Abstract
Background
The present study aims to investigate the subunit expression and enzyme activity of ribonucleotide reductase in cervical cancer patients, and detect the combined effect of the ribonucleotide reductase inhibitor gemcitabine and the chemotherapeutic agent carboplatin on cervical cancer cell lines.
Methods
Using quantitative reverse transcription polymerase chain reaction, Western blotting, and cytidine 5′-diphosphate reduction assays, we tested the expression and activity of ribonucleotide reductase in cervical cancer patients. The antitumor activity of gemcitabine and/or carboplatin treatments to SiHa and CaSki human cervical cancer cell lines were assessed by Cell Counting Kit-8 viability assay, EdU incorporation assay, immunofluorescence assay, flow cytometry assay, and Western blotting methods. Additionally, synergistic efficacy was quantitatively analyzed using a combination index based on the Chou-Talalay method.
Results
The mRNA levels of three ribonucleotide reductase subunits were all upregulated in the cervical cancer tissues compared with normal tissues (P<0.0001). Consistently, the protein expression and enzyme activity of ribonucleotide reductase were also increased in the cervical cancer tissues. Interestingly, gemcitabine inhibited DNA synthesis and carboplatin induced DNA damage. Further, the combined drug regime had a significant synergistic effect on inhibiting cervical cancer cell viability (log10[combination index] <0) via enhanced DNA damage and cell apoptosis.
Conclusion
The expression and activity of ribonucleotide reductase was increased in cervical cancer. Our study demonstrated the synergistic cytotoxicity of gemcitabine and carboplatin, through inhibiting DNA synthesis and increasing cell apoptosis in cervical cancer cell lines. This evidence might provide a rational clue of their combined application to improve cervical cancer treatment.
Introduction
Cervical cancer remains a serious health problem, with an annual incidence of 530,000 cases and 275,100 deaths in females worldwide.Citation1 More than 80% of cervical cancer is squamous cell carcinoma in pathologic classification.Citation1 Surgery is the main treatment for small stage I cervical cancers, but for patients with advanced, recurrent, or metastatic cervical cancer, chemotherapy remains the standard treatment.Citation2 Platinum compounds form bifunctional crosslinks with DNA, and DNA interstrand or intrastrand crosslinks induce DNA damage, including double-strand DNA breaks, and ultimately result in cell death.Citation3 Historically, cisplatin is regarded as the primary chemotherapy for advanced or metastatic cervical cancer, but the response rate is approximately 20%.Citation4 Carboplatin has activity in cervical cancer comparable with that of cisplatin, but has better chemical stability and less systemic toxicity.Citation5 With the encouraging Japan Clinical Oncology Group data, carboplatin may ultimately replace cisplatin in future practice and/or clinical trials.Citation6 A major clinical problem associated with platinum therapy is that although many patients may be initially responsive to it, they often relapse and become refractory to further single-agent platinum treatment.Citation7
The formation and growth of cancer cells are related to the synthesis of the DNA. Ribonucleotide reductase (RR) is an enzyme that catalyzes the reduction of ribonucleotides to their corresponding deoxyribonucleotides, which are the building blocks for DNA synthesis and repair in all living cells.Citation8 Human RR is composed of the catalytic large subunit RRM1 and the regulatory small subunit RRM2 or RRM2B.Citation8 Because RR plays a critical role in DNA synthesis, it is considered an important therapeutic target for cancer chemotherapy.Citation9
Gemcitabine (2,2-difluorodeoxycytidine; dFdC), currently the most potent RR inhibitor, has been widely used in treatment and assessment of the clinical benefit of different therapeutic approaches and combinations with other anticancer drugs or radiation therapy for solid tumors, including non-small cell lung cancer, pancreatic cancer, breast, ovarian, bladder, and head and neck cancer, as well as hematologic malignancies.Citation10 The compound dFdC is metabolized intracellularly to generate 5′-diphosphate (dFdCDP) and 5′-triphosphate (dFdCTP) nucleosides. While dFdCDP binds to RRM1 and inhibits RR activity, causing a reduction of the cellular dNTP concentration, dFdCTP competes with natural dCTP for incorporation into the replicating DNA, leading to DNA strand termination. The decrease of intracellular dCTP accelerates phosphorylation of dFdC to its two active forms, reduces metabolic clearance of gemcitabine nucleotides, and enhances incorporation of dFdCTP into DNA. This self-potentiation mechanism should account for the high anticancer efficacy of gemcitabine.Citation11,Citation12
Combination therapy based on drugs with different mechanisms of action is a major strategy for improving drug responses and cure rates and for overcoming resistance. Platinum agents and gemcitabine are ideal candidates for use in combination regimens because of their different but complementary biochemical mechanisms of action, similar antitumor activity profiles, and nonoverlapping side effect profiles.Citation13 There is clinical evidence indicating that the combination of platinum agents and gemcitabine improves response rates when compared with single platinum agents.Citation14,Citation15 However, the effect and mechanism of action of their combination has not been experimentally investigated previously in cervical cancer.
