Abstract
Molecularly targeted therapies are rapidly growing with respect to their clinical development and impact on cancer treatment due to their highly selective anti-tumor action. However, many aggressive cancers such as triple-negative breast cancer (TNBC) currently lack well-defined therapeutic targets against which such agents can be developed. The identification of tumor-associated antigens and the generation of antibody drug-conjugates represent an emerging area of intense interest and growth in the field of cancer therapeutics. Glycoprotein non-metastatic b (GPNMB) has recently been identified as a gene that is over-expressed in numerous cancers, including TNBC, and often correlates with the metastatic phenotype. In breast cancer, GPNMB expression in the tumor epithelium is associated with a reduction in disease-free and overall survival. Based on these findings, glembatumumab vedotin (CDX-011), an antibody-drug conjugate that selectively targets GPNMB, is currently being investigated in clinical trials for patients with metastatic breast cancer and unresectable melanoma. This review discusses the physiological and potential pathological roles of GPNMB in normal and cancer tissues, respectively, and details the clinical advances and challenges in targeting GPNMB-expressing malignancies.
Introduction
Breast cancer is a highly prevalent and devastating disease. Despite clear advances in screening, more accurate prognosis, and disease management over the past few decades, each year more than 1,600,000 cases are diagnosed and 420,000 deaths are attributed to breast cancer worldwide.Citation1 Breast cancer remains the most commonly diagnosed cancer, and one of the most significant causes of cancer-related deaths in women.Citation2
One of the primary challenges associated with the treatment of breast cancer is tumor heterogeneity, which is manifested by the diversity of histopathologies and molecular features associated with this disease. In the early 2000s, genomic studies employed gene expression signatures to classify breast cancer into five distinct subgroups, which include the luminal A, luminal B, HER2+, triple negative/basal-like, and normal breast-like subtypes.Citation3–Citation5 More recently, the evolution of gene expression profiling techniques has allowed further subclassification of breast cancer through the identification of the claudin-low subtype.Citation6,Citation7 The molecular complexity and heterogeneity of these subtypes is continually being refined as additional genomic, epigenomic, and transcriptomic data becomes available.Citation8–Citation12
Gene expression profiling has also proven useful in identifying patients with a high risk of disease progression and distant recurrence.Citation13 In general, patients with basal-like and HER2+ signatures displayed the shortest relapse-free survival rates, while a luminal A classification was associated with the lowest risk of developing distant metastases.Citation4 Breast cancer most commonly metastasizes to bone, followed by lung, liver, and brain, and the site of distant metastasis can predict the likelihood of overall survival. Typically, the presence of visceral metastasis or metastasis in multiple sites is associated with a shorter survival.Citation14 Accordingly, the majority of luminal A breast cancers metastasize exclusively to bone, while HER2 enriched cancers preferentially give rise to liver and lung metastases, and basal-like breast cancers are associated with increased liver and brain metastasis.Citation15,Citation16
The current availability of therapeutic targets varies according to the molecular subtype, which accounts in part for the differences between these groups with respect to survivalCitation3–Citation5,Citation17,Citation18 and response to therapy.Citation7,Citation19–Citation22 Luminal breast cancers are associated with the best prognosis and are characterized by the presence of estrogen and progesterone receptors, which makes them amenable to hormonal therapies such as tamoxifen or aromatase inhibitors. Similarly, drugs that target the HER2 receptor, such as trastuzumab, pertuzumab, and lapatinib, are clinically approved for the treatment of HER2+ breast cancers. Triple-negative breast cancers (TNBC) are diagnosed by the lack of ER, PR and HER2 expression and are largely characterized by a basal-like histopathology (basal-like breast cancer [BLBC]). TNBCs account for an estimated 10%–25% of invasive breast cancers, are associated with a high grade and poor prognosis, and, due to a lack of distinct molecular markers, there are currently no targeted interventions for this aggressive subset of the disease. Although poly ADP ribose polymerase (PARP) inhibitors have shown encouraging results for the BRCA-subset of TNBC patients,Citation23,Citation24 their utility in this subtype is not assured.Citation25 At present, chemotherapy remains the primary treatment option for TNBC.
One of the main problems associated with the use of chemotherapy for the treatment of cancer is the off-target action of the drugs on normal cells, which can lead to painful side effects and complications. In an effort to minimize the cytotoxicity of these therapies, approaches that selectively target tumor-associated antigens are emerging as promising therapeutic strategies for TNBC and other cancers. One such approach is the development of antibody-drug conjugates (ADCs), which synergistically combine the specificity of antibodies with the cytotoxic efficacy of chemotherapy. ADCs consist of antibodies bound to highly potent cytotoxins by a chemical linker.Citation26 These antibodies can be designed to target tumor-specific proteins and thereby serve as vehicles that deliver the drug to the cell of interest, often via internalization of the compound.Citation26 Accordingly, the expression pattern of the selected antigen, both in normal and cancer tissues, is an important consideration in predicting response to ADC therapy.
GPNMB has been recently identified as a potential therapeutic target for patients with BLBC and TNBC.Citation27–Citation29 GPNMB expression in the breast tumor epithelium was shown to strongly correlate with disease-free and overall survival. Additionally, GPNMB is highly expressed in BLBC and TNBC and its levels are associated with a poor prognosis and increased risk for recurrence in this subset. These findings, combined with evidence of high GPNMB expression in numerous cancers,Citation27–Citation34 have sparked an interest in investigating GPNMB as a target for antibody-based therapies in TNBC and other cancers.Citation35–Citation37 This review will discuss the suitability of GPNMB as a target for cancer therapy by summarizing our current understanding of GPNMB expression in normal tissues, its role in cancer progression, and the current use of ADCs for the treatment of GPNMB expressing cancers. We will consider GPNMB in the broader context of several cancers; however, when possible, we will emphasize emerging literature regarding GPNMB and breast cancer.
Homology and structure of GPNMB
GPNMB, initially termed glycoprotein non-metastatic gene B (NMB), was first cloned and described in 1995 as a protein highly expressed in a melanoma cell line with low metastatic potential.Citation38 However, since this initial publication, elevated GPNMB expression is observed in numerous cancers and is often associated with the metastatic phenotype.Citation27–Citation34 GPNMB is also known as hematopoietic growth factor inducible, neurokinin-1 type (HGFIN),Citation39 and is located on the small arm of chromosome 7 (7p15). The rat orthologue, termed osteoactivin, is expressed in the long bones of rats bearing a mutation associated with osteopetrosis and shares 65% protein identity with human GPNMB.Citation40 The mouse orthologue, which has 71% protein identity with human GPNMB, was coined dendritic cell heparin integrin ligand (DC-HIL) following its identification in a particular subset of dendritic cells.Citation41
GPNMB belongs to the vertebrate Pmel17/NMB family,Citation42 which encompasses GPNMB, Pmel17 (melanocyte protein 17), and their orthologues. Pmel17 is the main structural component of melanosomes, where it plays a key role in the pigment biogenesis of melanocytes.Citation43 To a lesser extent, GPNMB also shares homology with lysosome-associated membrane protein (LAMP-1) family members,Citation41 which are glycoproteins with potential roles in cell adhesion and metastasis.Citation44
GPNMB is a type I transmembrane protein that contains an N-terminal signal peptide, an integrin-binding (RGD) motif and a polycystic kidney disease (PKD) domain in its extracellular domain (ECD), a single pass transmembrane domain, and a 53 amino acid (AA) cytoplasmic tailCitation45,Citation46 (). The cytoplasmic tail harbors a half immunoreceptor tyrosine-based activation motif (hemITAM) and a dileucine motif, which functions as a sorting signal in QNR-71, the quail orthologue of GPNMB.Citation47 In addition to these domains and motifs, there are two known splice variants of GPNMB, comprising a short 560aa and a long 572aa isoform.Citation30 The long isoform contains a 12aa insertion within a poorly conserved region downstream of the PKD-domain.Citation30 To date, there has been no evidence that the short and long isoforms have disparate functions. However, one study reported that the short GPNMB isoform was more frequently expressed in glioma specimens and was significantly correlated with poor survival times whereas the correlation between the long GPNMB isoform and survival times failed to achieve statistical significance.Citation30
Figure 1 A schematic representation of GPNMB indicating the domains and motifs contributing to GPNMB function.
