful treatment of anti-PD-1 induces robust clinical responses in these cancer types. Furthermore, a genetic biomarker for the effectiveness of anti-PD-1 treatment called mismatch repair (MMR)-deficiency was recently identified.Citation21 These findings provide additional evidence indicating that PD-1 blockade counteracts T cell exhaustion, resulting in endogenous anti-tumor immune responses in the form of CTL activity toward mutation-associated neoantigen recognition.
In contrast to using anti-PD-1 antibodies which target endogenous PD-1 on T cells within the human body, adoptive immunotherapy involves the infusion of autologous ex vivo activated and expanded T cells into patients with the aim of combating the tumor.Citation22 Although the T cells used in adoptive immunotherapy can be derived from different sources, such as tumor infiltrating lymphocytes (TILs), peripheral blood mononuclear cells (PBMCs), and tumor draining lymph nodes, the general procedure involves T cell isolation, ex vivo activation/genetic manipulation, expansion, and then transfusion.Citation23 Previous studies have shown that during the chronic ex vivo activation and expansion of tumor-specific T cellsCitation24,Citation25 or MHC-independent chimeric antigen receptor (CAR) T cells,Citation25 the expression of iRs is significantly upregulated on the expanded T cells, leading to exhaustion and poor in vivo persistence. The autologous transfusion of T cells with high levels of iRs into patients might explain the poor clinical efficacy of adoptive immunotherapy against solid tumors. In the present study, we aimed to determine the effect of PD-1 blockade on ex vivo expanded CD8+ T cells which were subjected to the standard activation and expansion procedure for adoptive immune cell therapy.
Materials and methods
Patient samples and cell lines
The ethics committee of The Affiliated Hospital of Guizhou Medical University, People’s Republic of China approved this research. Human colon cancer cell lines were procured from American Type Cell Collection (ATCC, Manassas, VA, USA). Primary tumor samples–peripheral blood specimens, were obtained from the colorectal surgical department or from healthy volunteers after written informed consent which was obtained from the patients and donors. PBMCs were separated by density gradient centrifugation using Polymorphprep™ (AXIS-SHIELD PoC AS, Oslo, Norway) cell separation media. Microsatellite instability (MSI) was used to investigate the standard Bethesda microsatellite markers (Bat-25, Bat-26, D5S346, D17S250, and D2S123) by using fragment analysis of polymerase chain reaction products which were labeled with fluorescent dyes. Differences in the length of two or more markers were indicative of MSI status. Five high MSI (MSI-H) tumor specimens were selected for tumor epithelial cells’ (TECs’) isolation ().
Table 1 Disease characteristics of five colorectal patients selected for TEC isolation
Primary TECs and cell lines
The following TECs and colon cancer cell lines were selected for noted expression of cytokeratin 20 (CK20): MMR deficient (MSI-H) TECs, DLD-1, and MMR proficient (MSS) SW480, SW620.Citation26 TECs were isolated from post-surgically excised colon cancer tissues as previously described.Citation27 Briefly, tumor tissues were collected in RPMI 1640 medium supplemented with 100 U/mL penicillin/streptomycin, 10% heat-inactivated FBS, and 50 μg/mL gentamycin. The tumor tissues were further minced into small pieces in a dissociation solution containing collagenase, DNAse, and trypsin (Thermo Fisher Scientific, Waltham, MA, USA) for 48 hrs at 37°C. The suspensions were washed using PBS and centrifuged at 1,200 rpm for 3 min. The cell pellets were resuspended in RPMI 1640 medium supplemented with 10% FBS, gentamycin, hydrocortisone, and insulin and then seeded into a T25 flask. EpCAM+ TECs were purified using microbeads (Miltenyi Biotec Inc., Bergisch Gladbach, Germany) as described by the manufacturer, and the viability of purified TECs was determined by trypan exclusion assay. The isolated EpCAM+ TECs and colon cancer cell lines were utilized for further experiments. For the DLD-1 cells, RPMI 1640 medium was used for culture and passage, while Leibovitz media (Thermo Fisher Scientific) was used for culture and passage of SW480 and SW620. Both media were supplemented with 10% FBS (Thermo Fisher Scientific), and the cells were maintained in a humidified, 5% CO2 atmosphere at 37°C.