In this study we examined the expression and enzyme activity of three subunits of RR in patients with cervical cancer, and explored the combined effect of the RR inhibitor gemcitabine and the chemotherapeutic agent carboplatin on cervical cancer cells.
Materials and methods
Drugs, cell lines, and clinical tissue samples
Gemcitabine (Gemzar®) was purchased from Eli Lilly France (Fegersheim, France). Carboplatin (Paraplatin®) was obtained from Bristol-Myers Squibb Srl (Latina, Italy). SiHa and CaSki human cervical cancer cell lines (American Type Culture Collection, Manassas, VA, USA) were maintained in RPMI-1640 medium (Gibco, Carlsbad CA, USA) supplemented with 10% fetal bovine serum, 2 mM L-glutamine, and 100 U/mL penicillin-streptomycin at 37°C in a humidified atmosphere of 5% CO2. Paired surgical specimens of cancer and adjacent normal tissues were collected from 45 patients with cervical cancer at NingBo Women and Children’s Hospital from 2011 to 2012 following approval from The Scientific Research Institutional Review Board of Ningbo Women and Children’s Hospital. Tissue samples from patients with cervical cancer who received preoperative radiation or chemotherapy were excluded. All tissues were stored at −80°C immediately after excision.
RNA extraction and quantitative RT-PCR
Total RNA was isolated from clinical tissue samples with a total RNA isolation kit (AP-MN-MS-RNA, Axygen Scientific Inc., Union City, CA, USA) as described by the manufacturer. Single-strand complementary DNA was reverse-transcribed from 450 ng of total RNA using a first-strand complementary DNA synthesis kit (Takara Bio Inc., Shiga, Japan). Quantitative reverse transcription polymerase chain reaction (RT-PCR) was performed on an ABI Prism 7500 system (Applied Biosystems, Foster City, CA, USA) using SYBR Green Premix Ex Taq (TaKaRa Bio Inc., Shiga, Japan). The primers were as follows: RRM1 (sense, 5′-GGA GAC TAA TCA GGA CTG GT-3′, antisense, 5′-CAA CTT TGC GGA CAC GAC CT-3′), RRM2 (sense, 5′-GCA GCA AGC GAT GGC ATA GT-3′, antisense, 5′-GGG CTT CTG TAA TCT GAA CTT C-3′), and RRM2B (sense, 5′-TAA ACA GGC ACA GGC TTC CT-3′, antisense, 5′-CGC TCC ACC AAA TTT TCA TT-3′). For thermal cycling, the following conditions were applied: 10 minutes at 95°C, then 40 cycles of 15 seconds at 95°C and 2 minutes at 60°C. β-Actin was used as the loading control.
Western blotting analysis
Total tissue proteins were extracted as follows: clinical tissue specimens were triturated in buffer (50 mM Tris-HCl, pH 7.6, 2 mM DTT, and protease inhibitors) and centrifuged at 16,000 g and 4°C for 30 minutes. The supernatant was then collected for 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and Western blotting analysis. Primary antibodies for RRM1, RRM2, and RRM2B (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were used at 1:1,000 dilutions. GAPDH (Santa Cruz) antibody was used as a loading control. Secondary antibodies were purchased from Li-Cor Biosciences (Lincoln, NE, USA), and the blots were visualized and quantified using a Li-Cor Odyssey Infrared Imaging scanner.
RR activity assay
RR activity in clinical tissue samples was assayed based on the Steeper and Steuart CDP reduction methodCitation16 with modifications as described previouslyCitation17,Citation18 Briefly, the protein supernatants of total tissue lysates were collected as indicated above. Sequentially, 1% streptomycin sulfate precipitation and 40% saturated (NH4)2SO4 sedimentation were performed and the supernatants obtained were ultrafiltered through Amicon® ultra-15 centrifugal filters (Millipore, MA, USA) with 50 mM HEPES, pH 7.4, and 100 mM KCl. After quantification, the protein samples were added to 100 μL of reaction mixture containing 0.125 mM [3H] CDP (24 Ci/mmol), 50 mM HEPES (pH 7.2), 6 mM DTT, 4 mM MgOAc, 2 mM ATP, 0.05 mM CDP, and 100 mM KCl. After incubation at 37°C for 30 minutes and dephosphorylation, the samples were analyzed by high-performance liquid chromatography and liquid scintillation counting. The enzyme activity of RR was presented as follows: RR activity = dCDP/(CDP + dCDP) × 100%.