Abbreviations: GPNMB, glycoprotein non-metastatic b; hemiTAM, immunoreceptor tyrosine-based activation motif; PKD, polycystic kidney disease; RGD, integrin-binding.
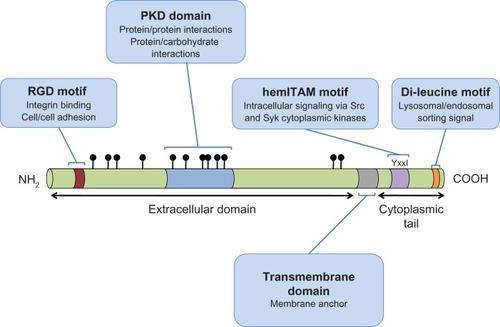
RGD domain
This motif, comprised of only 3 amino acids, arginine (R), glycine (G), and aspartic acid (D), is found near the N-terminus of the GPNMB ECD and is well characterized in numerous proteins as an integrin-binding motif.Citation48 Integrins are heterodimeric transmembrane proteins expressed on a wide variety of cells, which regulate cell spreading, adhesion, migration, proliferation, and apoptosis.Citation49
PKD domain
The PKD domain belongs to the immunoglobulin-(Ig) like fold superfamily (E-set), which also includes cadherins, protein families containing bacterial Ig-like domains, and several fibronectin type III domain-containing protein families. While the function of the PKD domain is still unclear, based on its structure, it has been proposed to mediate protein-protein or protein-carbohydrate interactions,Citation50 and has been shown to mediate cell-cell adhesion.Citation51
hemiTAM
ITAM (immunoreceptor tyrosine-based activation motif) motifs are commonly found in the cytoplasmic domains of receptors expressed by cells of the hematopoietic systems.Citation52 ITAM motifs are found in antigen receptors, cytokine receptors and toll-like receptors.Citation53 ITAM signaling usually occurs in response to ligand binding, via phosphorylation of the ITAM resident tyrosine residues, primarily by Src-family kinases (ie, Src, Hck, Fgr, Lyn).Citation54
GPNMB is one of several proteins whose cytoplasmic tail contains a highly conserved, single YxxI sequence, which has been referred to as a hemi-ITAM or hemITAM motif.Citation52 Proteins with hemITAMs still exhibit robust ITAM signaling capacity.Citation55 The current view suggests that ligand binding stimulates dimerization of hemITAM-bearing receptors; however, it remains to be seen whether GPNMB is capable of forming such homodimers.
Dileucine sorting motif
GPNMB contains a dileucine motif in its cytoplasmic tail, near the carboxy-terminus, with the sequence EKDPLL. Dileucine-based motifs of this type (D/ExxxLL) are often implicated in rapid receptor internalization from the plasma membrane and lysosomal/endosomal targeting.Citation56 Indeed, when either of these leucine residues is mutated to glycine in quail GPNMB, it is retained at the plasma membrane of HeLa or pigmented quail cells, and not routed to endosomes and lysosomes, as is the case for wild type GPNMB.Citation47 Interestingly, sequences of this type are associated with basolateral targeting in polarized epithelial cells.Citation56
Glycosylation
GPNMB is a heavily glycosylated protein, possessing 12 putative N-glycosylation sites within its extracellular domain, 6 of which are found in the PKD domain.Citation38,Citation45 Glycosidase treatments have confirmed that GPNMB can be N- and O-glycosylated in a variety of cell types.Citation30,Citation57,Citation58 Following immunoblot analyses, human GPNMB is detected as two broad bands that correspond to precursor (P1 ~90 kDa) and mature (M ~115 kDa) GPNMB isoforms.Citation58 In addition, the unglycosylated form of GPNMB (~65 kDa) has been detected in cells, such as osteoclasts (Sheng et al).Citation59 The relative abundance of these bands varies based on the cell type in which GPNMB is expressed.Citation28,Citation58 Studies using N-glycosidases suggest that GPNMB is first N-glycosylated in the ER to yield the P1 isoform, and these N-glycans are further modified during processing in the Golgi apparatus to produce the M-form.Citation58 While both isoforms are susceptible to tyrosine phosphorylation, only the mature form can be proteolytically processed through shedding (discussed below).Citation58,Citation60 A number of studies have linked the glycosylation status of GPNMB to its putative biological functions and will be addressed in the relevant sections described below.
Proteolytic cleavage and ECD shedding
GPNMB is also subject to proteolytic processing, which was first uncovered by the detection of two heavily glycosylated, high molecular weight forms of murine GPNMB (97 kDa, 116 kDa; discussed above) and a stable c-terminal fragment of ~20 kDa.Citation61 It was postulated that GPNMB was susceptible to shedding by members of the matrix metalloproteinase (MMP) family, such as A disintegrin and metalloproteinase (ADAMs), because treatment with a broad-spectrum inhibitor of MMPs (GM6001) reduced the degree to which GPNMB was shed.Citation61 Treatment with a calmodulin inhibitor (W7) or a protein kinase C activator (phorbol myristate acetate [PMA]) enhanced GPNMB shedding, further implicating the ADAMs, as these compounds have both been reported to enhance ADAM-10 and ADAM-17 activity, respectively.Citation58 In agreement with an important role for ADAMs in GPNMB processing, constitutive GPNMB shedding was observed in breast cancer cells and definitively characterized ADAM10 as a sheddase responsible for this cleavage event in breast cancer cells.Citation27 The potential functional implications for this shedding event will be discussed further in subsequent sections.
GPNMB expression and physiological functions in normal tissues
GPNMB mRNA has been detected in the long bones, calvaria, bone marrow, adipose, thymus, skin, placenta, heart, kidney, pancreas, lung, liver, and skeletal muscle;Citation39–Citation41 however, the precise expression patterns varied between these studies. It is also clear that GPNMB can be expressed in multiple cell types within a given tissue, which is evident by its expression in bone osteoblastsCitation40 and osteoclasts,Citation59 for example. Together, these studies clearly demonstrate that GPNMB is expressed in a wide range of tissues and suggest its involvement in a variety of physiological processes.