Generation of CTLs
PBMCs (1.5×106) were cultured overnight in T175 cell culture flasks in AIM V medium (Thermo Fisher Scientific) supplemented with 1% FBS and 500 U/mL IFN-γ. After 18 to 24 hrs, 50 ng/mL anti-CD3/28 antibody (OKT3) coated microbeads and 500 U/mL rIL-2 were added to the media. The cells were incubated at 37°C in a humidified, 5% CO2 atmosphere. Following the standard procedure of large-scale manufacturing, on day 7, the cell culture from each T175 flask was transferred to a disposable and gas-permeable cell expansion bag (CultiLife, Takara Bio, Otsu, Japan) in a final volume of 4–5 L. Flow cytometry was performed to evaluate the PD-1 levels on the T cells before and after 2 weeks of ex vivo expansion.
Apoptosis assay
The CD8+ T cells were harvested on day 14 and were sequentially co-cultured for another 48 hrs with cancer cells at an effector (CD8+ T cells)-to-target (EpCAM+ cancer cells) ratio of 25:1 in the presence of 1 μg/mL of anti-PD-1 purified antibody (eBioscience, San Diego, CA, USA) along with an isotype control.Citation28 The co-culture was incubated with neutralizing PD-1 (nPD-1 mAb; BPS Bioscience, San Diego, CA, USA) antibody for 20 minutes before adding blocking PD-1 for neutralization assay. Cells were harvested and washed in 2× PBS and centrifuged at 1,500 rpm for 10 min. The cells were pre-stained with EpCAM PE (BD Biosciences, San Jose, CA, USA) for 30 min and 5 μL of Annexin V per 0.5×106 was used to reconstitute cells in ice-cold binding buffer for 15 min at 4°C. The cells were acquired using flow cytometry (FC500, Beckman Coulter, Brea, CA, USA) and analyzed by CXP software.
Phenotyping and the CD107a-mobilization assay by flow cytometry
The antibodies used for CTL phenotyping (anti-CD3-ECD, CD4-PC5, CD8-PC7, PD-1-PE, CTLA-4PE, PD-L1 FITC; IgG1-FITC, IgG1-PE) were all obtained from BD Biosciences. To investigate competitive binding of staining antibody with blocking PD-1 mAb, flow cytometric analysis was performed using two different clones of staining antibody after 48 hrs: primary PD-1 (MIH4), secondary IgG1 FITC and PD-1 conjugated with PE.
Positive staining for PD-1 and PD-L1 was determined by comparing with the respective isotype control. The cell suspensions were incubated with mAb for 30 min at 4°C and then washed twice in PBS. PD-1 mAb-treated co-cultures were incubated with an anti-CD107a antibody and degranulation assay was performed.Citation29 The cells were acquired on an FC 500 (Beckman Coulter) and analyzed by the CXP analysis software.
Enzyme-linked immunosorbent spot (ELISpot) assay
To measure IFN-γ and GrB secretion during the co-culturing of CD8+ T and cancer cells, an ELISpot experiment was conducted using MultiScreen 96-well plates (Mabtech, Nacka Strand, Sweden) in triplicate. Wells that contained only cancer cells served as negative controls, and those that contained anti-CD28 (1:1,000) served as positive controls. The spots were visualized and counted on an Immunospot Imaging Analyzer (Cellular Technology Ltd, Shaker Heights, OH, USA).
Chemotaxis assay
The migration of CD8+ T cells was evaluated in vitro using a transwell insert (Corning Incorporated, Corning, NY, USA) with an 8.0 μm-pore size, as described previously.Citation30 Briefly, T cells were pre-treated with anti-PD-1 antibody for 48 hrs and then washed once with PBS. The cell count was then adjusted to 5×105 cells/mL using RPMI 1640 medium, and a 100 μL aliquot of the suspension was placed in the top chamber of the transwell insert. The cancer cells were cultured in RPMI 1640 medium, and conditioned medium was added to the bottom chamber of the transwell plate prior to the addition of the CTLs to the top chamber. After the cells were incubated for 90 min at 37°C in a 5% CO2 atmosphere, the top chamber was removed. The cells were stained with Giemsa, and the number of T cells which had migrated into the bottom chamber was counted using the trypan blue exclusion assay.
Statistical analysis
All quantitative data are presented as the means with standard errors of at least three or more independent experiments. Student’s t-tests were performed to analyze statistical significance. P-values <0.05 were considered significant.