Cell viability assay
Cell viability was detected using the Cell Counting Kit-8 assay (CCK-8, Dojindo Laboratories, Kumamoto, Japan). Briefly, SiHa and CaSki cells were dispensed in triplicate into 96-well plates (5 × 103 cells/well) and incubated overnight at 37°C. The cells were then treated with varied concentrations of gemcitabine or/and carboplatin. After 72 hours, 10 μL of CCK-8 kit solution was added to the cells, which were then incubated for 2.5 hours at 37°C. Absorbance was then measured by a microplate reader at 450 nm (model ELX800, Bio-Tek, Winooski, VT, USA). Data were obtained from at least three separate experiments done in triplicate.
EdU incorporation assay
The inhibitory effect of gemcitabine on DNA synthesis was assessed using the Cell-Light EdU (5-ethynyl-2′-deoxyuridine) DNA Cell Proliferation Kit (RiboBio Co, Guangzhou, People’s Republic of China). SiHa cells (2.5 × 104 cells/well) were seeded in 96-well plates in triplicate and exposed to gemcitabine for 12 hours, and then treated with 50 μmol/L of EdU for 2 hours at 37°C. After being fixed with 4% paraformaldehyde for 30 minutes, the cells were treated with 0.5% Triton X-100 for 20 minutes and washed with phosphate-buffered saline three times. The cells were then exposed to 100 μL of 1 × Apollo® reaction cocktail for 30 minutes and incubated with 1 × Hoechst 33342 to stain the cell nuclei for 30 minutes. Images of the cells were captured with a fluorescence microscope (Nikon, Tokyo, Japan). ImageJ software (National Institutes of Health, Bethesda, MD, USA) was used to count the fluorescent points.
γ-H2AX detection
For immunofluorescence analysis, SiHa cells were grown on glass coverslips in six-well plates. After treatment, the cells were washed three times with cold phosphate-buffered saline and fixed in 4% paraformaldehyde for 15 minutes, and then blocked with 5% bovine serum albumin for one hour at room temperature followed by incubation with primary antibodies against γ-H2AX mouse (Upstate/Millipore) at 4°C overnight. After rinsing in phosphate-buffered saline, the cells were incubated with fluorescein isothiocyanate (FITC)-conjugated secondary antibodies for one hour at room temperature. The cells were then stained with 4′,6-diamidino-2-phenylindole (DAPI) and viewed using a fluorescence microscope (AX70, Olympus, Tokyo, Japan).
For Western blotting analysis, total proteins were extracted from the cell lines. Anti-phospho-H2AX (Ser 139) antibody was purchased from Upstate. GAPDH antibody was used as a loading control. Secondary antibodies were purchased from Li-Cor Biosciences, and blots were visualized and quantified using a Li-Cor Odyssey® Infrared Imaging scanner.
Flow cytometry measurements
SiHa cells were seeded in 35 mm dishes at a density of 3 × 105 cells, incubated for 24 hours at 37°C, and treated with gemcitabine or/and carboplatin at the indicated concentrations for 72 hours. They were then washed with cold phosphate-buffered saline twice, followed by staining with Annexin V-FITC/propidium iodide (KeyGEN, Nanjing, People’s Republic of China). Apoptotic cells were analyzed immediately on a FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA, USA) using the CellQuest 3.0 software system.
Statistical analysis
To determine the nature of the interaction between gemcitabine and carboplatin, the data from the cell viability assays were analyzed based on the Chou-Talalay method using CalcuSyn version 2.0 software (Biosoft, Cambridge, UK).Citation19,Citation20 Interaction between the drugs was quantified by determining a combination index (CI). Using this method, CI <1 indicates synergy, CI =1 indicates an additive effect, and CI > 1 indicates antagonism. The EdU incorporation assay, γ-H2AX detection, and cell apoptosis assay were performed in three separate experiments and the data are expressed as the mean ± standard deviation. The Student’s t-test was used for the statistical analysis and P<0.05 was considered to be statistically significant. The statistical analyses were performed using Statistical Package for the Social Sciences version 13 software (SPSS Inc., Chicago, IL, USA).