Brain
Recently, widespread expression of GPNMB has been described within the central nervous system, and shown to be largely specific to the microglia/macrophages of the neural parenchyma.Citation62 Furthermore, GPNMB expression has been described within motor neurons of normal brain tissue. Interestingly, GPNMB upregulation has been observed in the brains of rats following stroke.Citation63 It is also upregulated in the motor neurons and astrocytes of a mouse model of amyotrophic lateral sclerosis (ALS) as well as in the spinal cords of patients with this disease.Citation64 Its over-expression was cytoprotective in ALS-affected tissues; GPNMB secretion from astrocytes induced survival signals in neighboring motor neurons through the PI3K and MEK/Erk pathways and was thus able to ameliorate ALS-induced neuronal degeneration. In this context, GPNMB glycosylation was shown to be important for increasing motor neuron stability, as ubiquitin-mediated degradation of glycosylated GPNMB ultimately triggered motor neuron death.Citation64 However, the presence of GPNMB was cytotoxic in normal neural tissues, suggesting a possible role for GPNMB in maintaining neuronal homeostasis.Citation64
Skin
GPNMB expression in the basal layer of the skin, particularly in melanocytes, has been well documented.Citation58,Citation65–Citation68 During development, GPNMB exhibits a punctate pattern of expression consistent with melanoblast cell populations, which represent precursors of melanocytes.Citation65 In adult melanocytes, it preferentially localizes to late-stage (III and IV) melanosomes, which are characterized by an accumulation of the melanin pigment, suggesting a putative role for GPNMB in melanosome maturation.Citation58,Citation67 Its weak cell-surface expression can be upregulated following UVA irradiation, or stimulation by αMSH, IFNγ and TNFα.Citation67 A separate study showed that GPNMB can be upregulated by UVB and suggested that UVB-mediated formation of early melanosomes is mediated by GPNMB.Citation68 Functionally, GPNMB can mediate melanocyte adhesion to keratinocytes through its RGD domain and is thought to be involved in the transport of late melanosomes to keratinocytes.Citation58,Citation67 GPNMB can be localized at the cell surface and internalized into endosomal and lysosomal compartments in a variety of cell types. In melanocytes and melanoma cells, it is thought that the extensive glycosylation of the GPNMB PKD domain contributes to the differential sorting and localization patterns that are observed between GPNMB and its close homologue, Pmel.Citation69 Indeed, while the PKD domain of Pmel plays an active role in the cellular distribution of Pmel, the degree of glycosylation in GPNMB blocks this sorting function, leading to differential localization of GPNMB.Citation69
Bone
The first link between GPNMB expression and bone physiology was made when it was identified as highly expressed by mature, matrix producing rat osteoblasts in osteopetrotic bones relative to normal bone.Citation40 Subsequent studies have shown that antisense oligonucleotide- or neutralizing antibody-mediated inhibition of GPNMB in developing osteoblasts impairs their differentiation and decreases their ability to produce bone matrix.Citation70,Citation71 Recently, it has been shown that GPNMB addition to a critical-size bone defect model was able to support bone regeneration/formation.Citation72
In addition, GPNMB is abundantly expressed in differentiated osteoclastsCitation73 and was found to play an important role in mediating cell fusion to produce multi-nucleated osteoclasts.Citation59 GPNMB has been shown to physically associate with β1 or β3 containing integrin complexes in osteoclasts and to be an important mediator of osteoclast differentiation/fusion. Interestingly, it was the unglycosylated form of GPNMB that was found in complexes containing β1 or β3 integrins.Citation59 Indeed, neutralizing antibodies against GPNMB reduced osteoclast size and number and decreased their ability to resorb bone.Citation59 Additionally, transgenic mice expressing GPNMB under the control of a tartrate-resistant acid phosphatase (TRAP) promoter displayed evidence of significant bone loss and elevated bone resorption markers compared to non-transgenic controls.Citation74 Osteoclasts isolated from these transgenic mice were twice as large, possessed elevated TRAP activity, exhibited enhanced expression of osteoclast markers, and could resorb bone matrix to a greater degree than osteoclasts isolated from wild-type controls.Citation74 Thus, GPNMB is expressed and contributes to the differentiation and function of both osteoblasts and osteoclasts within the bone microenvironment.
Immune system
The molecular functions of GPNMB are just beginning to be elucidated and perhaps have been best characterized in the immune system. Expression of GPNMB has been detected in leukocytes and antigen presenting cells, including macrophagesCitation62,Citation75,Citation76 and dendritic cellsCitation41,Citation77 and has been involved in promoting various cell-cell interactions. GPNMB expression on dendritic cells has been shown to mediate their adhesion to endothelial cells through its RGD domain.Citation41 Additionally, the extracellular domain of GPNMB can suppress T-cell activation and proliferation by binding to syndecan-4 on the surface of activated T-cells, and this interaction requires an intact PKD domain.Citation78,Citation79 GPNMB binding to syndecan-4 leads to the recruitment of syntenin and the CD148 protein tyrosine phosphatase, the activation of which occurs following complex formation and is required for syndecan-4 mediated suppression of T-cell activation.Citation80 This ability to modulate adaptive immunity has been documented in a variety of contexts including graft versus host disease (GVHD), where GPNMB suppresses the activity of alloreactive T cells.Citation81
In contrast to these immunosuppressive roles, activation of GPNMB in dendritic cells, either by ligand binding or antibody cross-linking, can induce an innate immune response against fungal antigens. Under these conditions, the hemITAM tyrosine residue of GPNMB became phosphorylated, which induced widespread changes in gene and protein expression, including increased cytokine secretion (TNFα, IL-1β).Citation60 This activation of GPNMB stimulated dendritic cell maturation and augmented their ability to potentiate the activation of naive T-cells.Citation60 While these findings are strongly suggestive of functional hemITAM-based signaling in GPNMB, more research is needed to definitively characterize the role of this motif when GPNMB is expressed in immune or non-immune cells.
In models of cardiomyopathy, liver fibrosis, and kidney disease, increased GPNMB expression was observed in resident and infiltrating macrophages and is thought to serve as a compensatory response to promote tissue repair through autophagy and phagocytosis of cell debris.Citation75,Citation82–Citation84 During tissue repair, GPNMB localizes to LC3-positive lysosomes, which form during autophagy, and mediates degradation of cellular debris by promoting the fusion of autophagosomes to lysosomes.Citation82,Citation84
It is clear from these observations that GPNMB expression is widespread and it is able to regulate a wide range of physiological and pathological processes. Its established roles during normal tissue processes, such as adhesion during transendothelial migration of dendritic cells and autophagy during tissue repair, are also important mechanisms observed during cancer progression and metastasis. Intriguingly, GPNMB expression can be upregulated in pathological conditions, such as chronic liver disease, which can lead to carcinogenesis.Citation31 As discussed below, it is possible that GPNMB expression in infiltrating immune cells may play important roles in supporting the tumor microenvironment. Considering that the mechanisms of action for GPNMB in tumor progression have yet to be fully elucidated, these observations of GPNMB function in normal tissues represent compelling potential roles for GPNMB in cancer and warrant further investigation.
GPNMB and cancer
Tumor suppressive properties
While it has become increasingly clear that the initial designation of GPNMB as “glycoprotein non-metastatic gene B” is inaccurate in the context of melanoma (see below), there are cancers in which GPNMB appears to exert a tumor-suppressive response.
In the vast majority of colorectal carcinomas, GPNMB is epigenetically silenced by promoter methylation and could thus be involved in attenuating aggressiveness and delaying tumor progression.Citation85 Additionally, a recent study examining GPNMB over-expression in prostate carcinoma cell lines reported a reduction in invasion and proliferation in vitro and tumor growth in vivo.Citation86 Upregulation of anti-metastatic genes, including Ndrg1 and maspin, was observed following forced GPNMB expression in this model, and was proposed as a potential mechanism to explain the anti-tumorigenic effects associated with GPNMB expression.Citation86 These findings emphasize the complexity of GPNMB’s role in tumor biology and the need to obtain a more comprehensive understanding of its mechanisms of action.
Tumor promoting properties
Emerging data has generated a more complex picture with respect to GPNMB in cancer progression, and it is now evident that GPNMB can function to promote tumor progression in certain types of cancer and can act as a tumor suppressor in others.Citation46 The literature investigating the relationship between GPNMB and cancer continues to grow, with an increasing number of reports describing positive correlations between GPNMB expression, poor outcomes and pro-invasive/pro-metastatic phenotype in a variety of cancers.