Results
Extracellular level of EpCAM and PD-L1 in the colon cancer cell cultures
Primary tumors (5 μm paraffinized sections) were chromogenically immunostained for PD-L1, and found positive at the invasive front (Figure S1A). MHC-I/II expression levels were assessed by flow cytometry. We found a variable range of MHC-I (mean 94.6%, range 96.6%–90.1%) and MHC-II (mean 4%, range 2%–5%) expression level (Figure S1B). The purified TECs and colon cancer cell lines, including DLD-1, SW480, and SW620, were tested to determine cell viability, EpCAM, and intracellular CK20 levels. The viability of purified TECs was found to be 92.9% (range 89.62%–96.47%; n=5) before further experiments were pursued. The TECs and colon cancer cell lines were observed to be ~100% positive for EpCAM, and >90% for CK20 (Figure S2A and B). We further analyzed PD-L1 expression in these cells. We found a significant variation in PD-L1 expression levels on the TECs and colon cancer cell lines: TECs (range 92%–97%, mean =94.33%); DLD-1 (range 13.3%–21.0%, mean =17.23%); SW480 (<1%); SW620 (<1%); and K562 (<0.5%) (Figure S2C).
PD-1 levels on CD8+ T cells before and after ex vivo expansion
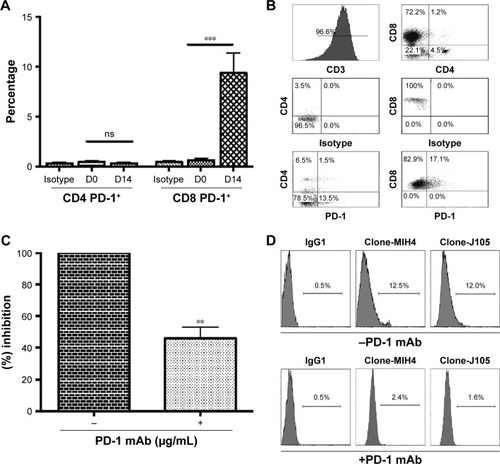
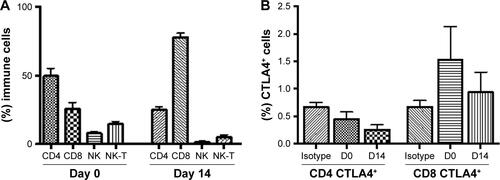
The CD8+ T cells were expanded using classic methods for 2 weeks using patients’ PBMCs via the timed addition of IFN-γ, anti-CD3/CD28 coated microbeads, and rIL-2. The median expansion of the T cells was ~1,000-fold, and the majority of the cells were CD3+CD8+ T cells (range 64%–87%, mean =79%) after expansion.Citation31,Citation32 The presence of other cytotoxic cell types, including CD3−CD56+ NK cells (range 0.1%–5.4%; mean =1.72%) and CD3+CD56+ NKT cells (range 0.3%–15.1%; mean =6.34%) was also determined (Figure S3A) and found to be unchanged. CTLA-4 and PD-1 expression was determined and we observed a significant increase in the surface expression of PD-1 in most of the CD8+ T cells (range 1.6%–31%, mean =9.4% P<0.0001) (), whereas the changes in CTLA-4 expression were unremarkable (Figure S3B). PD-1 expression was significantly lower (P=0.0037) () after incubating with a PD-1 mAb for 48 hrs. Flow cytometric analysis revealed that anti-PD-1 (J105) binds a different epitope than the MIH-4 epitope of purified PD-1 mAb ().
Figure 1 The PD-1 expression on ex vivo expanded CD8+ T cells obtained from patient’s blood before surgery.
Abbreviations: mAb, monoclonal antibody; ns, not significant.