Results
RR subunit expression and enzyme activity were upregulated in human cervical cancer tissues
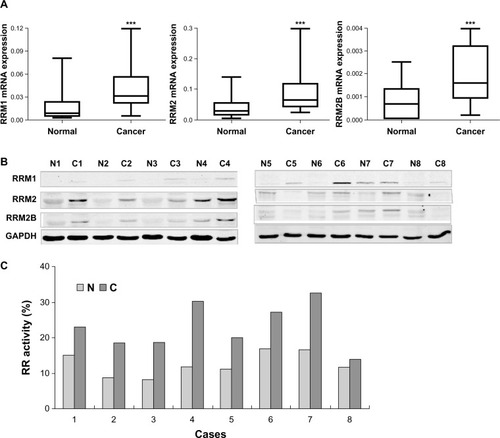
In order to investigate the roles of RR in cervical cancer, we examined the mRNA levels of the three RR subunits in the paired cancer and adjacent normal tissues from 45 cases of cervical cancer by quantitative RT-PCR. As shown in , the mRNA levels of RRM1, RRM2, and RRM2B were all upregulated in the cancer tissues compared with normal tissues (P<0.0001). In addition, we also randomly measured the subunit protein levels and enzyme activity of RR in clinical tissues from eight cases. The results showed that both the activity and subunit protein levels of RR were consistently increased in these cancer tissues when compared with normal tissues ( and ).
Figure 1 Expression and enzyme activity of three subunits of RR were upregulated in clinical cervical cancer tissues. (A) Levels of RRM1/β-actin, RRM2/β-actin, and RRM2B/β-actin in paired cervical cancer and adjacent normal tissues from 45 cervical cancer patients. Nonparametric paired (Wilcoxon signed-rank) testing indicated significantly higher levels of RRM1, RRM2, and RRM2B mRNA expression in cancer tissues compared with normal tissues. ***P<0.0001. (B) Western blot results for RRM1, RRM2, and RRM2B protein levels in the paired cancer (C) and adjacent normal (N) tissues from eight representative cervical cancer patients. (C) RR enzyme activity measured in paired cancer and adjacent normal tissues from eight representative cervical cancer patients.
Gemcitabine decreased cell viability and inhibited DNA synthesis in cervical cancer cells
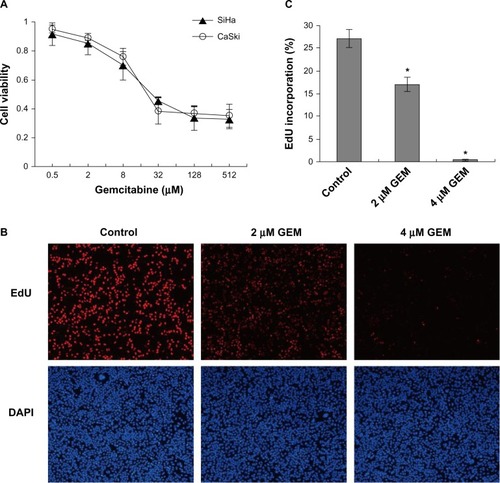
Using CCK-8 viability assays, we determined the growth inhibitory effect of gemcitabine on SiHa and CaSki cervical cancer cells in which RR are overexpressed. Cell viability was suppressed by gemcitabine in a dose-dependent manner (), and the calculated IC50 values for gemcitabine in SiHa and CaSki cells were 29.6 μmol/L and 23.8 μmol/L, respectively. EdU incorporation assays were performed to detect whether gemcitabine could affect DNA synthesis in cervical cancer cells. In and , the number of EdU-incorporated cells were decreased by treatment with gemcitabine when compared with the control. These results demonstrate that gemcitabine inhibited DNA synthesis and reduced proliferation of the cervical cancer cells.
Figure 2 Gemcitabine decreased cell viability and inhibited DNA synthesis in cervical cancer cells. (A) Cells were treated with the indicated concentrations of gemcitabine for 72 hours, and cell viability was measured with Cell Counting Kit-8 viability assays. (B) SiHa cells were exposed to gemcitabine (0, 2, and 4 μmol/l) for 12 hours. The percentage of EdU (5-ethynyl-2′-deoxyuridine) incorporation was calculated as the number of EdU-positive nuclei (red) divided by the total nuclei (blue), the total nuclei were counterstained with DAPI. (C) We selected ten visual fields under the fluorescent microscope and calculated more than 10,000 cells for each well. Counting was carried out by imageJ software. *P<0.05 for the treatment compared with the control.