GPNMB expression and function in breast cancer
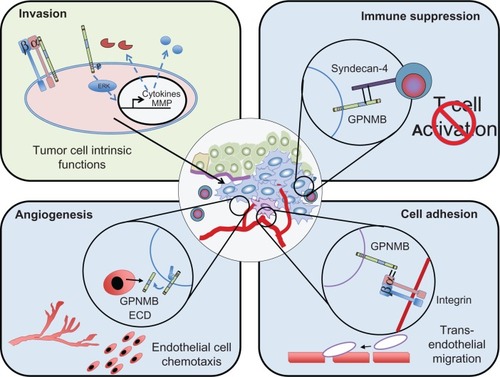
In a screen for metastatic modulators of breast cancer, GPNMB was identified as a gene that is frequently and highly expressed in aggressively metastatic breast cancer cell populations.Citation27,Citation29 Over-expression of GPNMB in weakly metastatic breast cancer cells was shown to drive the acquisition of an invasive phenotype in vitro, characterized by elevated MMP-3 levels, and enhance the bone metastatic potential of these cells.Citation29 A recent study looking at GPNMB over-expression in a murine mammary carcinoma model found that GPNMB could also promote primary mammary tumor growth.Citation27 GPNMB-expressing tumors were characterized by a high endothelial cell density compared to tumors that lacked GPNMB, and in vitro studies revealed that the soluble GPNMB ECD is biologically active as it was capable of inducing endothelial migration.Citation27 These data suggest that GPNMB could regulate the ability of breast cancer cells to recruit vasculature to permit tumor growth and metastasis. Combined, these observations reveal both tumor intrinsic effects of GPNMB that can enhance the invasiveness of tumor cells as well as numerous mechanisms through which GPNMB can facilitate interactions with, and influence the behavior of, cells within the tumor microenvironment to promote the growth and spread of cancer cells ().
Figure 2 Potential mechanisms through which GPNMB promotes malignant cellular phenotypes within cancer cells.
Abbreviations: ECD, extracellular domain; GPNMB, glycoprotein non-metastatic b.
In an independent study of GPNMB expression in breast cancer, where the authors employed in situ mRNA hybridization to detect GPNMB in human breast tumors, its expression was reported to be lower in tumors compared to normal tissues.Citation87 GPNMB was also found to be expressed at high levels in immortalized cell lines derived from normal breast epithelium and at low levels in breast cancer cell lines in this study. These studies are in opposition to other published findings and may reflect the fact that the authors did not take the breast cancer subtype into account during their analysis.Citation29,Citation88,Citation89
GPNMB expression and function in other solid malignancies
Brain cancer
The first association of GPNMB with cancer progression was in 2003, when it was reported to promote the invasion of glioma cells.Citation33 These pro-invasive effects were attributed to the ability of GPNMB to enhance the expression of MMP-3 and MMP-9.Citation33 Subsequent studies have confirmed that GPNMB expression is elevated in both benign subependymal giant cell astrocytomasCitation90 as well as malignant glioblastomas.Citation30 Importantly, glioblastoma patients with high levels of GPNMB transcript and protein levels were at significantly higher risk of death.Citation30
Melanoma
The notion that GPNMB is linked to melanomas with low-metastatic potentialCitation38 has been dispelled by subsequent studies that report high GPNMB expression in malignant cutaneous melanoma.Citation32,Citation91 In a murine melanoma model, it has been suggested that GPNMB promotes tumor growth via an immunosuppressive mechanism involving a block in T-cell activation.Citation92 Interestingly, this study also reported that GPNMB could be released from melanoma cells in the form of exosomes, and that this dissemination of GPNMB might facilitate systemic immunosuppression of anti-tumor responses.Citation92 It was in the context of cutaneous melanoma that anti-GPNMB therapies were first considered,Citation32,Citation93,Citation94 which is discussed in greater detail below. Interestingly, a recent survey of uveal melanomas revealed that a high percentage of these aggressive tumors also express GPNMB.Citation34
GPNMB function in tumor stroma
GPNMB expression in the stromal compartment of different cancers could also potentially be linked to tumor progression. GPNMB was over-expressed in a subset of CD10-positive cancer associated fibroblasts derived from colon tissue,Citation95 which is in line with previous reports that GPNMB can activate fibroblasts by inducing upregulation of pro-invasive matrix metalloproteases, such as MMP-3 and MMP-9, via Erk-dependent signaling.Citation61,Citation96 In macrophages, treatment with tumor-cell conditioned media induced an 83-fold increase in GPNMB expression.Citation97 Interestingly, these tumor-conditioned macrophages adopted a phenotype similar to the M2-type macrophages,Citation97 which are known for their role in promoting tumor progression.Citation98 In the breast, GPNMB expression is abundant in the stromal compartment of tumor tissue,Citation28 which could be attributed to its expression in a variety of stromal subtypes described above. Taken together, these studies suggest a role for GPNMB in sustaining the tumor microenvironment; however, it remains to be seen if stromal GPNMB can directly influence tumor progression. In this regard, it is interesting to note that GPNMB expression in the tumor epithelium of breast cancers was associated with poorer prognosis, whereas breast cancers that lacked GPNMB or displayed predominantly stromal GPNMB expression displayed better outcomes.Citation28 However, this may be a reflection of the fact that tumor-cell-intrinsic GPNMB expression is required for breast cancer progression and does not necessarily negate an important role for stromal-derived GPNMB in this disease.
GPNMB as a therapeutic target
Given the increasing association between GPNMB expression and a variety of cancers, and the acquisition of aggressive cellular phenotypes in GPNMB-expressing cancer cells, there has been growing interest in the development of GPNMB-targeted therapies.Citation35–Citation37 The pattern of GPNMB expression in normal and cancerous tissues makes it an intriguing target for cancer therapy. Generally speaking, GPNMB localization tends to be restricted to intracellular compartments in normal cells, such as macrophages, melanocytes and pigmented retinal epithelial cells.Citation67,Citation76,Citation99 In contrast, GPNMB expression in tumor cells is enriched on the cell surface.Citation28,Citation32,Citation94 This pattern of sub-cellular localization makes tumor-specific GPNMB more readily available for antibody targeting, thus providing a therapeutic window and making GPNMB a uniquely attractive target for antibody based therapies.
Targeting GPNMB in brain cancers
A single chain antibody coupled to an immunotoxin (F6V-PE38), which is directed against the extracellular domain of GPNMB, has recently been generated for the treatment of glioblastoma multiforme.Citation100 A GPNMB-specific single chain variable fragment (scFv) antibody was first isolated from a phage display library and subsequent mutagenesis/selection of this clone produced a high-affinity GPNMB-specific scFv antibody (F6V). This scFv was then conjugated to a truncated form of Pseudomonas endotoxin A to generate F6V-PE38, which causes protein synthesis inhibition and apoptosis following internalization by GPNMB-expressing target cells. Two xenograft models of malignant glioma (glioblastoma multiforme and medulloblastoma) were subjected to treatment with the anti-GPNMB immunotoxin (F6V-PE38), which resulted in a significant impairment in tumor growth compared to PBS-treated controls.Citation100 Although these findings are preliminary, they address the potential for development of small-size targeted therapeutics against GPNMB, which will penetrate the tumor mass with higher efficiency compared to full-length conjugated antibodies.Citation101
Targeting GPNMB in melanoma and breast cancer
A more developed GPNMB-targeted therapeutic agent is CDX-011, an antibody-drug conjugate also known as CR011-vcMMAE (CR011) or glembatumumab vedotin.Citation94 In the case of CDX-011, the cytotoxin auristatin E, a tubulin destabilizer, is conjugated to an antibody directed against the extracellular domain of GPNMB.Citation94 Upon GPNMB binding and internalization, the drug is released and induces cell cycle arrest and apoptosis of the target cell.