PD-1 blockade enhances the cytotoxic effect of CD8+ T cells against MMR deficient EpCAM+PD-L1+ cancer cells
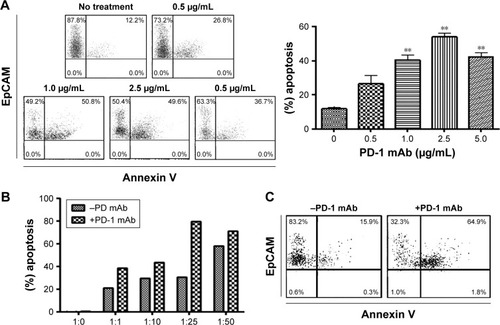
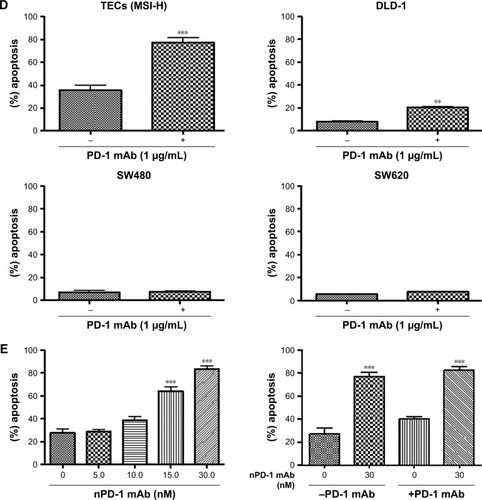
To test the effect of the PD-1 blockade on the anti-tumor activity of ex vivo expanded CD8+ T cells, we evaluated cytotoxicity in colorectal cancer cells in the presence of a PD-1 mAb. We observed an insignificant apoptotic effect in MMR proficient TECs in the presence of PD-1 mAb after 48 hrs (Figure S1C). Therefore, MMR proficient TECs were excluded for further experiments. A dose dependent treatment was carried out at 0.5, 1.0, 2.5, and 5.0 μg/mL to select the optimal dose of PD-1 mAb for the subsequent experiments. The best killing activity of CD8+ T cells was achieved at doses from 1.0–2.5 μg/mL (). Therefore, 1.0 μg/mL was chosen as the concentration for the following experiments. We further selected an effector-to-target (ET) ratio of 25:1 based on the co-incubation of TECs with CD8+ T cells at different ET ratios (). Flow cytometric analysis detected a significant increase in apoptosis achieved with or without PD-1 mAb () in EpCAM+PD-L1+ cells: MMR deficient TECs (P=0.005) and DLD-1 cells (P=0.0025) (). Indeed, no noteworthy apoptotic effect in MMR proficient SW480 (P=0.350) and SW620 (P=0.050) were observed. We observed dose-dependent (5, 10, 15, and 30 nM) apoptosis to select suitable concentration of nPD-1 mAb, and found no significant effect of blocking PD-1 mAb in the presence of nPD-1 mAb () in TECs.
Figure 2 Apoptotic effect of CD8+ T cells on EpCAM+ tumor cells after PD-1 blockade for 48 hrs.
Abbreviations: mAb, monoclonal antibody; TECs, tumor epithelial cells; MSI-H, high microsatellite instability.
PD-1 blockade increases cytotoxic cytokine secretion and degranulation of CD8+ T cells against MMR deficient EpCAM+PD-L1+ cancer cells
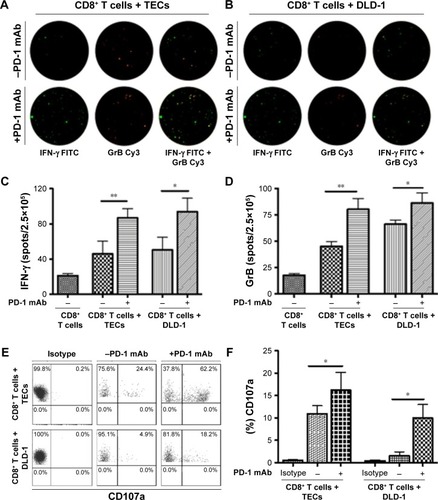
We performed quantitative ELISpot assays for both IFN-γ and GrB after cells were incubated with the PD-1 mAb at 1.0 μg/mL (). IFN-γ secretion was higher in CD8+ T cells which were co-cultured with TECs (P=0.0256) or DLD-1 cells (P=0.0361) than in the control (). Similar results were also observed in GrB+ CD8+ T cells co-cultured with TECs (P=0.0072) or DLD-1 cells (P=0.0461) (). To determine the level of degranulation, CD107a mobilization was studied using flow cytometry (). PD-1 blockade enhanced the surface CD107a mobilization on CD8+ T cells which were co-cultured with TECs (P=0.018) or DLD-1 cells (P=0.0782), as shown in .
Figure 3 Effect of PD-1 mAb on cytolytic IFN-γ, GrB, and CD107a.
Abbreviations: mAb, monoclonal antibody; TECs, tumor epithelial cells.
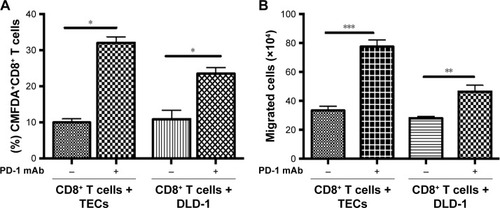
PD-1 blockade enhanced the interaction and cytosolic uptake between CD8+ T cells and EpCAM+PD-L1+ cancer cells
To analyze the chemotactic interactions between CD8+ T cells and tumor cells after PD-1 blockade, tumor cells were stained with the non-invasive dye CMFDA (5 μM), for 20 min at 37°C and then co-cultured with CD8+ T cells for 48 hrs.Citation33 After culture, the cells were analyzed by flow cytometry. PD-1 blockade significantly increased the proportion of CD8+CMFDA+ cells among T cells co-cultured with TECs (P=0.0002) or DLD-1 cells (P=0.05) (). We evaluated chemotactic attraction using a transwell migration assay. Pretreatment with PD-1 mAb at 1.0 μg/mL significantly enhanced the number of CD8+ T cells which migrated to the lower chamber of the transwell insert in the presence of TECs (P<0.0001) or DLD-1 cells (P=0.0015), as shown in .