Pre-clinical models
The first evidence of successful therapeutic targeting of GPNMB using this ADC demonstrated that CDX-011 was selectively able to inhibit the growth of GPNMB-expressing metastatic melanoma cells, both in culture and xenograft assays.Citation94 A subsequent study examining the pharmacological properties of this antibody-drug conjugate showed that, at concentrations as low as 2.5 mg/kg, CDX-011 was capable of inducing complete regression in 100% of GPNMB-expressing xenografted SK-Mel-2 and SK-Mel-5 melanoma cells.Citation93 In breast cancer, a single dose of 20 mg/kg CDX-011 was sufficient to induce sustained MDA-MB-468 tumor regression in vivo.Citation28 Numerous studies have reported that cell killing efficacy of CDX-011 is directly proportional to the level of GPNMB expressed on the cell surface.Citation28,Citation32,Citation93,Citation94
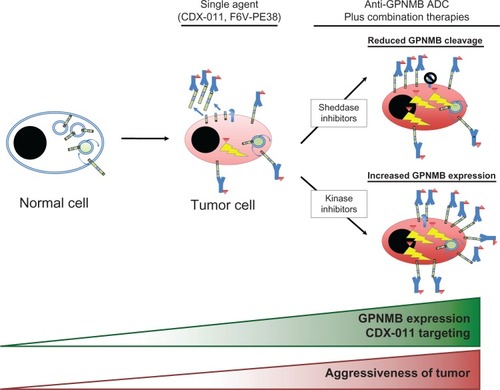
Interestingly, treatment of cancer cells with imatinib or inhibitors of the Erk pathway enhances cell surface expression of GPNMB in cancer cells, which in turn increases sensitivity to CDX-011.Citation32 Additionally, a separate study examining monocyte-derived dendritic cells (moDC) reported that BCR-ABL and Src family kinase inhibitors such as imatinib, dasatinib, and nilotinib increased GPNMB expression and thereby potentiated immune-suppression by moDCs.Citation102 Inhibitors of metalloproteinases, such as GM6001, have also been shown to enhance cell surface GPNMB expression by preventing shedding of its extracellular domain.Citation32,Citation61 In addition to increasing target availability, such inhibitors can minimize the potential for sequestration of CDX-011 by the shed form of GPNMB and thereby increase the targeted killing of GPNMB-expressing tumor cells. However, the effect of these inhibitors on tumor cell sensitivity to CDX-011 has not yet been examined. These findings suggest that combinations with additional targeted therapies (that are capable of enhancing cell surface GPNMB expression) could further enhance the efficacy of CDX-011. Given the pro-invasive and pro-metastatic functions of GPNMB, such a strategy would require careful evaluation in pre-clinical models to ensure that these combination therapies did not increase metastasis of cancer cells that escape CDX-011 mediated killing. ().
Figure 3 Therapeutic strategies employing anti-GPNMB antibody-drug conjugates (ADCs).
Abbreviations: ADC, antibody-drug conjugate; CDX-011, glembatumumab vedotin; GPNMB, glycoprotein non-metastatic b.
Clinical trials
CDX-011 was initially tested in two multi-centre phase I/II clinical trials; one for patients with unresectable melanomaCitation103 and the other for patients with locally advanced or metastatic breast cancer.Citation104,Citation105 Tumor shrinkage was reported in 56% of melanoma patients and 62% of breast cancer patients who were treated with a maximum tolerated dose (MTD) of 1.88 mg/kg once every 3 weeks.Citation103,Citation104 GPNMB expression appeared to be a predictive biomarker in the melanoma study. A small subset of melanoma patients with the highest levels of tumoral GPNMB expression (n = 7) had longer median progression-free survival (PFS) times (4.9 months) compared to the median PFS for all patients in the cohort (n = 34; including those with high tumoral GPNMB), which ranged from 1–3.9 months depending on the dose frequency.Citation103 This observation was recapitulated in a subset of breast cancer patients treated with CDX-011. In this study, the median PFS for GPNMB-positive patients (n = 9) was 17.3 weeks compared to 9.1 weeks for all patients (n = 34) treated with the MTD.Citation104 Interestingly, patients with strong GPNMB expression in stromal cells responded to CDX-011 just as well, if not better, than patients with strong GPNMB expression in the tumor epithelium.Citation104 It is conceivable that GPNMB-expressing cells that initially take up CDX-011 can release the drug moiety when the targeted cells die, which can freely diffuse into neighboring cells and kill them regardless of whether they expressed GPNMB. This “bystander” effect has been described with SGN-35, which is an antibody drug conjugate that targets CD30.Citation106
Based on these observations, a subsequent EMERGE (NCT01156753) phase IIb clinical trial was recently carried out to investigate the efficacy and safety of CDX-011 for patients with heavily pre-treated metastatic breast cancer that were positive for GPNMB.Citation104 The final results from this trial were presented at the 2012 San Antonio Breast Cancer Symposium and showed promise for CDX-011 treatment of patients with GPNMB-expressing and triple negative breast cancer.Citation107 The trial enrolled 122 patients and was carried out in a 2:1 randomized fashion where 81 patients received CDX-011 and 41 received investigator’s choice of therapy (IC). Patients treated with IC were allowed to crossover to CDX-011 therapy if they continued to be eligible after confirmation of pharmacodynamics. Eligible patients were required to have GPNMB expression in ≥5% of tumor epithelial and/or stromal tissue, as confirmed by immunohistochemistry on archived tumor samples. Patients were required to have been previously treated with all of the following therapeutic regimens, when indicated, prior to enrollment: taxane, anthracycline, capecitabine, traztuzumab, and lapatinib. Interestingly, 99% of patients tested displayed some level of tumoral GPNMB expression, which was significantly higher than earlier reports of GPNMB expression from breast cancer tissue microarrays.Citation28 To assess the potential for utilizing GPNMB as a predictive marker for CDX-011 therapy, patients were classified as having high or low GPNMB expression based on a threshold cutoff of ≥25% GPNMB positivity, post-hoc.Citation103 The trial reported that 41% of TNBC patients had high GPNMB expression, which was consistent with previous studies, and further confirmed GPNMB as a promising target in this aggressive disease subtype. Partial response was observed in 19% of patients with triple negative disease, compared to 0% with IC, which is an encouraging result for a subgroup of breast cancer patients with currently limited treatment options. The response rate was even higher (33% versus 0%) in the TNBC subset of patients displaying high GPNMB expression, substantiating findings from the melanoma phase I/II trial. Additionally, patients with high GPNMB expression and TNBC had a doubling in progression free survival (3.0 months [n = 12 patients receiving CDX-011] versus 1.5 months [n = 6 patients receiving IC]; P = 0.008) and overall survival (10.0 months [n = 12 patients receiving CDX-011] versus 5.5 months [n = 6 patients receiving IC]; P = 0.003). While the results are encouraging, it must be noted that the sample sizes in these groups are very small. Also, no statistically significant differences were observed across all subtypes between CDX-011 and IC treated patients with high GPNMB expression. However, contrary to reports from previous trials,Citation104 stromal GPNMB expression did not appear to be a predictive marker of response to therapy in the EMERGE study.
In these studies, development of a skin rash was one of the most common side effects experienced by melanoma (57%) and breast cancer patients treated with CDX-011 (48%, 47%).Citation103,Citation104,Citation107 This finding was of great interest, given that GPNMB is expressed in the skin.Citation65,Citation67 Interestingly, melanoma patients who experienced rash within their first cycle of treatment also had significantly longer PFS than CDX-011-treated patients who didn’t develop rash (4.8 versus 1.2 months; P < 0.001), suggesting that rash may be an early indicator of a patient’s ability to tolerate and respond to the drug.Citation103 Additional side effects in the EMERGE study that were worsened in patients treated with CDX-011 compared to IC include other dermatological conditions such as alopecia (hair loss) and pruritus (itch) as well as peripheral neuropathy and vomiting.Citation107 However, patients undergoing CDX-011 therapy witnessed a reduction in hematologic side effects such as neutropenia, leucopenia, and thrombocytopenia. Although GPNMB is largely expressed in intracellular compartments in normal tissues, CDX-011 can adversely affect certain tissue types, which is evident by its ability to induce skin rash.
One tissue that could be susceptible to side effects of CDX-011 treatment includes the bone. The potential use of CDX-011 to target breast cancer bone metastases should be met with caution. Osteoblasts and osteoclasts both express cell-surface localized GPNMB and their targeting by CDX-011 could have detrimental effects on bone turnover. Decreased osteoblast numbers would reduce bone formation, which could lead to an increased risk of fracture for the patients. Conversely, targeted killing of osteoclasts could delay bone healing and lead to osteopetrosis. Indeed, antibodies directed against GPNMB were shown to impair osteoclast formation and function.Citation59 Bone remodeling is a finely-tuned process and tipping the scale in either direction could exacerbate the side effects of CDX-011. These considerations should be kept in mind when choosing patient cohorts for CDX-011 treatment. Overall, in light of the scarcity of treatment options for TNBC patients, these data substantiate further studies investigating the efficacy of CDX-011 in the treatment of metastatic breast cancer.
Conclusions and future perspectives
The development of antibody-based therapeutic agents targeting GPNMB (single chain variable fragment antibodies, antibody drug conjugates) is a promising avenue for several GPNMB-expressing cancers. The latest phase II clinical trial data reinforces the early results from phase I/II trials, which supports the use of CDX-011 in women with triple negative breast cancer. Early efforts to identify potential therapeutic combinations that will increase the efficacy of anti-GPNMB agents will need to be investigated with caution. Enhancing cell surface expression of GPNMB may sensitize tumor cells to more effective killing by agents such as CDX-011; however, the acquisition of malignant phenotypes in cancer cells with elevated levels of GPNMB expression, which are not eliminated, is cause for concern.
A better understanding of the molecular mechanisms through which GPNMB induces aggressive cellular phenotypes, such as enhanced migration and invasion, will be needed in order to fully optimize therapeutic molecules targeting GPNMB. Another aspect that requires further investigation is the contribution of stromal cells within the tumor microenvironment that express GPNMB and how this impacts tumor progression and response to anti-GPNMB therapies.
Acknowledgments
The authors would like to thank Dr Josie Ursini-Siegel and members of the Siegel laboratory for their thoughtful and insightful comments on the manuscript. GM acknowledges studentship support from the Canadian Institutes for Health Research (CIHR). Research conducted in the author’s laboratory, cited in this review, was supported by grants from the CIHR (MOP-119401).
Disclosure
The authors report no conflict of interests in this work.
References:
- ForouzanfarMHForemanKJDelossantosAMBreast and cervical cancer in 187 countries between 1980 and 2010: a systematic analysisLancet201137898011461148421924486
- JemalABrayFCenterMMFerlayJWardEFormanDGlobal cancer statisticsCA Cancer J Clin2011612699021296855
- PerouCMSørlieTEisenMBMolecular portraits of human breast tumoursNature2000406679774775210963602
- SorlieTPerouCMTibshiraniRGene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implicationsProc Natl Acad Sci USA20019819108691087411553815
- SorlieTTibshiraniRParkerJRepeated observation of breast tumor subtypes in independent gene expression data setsProc Natl Acad Sci USA2003100148418842312829800
- HerschkowitzJISiminKWeigmanVJIdentification of conserved gene expression features between murine mammary carcinoma models and human breast tumorsGenome Biol200785R7617493263
- PratAParkerJSKarginovaOPhenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancerBreast Cancer Res2010125R6820813035
- CurtisCShahSPChinSFThe genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroupsNature2012486740334635222522925
- Nik-ZainalSAlexandrovLBWedgeDCMutational processes molding the genomes of 21 breast cancersCell2012149597999322608084
- Nik-ZainalSVan LooPWedgeDCThe life history of 21 breast cancersCell20121495994100722608083
- ShahSPRothAGoyaRThe clonal and mutational evolution spectrum of primary triple-negative breast cancersNature2012486740339539922495314
- StephensPJTarpeyPSDaviesHThe landscape of cancer genes and mutational processes in breast cancerNature2012486740340040422722201
- WangYKlijnJGZhangYGene-expression profiles to predict distant metastasis of lymph-node-negative primary breast cancerLancet2005365946067167915721472
- LargillierRFerreroJMDoyenJPrognostic factors in 1,038 women with metastatic breast cancerAnn Oncol200819122012201918641006
- SihtoHLundinJLundinMBreast cancer biological subtypes and protein expression predict for the preferential distant metastasis sites: a nationwide cohort studyBreast Cancer Res2011135R8721914172
- SmidMWangYZhangYSubtypes of breast cancer show preferential site of relapseCancer Res20086893108311418451135
- CheangMCChiaSKVoducDKi67 index, HER2 status, and prognosis of patients with luminal B breast cancerJ Natl Cancer Inst20091011073675019436038
- HuZFanCOhDSThe molecular portraits of breast tumors are conserved across microarray platformsBMC Genomics200679616643655
- CareyLADeesECSawyerLThe triple negative paradox: primary tumor chemosensitivity of breast cancer subtypesClin Cancer Res20071382329233417438091
- HughJHansonJCheangMCBreast cancer subtypes and response to docetaxel in node-positive breast cancer: use of an immunohistochemical definition in the BCIRG 001 trialJ Clin Oncol20092781168117619204205
- NielsenTOParkerJSLeungSA comparison of PAM50 intrinsic subtyping with immunohistochemistry and clinical prognostic factors in tamoxifen-treated estrogen receptor-positive breast cancerClin Cancer Res201016215222523220837693
- RouzierRPerouCMSymmansWFBreast cancer molecular subtypes respond differently to preoperative chemotherapyClin Cancer Res200511165678568516115903