Figure 4 Cellular interactions, cytosolic uptake, and CD8+ T cells’ migration after 48 hrs.
Abbreviations: mAb, monoclonal antibody; TECs, tumor epithelial cells.
Discussion
In recent years, cancer immunotherapy has emerged as an efficacious treatment modality which produces clinically valid outcomes in certain types of cancer.Citation34,Citation35 The most promising strategies for cancer immunotherapy include a variety of approaches, such as the use of therapeutic antibodies which target immune checkpoint molecules, dendritic cell-based vaccines, adoptive transfer of ex vivo activated T cells, and the combinations of these strategies with or without chemo/radiotherapy.Citation36–Citation38
Although the impressive treatment outcomes have been obtained using TILs and CD19 targeted CAR T cells, demonstrating the promise of T cell-based adoptive immunotherapy, these successes have not been translated beyond early stage clinical studies.Citation35 It has become apparent that tumors develop multiple inhibitory networks to effectively evade immune surveillance.Citation39 The immunosuppressive microenvironment has been recognized as one of the hallmarks of tumors.Citation40 Strategies aimed at attenuating the endogenous immunosuppressive environment using non-myeloablative chemotherapy, irradiation or therapeutic mAbs against immune checkpoint inhibitors have been shown to improve the clinical efficacy of adoptive T cell immunotherapy for solid tumors.Citation41 However, lower effectiveness of ex vivo expanded T cells which express high levels of iRs is often ignored.
Adoptive T cell therapy generally requires the ex vivo expansion of a large number of T cells to allow an infusion to achieve therapeutic benefits. The CD3/TCR complex stimulation of tumor reactive T cells mediated by co-stimulatory signals to retain proliferation and functional properties of antigen specific T cells.Citation32 This procedure is similar to that used to expand tumor-specific T cells, non-specific cytotoxic cells (eg, NK, NKT, CD8+ T), and MHC-independent CAR T cells. Chronic exposure to cytokines or tumor antigens leads to the expansion of T cells and causes them to develop artificial exhaustion prior to transfusion.Citation42,Citation43 In our study, we demonstrate that surface expression of PD-1 on the expanded CD8+ T cells was dramatically increased (by 10-fold) during the classic ex vivo expansion procedure with OKT3 and IL-2. This finding supports a previous study which showed that transfused TILs produced a high PD-1 level in a melanoma animal model.Citation44 We showed that blocking PD-1 interaction after ex vivo expansion significantly enhanced the cytotoxicity of CD8+ T cells toward EpCAM+PD-L1+ cancer cells, and this increased the release of the cytotoxic IFN-γ and GrB, as well as CD8+ T cell degranulation. Moreover, PD-1 blockade improved the interaction and cytosolic exchange between CD8+ T cells and EpCAM+PD-L1+ cancer cells.
The expression of PD-L1 on cancer cells has been associated with poor prognosis in cancer, demonstrating that PD-1/PD-L1 pathway-mediated immune tolerance has clinical significance.Citation39,Citation40 The results of a previous study demonstrated that PD-1 blockade partially enhanced the cytotoxicity of PD-1+ CD8+ T cells against lymphoma.Citation41
The CK20 identified as a diagnostic and prognostic biomarker that classified in colorectal cancer by MMR status, and the inverse correlations have observed in the higher tumor grade. However, the association between MMR status and tumor grades has not been specifically investigated.Citation45 In our study we found typical expression of CK20 in MMR deficient TECs, which were isolated from the primary tumor site.