- HillerDJChuQDCurrent Status of Poly(ADP-ribose) Polymerase Inhibitors as Novel Therapeutic Agents for Triple-Negative Breast CancerInt J Breast Cancer2012201282931522295252
- TuttARobsonMGarberJEOral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and advanced breast cancer: a proof-of-concept trialLancet2010376973723524420609467
- BalmanaJDomchekSMTuttAGarberJEStumbling blocks on the path to personalized medicine in breast cancer: the case of PARP inhibitors for BRCA1/2-associated cancersCancer Discov201111293422586318
- AdairJRHowardPWHartleyJAWilliamsDGChesterKAAntibody-drug conjugates – a perfect synergyExpert Opin Biol Ther20121291191120622650648
- RoseAAAnnisMGDongZADAM10 releases a soluble form of the GPNMB/Osteoactivin extracellular domain with angiogenic propertiesPLoS One201058e1209320711474
- RoseAAGrossetAADongZGlycoprotein nonmetastatic B is an independent prognostic indicator of recurrence and a novel therapeutic target in breast cancerClin Cancer Res20101672147215620215530
- RoseAAPepinFRussoCAbou KhalilJEHallettMSiegelPMOsteoactivin promotes breast cancer metastasis to boneMol Cancer Res20075101001101417951401
- KuanCTWakiyaKDowellJMGlycoprotein nonmetastatic melanoma protein B, a potential molecular therapeutic target in patients with glioblastoma multiformeClin Cancer Res2006127 Pt 11970198216609006
- OnagaMIdoAHasuikeSOsteoactivin expressed during cirrhosis development in rats fed a choline-deficient, L-amino acid-defined diet, accelerates motility of hepatoma cellsJ Hepatol200339577978514568261
- QianXMillsETorgovMLaRochelleWJJeffersMPharmacologically enhanced expression of GPNMB increases the sensitivity of melanoma cells to the CR011-vcMMAE antibody-drug conjugateMol Oncol200821819319383330
- RichJNShiQHjelmelandMBone-related genes expressed in advanced malignancies induce invasion and metastasis in a genetically defined human cancer modelJ Biol Chem200327818159511595712590137
- WilliamsMDEsmaeliBSoheiliAGPNMB expression in uveal melanoma: a potential for targeted therapyMelanoma Res6201020318419020375921
- KeirCHVahdatLTThe use of an antibody drug conjugate, glembatumumab vedotin (CDX-011), for the treatment of breast cancerExpert Opin Biol Ther201212225926322229970
- NaumovskiLJunutulaJRGlembatumumab vedotin, a conjugate of an anti-glycoprotein non-metastatic melanoma protein B mAb and monomethyl auristatin E for the treatment of melanoma and breast cancerCurr Opin Mol Ther201012224825720373269
- ZhouLTLiuFYLiYPengYMLiuYHLiJGpnmb/osteoactivin, an attractive target in cancer immunotherapyNeoplasma20125911522017590
- WetermanMAAjubiNvan DinterIMnmb, a novel gene, is expressed in low-metastatic human melanoma cell lines and xenograftsInt J Cancer199560173817814155
- BandariPSQianJYehiaGHematopoietic growth factor inducible neurokinin-1 type: a transmembrane protein that is similar to neurokinin 1 interacts with substance PRegul Pept20031111–316917812609765
- SafadiFFXuJSmockSLRicoMCOwenTAPopoffSNCloning and characterization of osteoactivin, a novel cDNA expressed in osteoblastsJ Cell Biochem2001841122611746512
- ShikanoSBonkobaraMZukasPKAriizumiKMolecular cloning of a dendritic cell-associated transmembrane protein, DC-HIL, that promotes RGD-dependent adhesion of endothelial cells through recognition of heparan sulfate proteoglycansJ Biol Chem2001276118125813411114299
- TurqueNDenhezFMartinPCharacterization of a new melanocyte-specific gene (QNR-71) expressed in v-myc-transformed quail neuroretinaEMBO J19961513333833508670835
- YamaguchiYHearingVJPhysiological factors that regulate skin pigmentationBiofactors200935219319919449448
- SaitohOWangWCLotanRFukudaMDifferential glycosylation and cell surface expression of lysosomal membrane glycoproteins in sublines of a human colon cancer exhibiting distinct metastatic potentialsJ Biol Chem19922678570057111544942
- SelimAAOsteoactivin bioinformatic analysis: prediction of novel functions, structural features, and modes of actionMed Sci Monit2009152MT19MT3319179976
- SinghMDel Carpio-CanoFBelcherJYFunctional roles of osteoactivin in normal and disease processesCrit Rev Eukaryot Gene Expr201020434135721395506
- Le BorgneRPlanqueNMartinPDewitteFSauleSHoflackBThe AP-3-dependent targeting of the melanosomal glycoprotein QNR-71 requires a di-leucine-based sorting signalJ Cell Sci2001114Pt 152831284111683416
- BarczykMCarracedoSGullbergDIntegrinsCell Tissue Res2010339126928019693543
- TakadaYYeXSimonSThe integrinsGenome Biol20078521517543136
- WestonBSMalhasANPriceRGStructure-function relationships of the extracellular domain of the autosomal dominant polycystic kidney disease-associated protein, polycystin-1FEBS Lett20035381–381312633844
- Ibraghimov-BeskrovnayaOBukanovNODonohueLCDackowskiWRKlingerKWLandesGMStrong homophilic interactions of the Ig-like domains of polycystin-1, the protein product of an autosomal dominant polycystic kidney disease gene, PKD1Hum Mol Genet20009111641164910861291
- KerriganAMBrownGDSyk-coupled C-type lectin receptors that mediate cellular activation via single tyrosine based activation motifsImmunol Rev2010234133535220193029
- IvashkivLBCross-regulation of signaling by ITAM-associated receptorsNat Immunol200910434034719295630
- MocsaiARulandJTybulewiczVLThe SYK tyrosine kinase: a crucial player in diverse biological functionsNat Rev Immunol201010638740220467426
- BradshawJMThe Src, Syk, and Tec family kinases: distinct types of molecular switchesCell Signal20102281175118420206686
- BonifacinoJSTraubLMSignals for sorting of transmembrane proteins to endosomes and lysosomesAnnu Rev Biochem20037239544712651740
- AbdelmagidSMBarbeMFRicoMCOsteoactivin, an anabolic factor that regulates osteoblast differentiation and functionExp Cell Res2008314132334235118555216
- HoashiTSatoSYamaguchiYPasseronTTamakiKHearingVJGlycoprotein nonmetastatic melanoma protein b, a melanocytic cell marker, is a melanosome-specific and proteolytically released proteinFASEB J20102451616162920056711
- ShengMHWergedalJEMohanSLauKHOsteoactivin is a novel osteoclastic protein and plays a key role in osteoclast differentiation and activityFEBS Lett2008582101451145818381073
- ChungJSYudateTTomihariMAkiyoshiHCruzPDJrAriizumiKBinding of DC-HIL to dermatophytic fungi induces tyrosine phosphorylation and potentiates antigen presenting cell functionJ Immunol200918385190519819794069
- FurochiHTamuraSMameokaMOsteoactivin fragments produced by ectodomain shedding induce MMP-3 expression via ERK pathway in mouse NIH-3T3 fibroblastsFEBS Lett2007581305743575018036345
- HuangJJMaWJYokoyamaSExpression and immunolocalization of Gpnmb, a glioma-associated glycoprotein, in normal and inflamed central nervous systems of adult ratsBrain Behav201222859622574278
- BugaAMScholzCJKumarSIdentification of new therapeutic targets by genome-wide analysis of gene expression in the ipsilateral cortex of aged rats after strokePLoS One2012712e5098523251410
- TanakaHShimazawaMKimuraMThe potential of GPNMB as novel neuroprotective factor in amyotrophic lateral sclerosisSci Rep2012257322891158
- LoftusSKAntonellisAMateraIGpnmb is a melanoblast-expressed, MITF-dependent genePigment Cell Melanoma Res20092219911018983539
- OwenTASmockSLPrakashSIdentification and characterization of the genes encoding human and mouse osteoactivinCrit Rev Eukaryot Gene Expr2003132–420522014696968