The clinical trial of PD-1 mAb reached the primary endpoint, showing striking immune related response and significant disease control rate. Perhaps the increased mutation-associated tumor antigens need to be further demonstrated in MMR deficient colon cancer.Citation21,Citation46 In the current study, we have shown that PD-1 blockade induced a high killing effect in MMR deficient (MSI-H) EpCAM+PD-L1+ cancer cells. Mechanistically, we have shown that EpCAM+PD-L1+ TECs and DLD-1 cells engage PD-1+ T cells via the PD-1 ligand. Our results demonstrate that PD-1 blockade enhanced cancer cell apoptosis, the production of cytolytic molecules, and CD8+ T cell degranulation. Moreover, it was previously shown that blocking PD-1 promoted the migration of transfused T cells to tumor sites.Citation38 Our data also indicate that PD-1 blockade promotes the migration and cytosolic exchange between CD8+ T cells with EpCAM+PD-L1+ TECs and DLD-1 cancer cells.
Our results offer a viable means of restoring T cell exhaustion during the traditional expansion procedure and obtaining highly functional CD8+ T cells which can be used for adoptive immunotherapy. Furthermore, this function can be restored after blocking PD-1 using mAb, peptides, small molecule inhibitors and advance knock-out technologies during ex vivo expansion prior to T cell transfusion.Citation47–Citation49 The transfusion of effective CD8+ CTLs with lower iRs might be associated with better clinical efficacy, but verification of this hypothesis awaits further human studies.
Conclusion
One of the challenges of adoptive T cell transfusion is the lack of efficient tumor-killing T cells, which results in poor clinical responses. Here, we investigated whether blocking of PD-1 on CD8+ CTLs during ex vivo expansion could potentiate the tumor-killing activity and therefore, achieve effective adoptive immunotherapy. We demonstrated that after ex vivo expansion, the level of PD-1 on CD8+ CTLs increased dramatically. PD-1 blockade could significantly potentiate the cytotoxic activity of CD8+ CTLs against MMR deficient EpCAM+PD-L1+ tumor cells in addition to increasing the secretion of IFN-γ, GrB, and CD107a. Moreover, PD-1 blockade enhanced cell migration and cytosolic exchange between CD8+ CTLs and EpCAM+PD-L1+ tumor cells. Our findings might provide a new insight that may improve the clinical efficacy of cell-based adoptive immunotherapy.
Author contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (project no 81560499 & 81460448), the Guizhou Science & Technology Fund (project no 20127016), and the Chinese Ministry of Science and Technology (project no 2013ZX09101015). The abstract of this paper was presented at the International Congress of Immunology 2016 (ICI-2016), August 21–26, 2016 in Melbourne, Australia as a poster presentation with interim findings. The poster’s abstract was published in European Journal of Immunology.
Supplementary materials
Figure S1 Information on primary tumor.
Notes: (A) PD-L1 staining on 5 μM tumor sections utilized for TECs’ isolation. (B) MHC-I/II staining of isolated TECs shown by flow cytometry, and (C) apoptotic effect of CD8+ T cells on TECs (MSS).
Abbreviations: TECs, tumor epithelial cells; Pt, patient; MSS, MMR proficient; MMR, mismatch repair.
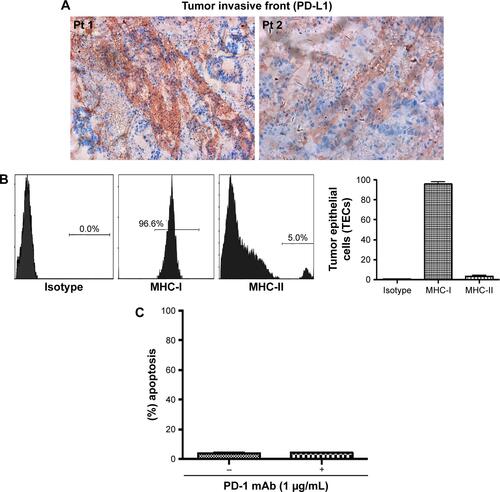
Figure S2 The representative dot plot of flow cytometry staining of tumor antigens, and average level on TECs, DLD-1, SW480, and SW620 along with K562 as a negative control (n=3).
Note: (A) Extracellular EpCAM, (B) intracellular CK20, and (C) intracellular PD-L1.
Abbreviations: TECs, tumor epithelial cells; CK20, cytokeratin 20; SS lin, side scatter linear; FS lin, forward scatter linear.
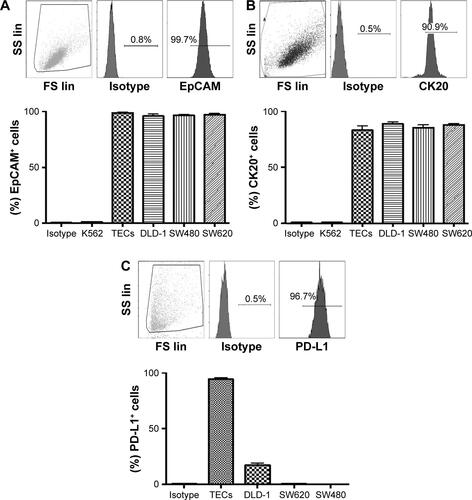
Figure S3 Percentages of (A) killer immune cells, and (B) intracellular CTLA4 before and after ex vivo T cell expansion by flow cytometry (n=11).
Disclosure
The authors report no conflicts of interest in this work.
References
- ErricoAMelanoma: CheckMate 067 – frontline nivolumab improves PFS alone or in combination with ipilimumabNat Rev Clin Oncol201512843526099982
- ValsecchiMECombined nivolumab and ipilimumab or monotherapy in untreated melanomaN Engl J Med2015373131270
- YaqubFNivolumab for squamous-cell non-small-cell lung cancerLancet Oncol2015167e319
- BorghaeiHPaz-AresLHornLNivolumab versus docetaxel in advanced nonsquamous non-small-cell lung cancerN Engl J Med2015373171627163926412456
- ButtAQMillsKHImmunosuppressive networks and checkpoints controlling antitumor immunity and their blockade in the development of cancer immunotherapeutics and vaccinesOncogene201433384623463124141774
- ByrneWLMillsKHLedererJAO’SullivanGCTargeting regulatory T cells in cancerCancer Res201171226915692022068034
- JarnickiAGLysaghtJTodrykSMillsKHSuppression of antitumor immunity by IL-10 and TGF-beta-producing T cells infiltrating the growing tumor: influence of tumor environment on the induction of CD4+ and CD8+ regulatory T cellsJ Immunol2006177289690416818744
- WherryEJKurachiMMolecular and cellular insights into T cell exhaustionNat Rev Immunol201515848649926205583
- Twyman-Saint VictorCRechAJMaityARadiation and dual checkpoint blockade activate non-redundant immune mechanisms in cancerNature2015520754737337725754329
- WherryEJT cell exhaustionNat Immunol201112649249921739672
- LegatASpeiserDEPircherHZehnDFuertes MarracoSAInhibitory receptor expression depends more dominantly on differentiation and activation than “exhaustion” of human CD8 T cellsFront Immunol2013445524391639
- JiangYLiYZhuBT-cell exhaustion in the tumor microenvironmentCell Death Dis20156e179226086965
- TumehPCHarviewCLYearleyJHPD-1 blockade induces responses by inhibiting adaptive immune resistanceNature2014515752856857125428505
- DonnemTHaldSMPaulsenEEStromal CD8+ T-cell density – a promising supplement to TNM staging in non-small cell lung cancerClin Cancer Res201521112635264325680376
- LingAEdinSWikbergMLObergAPalmqvistRThe intratumoural subsite and relation of CD8(+) and FOXP3(+) T lymphocytes in colorectal cancer provide important prognostic cluesBr J Cancer2014110102551255924675384
- MeiZLiuYLiuCTumour-infiltrating inflammation and prognosis in colorectal cancer: systematic review and meta-analysisBr J Cancer201411061595160524504370
- MellaMKauppilaJHKarihtalaPTumor infiltrating CD8+ T lymphocyte count is independent of tumor TLR9 status in treatment naive triple negative breast cancer and renal cell carcinomaOncoimmunology201546e100272626155410
- StumpfMHasenburgARienerMOIntraepithelial CD8-positive T lymphocytes predict survival for patients with serous stage III ovarian carcinomas: relevance of clonal selection of T lymphocytesBr J Cancer200910191513152119861998
- HanSZhangCLiQTumour-infiltrating CD4(+) and CD8(+) lymphocytes as predictors of clinical outcome in gliomaBr J Cancer2014110102560256824691423
- SharmaPShenYWenSCD8 tumor-infiltrating lymphocytes are predictive of survival in muscle-invasive urothelial carcinomaProc Natl Acad Sci U S A2007104103967397217360461
- LeDTUramJNWangHPD-1 blockade in tumors with mismatch-repair deficiencyN Engl J Med2015372262509252026028255
- RestifoNPDudleyMERosenbergSAAdoptive immunotherapy for cancer: harnessing the T cell responseNat Rev Immunol201212426928122437939
- WangXRiviereIManufacture of tumor- and virus-specific T lymphocytes for adoptive cell therapiesCancer Gene Ther2015222859425721207
- JanelleVCarliCTailleferJOrioJDelisleJSDefining novel parameters for the optimal priming and expansion of minor histocompatibility antigen-specific T cells in cultureJ Transl Med20151312325925868
- FourcadeJSunZBenallaouaMUpregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patientsJ Exp Med2010207102175218620819923