- TomihariMHwangSHChungJSCruzPDJrAriizumiKGpnmb is a melanosome-associated glycoprotein that contributes to melanocyte/keratinocyte adhesion in a RGD-dependent fashionExp Dermatol200918758659519320736
- ZhangPLiuWZhuCSilencing of GPNMB by siRNA inhibits the formation of melanosomes in melanocytes in a MITF-independent fashionPLoS One201278e4295522912767
- TheosACWattBHarperDCThe PKD domain distinguishes the trafficking and amyloidogenic properties of the pigment cell protein PMEL and its homologue GPNMBPigment Cell Melanoma Res Epub342013
- AbdelmagidSMBarbeMFArango-HisijaraIOwenTAPopoffSNSafadiFFOsteoactivin acts as downstream mediator of BMP-2 effects on osteoblast functionJ Cell Physiol20072101263717034042
- SelimAAAbdelmagidSMKanaanRAAnti-osteoactivin antibody inhibits osteoblast differentiation and function in vitroCrit Rev Eukaryot Gene Expr2003132–426527514696973
- BatemanJPSafadiFFSusinCWikesjoUMExploratory study on the effect of osteoactivin on bone formation in the rat critical-size calvarial defect modelJ Periodontal Res201247224324722097911
- RipollVMMeadowsNARaggattLJMicrophthalmia transcription factor regulates the expression of the novel osteoclast factor GPNMBGene20084131–2324118313864
- ShengMHWergedalJEMohanSAmouiMBaylinkDJLauKHTargeted overexpression of osteoactivin in cells of osteoclastic lineage promotes osteoclastic resorption and bone loss in micePLoS One201274e3528022536365
- RamachandranPPellicoroAVernonMADifferential Ly-6C expression identifies the recruited macrophage phenotype, which orchestrates the regression of murine liver fibrosisProc Natl Acad Sci USA201210946E3186E319523100531
- RipollVMIrvineKMRavasiTSweetMJHumeDAGpnmb is induced in macrophages by IFN-gamma and lipopolysaccharide and acts as a feedback regulator of proinflammatory responsesJ Immunol2007178106557656617475886
- AhnJHLeeYJeonCIdentification of the genes differentially expressed in human dendritic cell subsets by cDNA subtraction and microarray analysisBlood200210051742175412176896
- ChungJSDoughertyICruzPDJrAriizumiKSyndecan-4 mediates the coinhibitory function of DC-HIL on T cell activationJ Immunol200717995778578417947650
- ChungJSSatoKDoughertyIICruzPDJrAriizumiKDC-HIL is a negative regulator of T lymphocyte activationBlood2007109104320432717284525
- ChungJSCruzPDJrAriizumiKInhibition of T-cell activation by syndecan-4 is mediated by CD148 through protein tyrosine phosphatase activityEur J Immunol20114161794179921469128
- ChungJSTomihariMTamuraKKojimaTCruzPDJrAriizumiKThe DC-HIL ligand syndecan-4 is a negative regulator of T-cell allo-reactivity responsible for graft-versus-host diseaseImmunology2013138217318223113638
- LiBCastanoAPHudsonTEThe melanoma-associated transmembrane glycoprotein Gpnmb controls trafficking of cellular debris for degradation and is essential for tissue repairFASEB J201024124767478120709912
- PahlMVVaziriNDYuanJAdlerSGUpregulation of monocyte/macrophage HGFIN (Gpnmb/Osteoactivin) expression in end-stage renal diseaseClin J Am Soc Nephrol201051566119833906
- Patel-ChamberlinMWangYSatirapojBHematopoietic growth factor inducible neurokinin-1 (Gpnmb/Osteoactivin) is a biomarker of progressive renal injury across speciesKidney Int201179101138114821389974
- MokarramPKumarKBrimHDistinct high-profile methylated genes in colorectal cancerPLoS One200949e701219750230
- TsuiKHChangYLFengTHChangPLJuangHHGlycoprotein transmembrane nmb: an androgen-downregulated gene attenuates cell invasion and tumorigenesis in prostate carcinoma cellsProstate201272131431144222290289
- MetzRLPatelPSHameedMBryanMRameshwarPRole of human HGFIN/nmb in breast cancerBreast Cancer Res200795R5817845721
- NeveRMChinKFridlyandJA collection of breast cancer cell lines for the study of functionally distinct cancer subtypesCancer Cell200610651552717157791
- RoseAASiegelPMOsteoactivin/HGFIN: is it a tumor suppressor or mediator of metastasis in breast cancer?Breast Cancer Res20079640318086324
- TyburczyMEKotulskaKPokarowskiPNovel proteins regulated by mTOR in subependymal giant cell astrocytomas of patients with tuberous sclerosis complex and new therapeutic implicationsAm J Pathol201017641878189020133820
- ZhaoYQiaoZGShanSJSunQMZhangJZExpression of glycoprotein non-metastatic melanoma protein B in cutaneous malignant and benign lesions: a tissue microarray studyChin Med J (Engl)2012125183279328222964323
- TomihariMChungJSAkiyoshiHCruzPDJrAriizumiKDC-HIL/glycoprotein Nmb promotes growth of melanoma in mice by inhibiting the activation of tumor-reactive T cellsCancer Res201070145778578720570888
- PollackVAAlvarezETseKFTreatment parameters modulating regression of human melanoma xenografts by an antibody-drug conjugate (CR011-vcMMAE) targeting GPNMBCancer Chemother Pharmacol200760342343517541593
- TseKFJeffersMPollackVACR011, a fully human monoclonal antibody-auristatin E conjugate, for the treatment of melanomaClin Cancer Res20061241373138216489096
- GhilardiCChiorinoGDossiRNagyZGiavazziRBaniMIdentification of novel vascular markers through gene expression profiling of tumor-derived endotheliumBMC Genomics2008920118447899
- OgawaTNikawaTFurochiHOsteoactivin upregulates expression of MMP-3 and MMP-9 in fibroblasts infiltrated into denervated skeletal muscle in miceAm J Physiol Cell Physiol20052893C697C70716100390
- SolinasGSchiareaSLiguoriMTumor-conditioned macrophages secrete migration-stimulating factor: a new marker for M2-polarization, influencing tumor cell motilityJ Immunol2010185164265220530259
- JoyceJAPollardJWMicroenvironmental regulation of metastasisNat Rev Cancer20099423925219279573
- BächnerDSchröderDGrossGmRNA expression of the murine glycoprotein (transmembrane) nmb (Gpnmb) gene is linked to the developing retinal pigment epithelium and irisBrain Res Gene Expr Patterns200213–415916512638126
- KuanCTWakiyaKKeirSTAffinity-matured anti-glycoprotein NMB recombinant immunotoxins targeting malignant gliomas and melanomasInt J Cancer2011129111112120824708
- YokotaTMilenicDEWhitlowMSchlomJRapid tumor penetration of a single-chain Fv and comparison with other immunoglobulin formsCancer Res19925212340234081596900
- SchwarzbichMAGutknechtMSalihJThe immune inhibitory receptor osteoactivin is upregulated in monocyte-derived dendritic cells by BCR-ABL tyrosine kinase inhibitorsCancer Immunol Immunother201261219320221874302
- HamidOSnzolMPavlickACFrequent dosing and GPNMB expression with CDX-011 (CR011-vcMMAE), an antibody-drug conjugate (ADC), in patients with advanced melanomaJ Clin Oncol (Meeting Abstracts)20102815s8525
- SalehMNBendellJCRoseACorrelation of GPNMB expression with outcome in breast cancer (BC) patients treated with the antibody-drug conjugate (ADC), CDX-011 (CR011-vcMMAEJ Clin Oncol20102815s1095
- BurrisHSalehMNBendellJA Phase (Ph) I/II Study of CR011-VcMMAE, an Antibody-Drug Conjugate, in Patients (Pts) with Locally Advanced or Metastatic Breast Cancer (MBC)Cancer Res20096924s36096
- OkeleyNMMiyamotoJBZhangXIntracellular activation of SGN-35, a potent anti-CD30 antibody-drug conjugateClin Cancer Res201016388889720086002
- YardleyDAWeaverRMeliskoMA randomized phase 2 study of the antibody-drug conjugate CDX-011 in advanced GPNMB-overexpressing breast cancer: The EMERGE studyCancer Res20127224s3P6-10-01