- YuanZSotsky KentTWeberTKDifferential expression of DOC-1 in microsatellite-unstable human colorectal cancerOncogene200322406304631013679870
- OikonomouEKothonidisKTaoufikENewly established tumourigenic primary human colon cancer cell lines are sensitive to TRAIL-induced apoptosis in vitro and in vivoBr J Cancer2007971738417551494
- Dons’koiBVChernyshovVPOsypchukDVMeasurement of NK activity in whole blood by the CD69 up-regulation after co-incubation with K562, comparison with NK cytotoxicity assays and CD107a degranulation assayJ Immunol Methods20113721–218719521839083
- Hardtke-WolenskiMKrausLSchmetzCExchange of cytosolic content between T cells and tumor cells activates CD4 T cells and impedes cancer growthPloS One2013810e7855824205259
- DemmersMWBaanCCvan BeelenEIjzermansJNWeimarWRowshaniATDifferential effects of activated human renal epithelial cells on T-cell migrationPloS One201385e6491623717673
- TrickettAKwanYLT cell stimulation and expansion using anti-CD3/CD28 beadsJ Immunol Methods20032751–225125512667688
- TeschnerDWenzelGDistlerEIn vitro stimulation and expansion of human tumour-reactive CD8+ cytotoxic T lymphocytes by anti-CD3/CD28/CD137 magnetic beadsScand J Immunol201174215516421517928
- LiuMFWengCTWengMYVariable increased expression of program death-1 and program death-1 ligands on peripheral mononuclear cells is not impaired in patients with systemic lupus erythematosusJ Biomed Biotechnol2009200940613619759858
- Couzin-FrankelJBreakthrough of the year 2013. Cancer immunotherapyScience201334261651432143324357284
- RosenbergSARestifoNPAdoptive cell transfer as personalized immunotherapy for human cancerScience20153486230626825838374
- MeleroIBermanDMAznarMAKormanAJGraciaJLHaanenJEvolving synergistic combinations of targeted immunotherapies to combat cancerNat Rev Cancer201515845747226205340
- MaudeSLFreyNShawPAChimeric antigen receptor T cells for sustained remissions in leukemiaN Engl J Med2014371161507151725317870
- PodrazilMHorvathRBechtEPhase I/II clinical trial of dendritic-cell based immunotherapy (DCVAC/PCa) combined with chemotherapy in patients with metastatic, castration-resistant prostate cancerOncotarget2015620181921820526078335
- FinnOJImmuno-oncology: understanding the function and dysfunction of the immune system in cancerAnn Oncol201223Suppl 8viii6viii922918931
- HanahanDWeinbergRAHallmarks of cancer: the next generationCell2011144564667421376230
- MahnkeKSkorokhodAGrabbeSEnkAHOpening a niche for therapy: local lymphodepletion helps the immune system to fight melanomaJ Invest Dermatol201413471794179624924759
- FourcadeJKudelaPSunZPD-1 is a regulator of NY-ESO-1-specific CD8+ T cell expansion in melanoma patientsJ Immunol200918295240524919380770
- LongAHHasoWMShernJF4-1BB costimulation ameliorates T cell exhaustion induced by tonic signaling of chimeric antigen receptorsNat Med201521658159025939063
- PengWLiuCXuCPD-1 blockade enhances T-cell migration to tumors by elevating IFN-gamma inducible chemokinesCancer Res201272205209521822915761
- LugliATzankovAZlobecITerraccianoLMDifferential diagnostic and functional role of the multi-marker phenotype CDX2/CK20/CK7 in colorectal cancer stratified by mismatch repair statusMod Pathol200821111403141218587323
- DudleyJCLinMTLeDTEshlemanJRMicrosatellite instability as a biomarker for PD-1 blockadeClin Cancer Res201622481382026880610
- ChangHNLiuBYQiYKBlocking of the PD-1/PD-L1 interaction by a D-peptide antagonist for cancer immunotherapyAngew Chem Int Ed Engl20155440117601176426259671
- ZhanMMHuXQLiuXXRuanBFXuJLiaoCFrom monoclonal antibodies to small molecules: the development of inhibitors targeting the PD-1/PD-L1 pathwayDrug Discov Today20162161027103627094104
- SuSHuBShaoJCRISPR-Cas9 mediated efficient PD-1 disruption on human primary T cells from cancer patientsSci Rep201662007026818188