Carboplatin reduced cell viability and induced DNA damage in cervical cancer cells
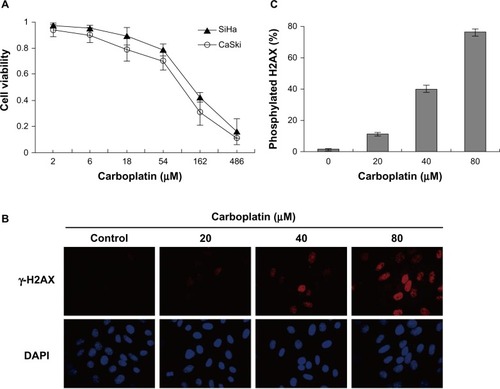
We tested the ability of carboplatin to suppress the growth of cervical cancer cells. The cell viability assays showed that carboplatin significantly inhibited growth of SiHa and CaSki cells (). The IC50 values for carboplatin were 142.4 μmol/L and 103 μmol/L for the two cell lines, respectively. In addition, to validate whether the cytotoxicity of carboplatin was associated with DNA damage, we examined phosphorylated H2AX (Ser-139, γ-H2AX) expression in SiHa cells by immunofluorescence assay. γ-H2AX has many functions and is best known for its role in DNA double-strand break repair. The results confirm that H2AX was phosphorylated after exposure to carboplatin in a dose-dependent manner, and suggest that carboplatin induced DNA damage in cervical cancer cells ( and ).
Figure 3 Carboplatin reduced cell viability and induced DNA damage in cervical cancer cells. (A) SiHa and CaSki cells were treated with the indicated concentrations of carboplatin for 72 hours, and cell viability was measured by Cell Counting Kit-8 viability assay. (B) Carboplatin induced DNA damage in SiHa cells. SiHa cells were exposed to 20, 40, and 80 μmol/L carboplatin for 12 hours, respectively. Immunofluorescence analysis was used to detect nuclear γ-H2AX foci formation with anti-H2AX antibody (red, fluorescein isothiocyanate). Nuclei were counterstained with DAPI (blue). (C) γ-H2AX-positive cells were counted under a fluorescent microscope. We calculated more than 1,000 cells for each well. Quantitative data are represented the mean ± standard deviation of three individual experiments.
Synergistic inhibitory effect of gemcitabine and carboplatin in cervical cancer cell lines
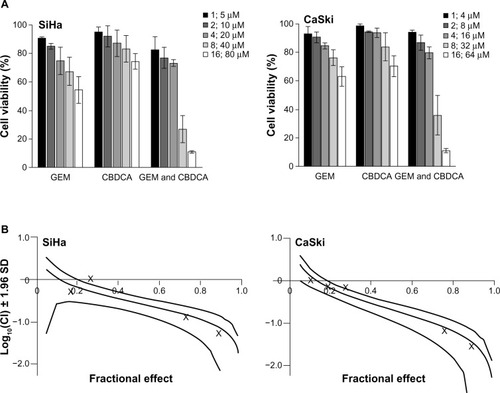
In order to assess whether gemcitabine and carboplatin have a synergistic effect, the SiHa and CaSki cervical cancer cells were treated with serial dilutions of the two drugs either alone or in combination for 72 hours (). The concentrations of gemcitabine and carboplatin maintained a constant equipotent ratio, ie, a 1:5 ratio for SiHa cells and a 1:4 ratio for CaSki cells, according to their IC50 values for the two cell lines. Gemcitabine and carboplatin were exposed at the same time in the combination group. The results show a dose response by the two cervical cancer cell lines to the treatments of gemcitabine and carboplatin either alone or in combination. The data were quantitatively analyzed using a CI based on the Chou-Talalay method. As shown in , combined treatment with gemcitabine and carboplatin yielded significantly greater growth inhibition than either agent used alone, ie, showed synergistic cytotoxicity in both SiHa and CaSki cells (log10[CI] <0).
Figure 4 Synergistic cytotoxicity of gemcitabine combined with carboplatin in cervical cancer cell lines. (A) SiHa and CaSki cells were seeded into 96-well plates and treated with gemcitabine or/and carboplatin at the indicated concentrations. After 72 hours, cell viability was measured using the Cell Counting Kit-8 viability assay. The data shown represent the mean ± standard deviation (n=3). (B) The synergistic effect of gemcitabine combined with carboplatin was quantitatively analyzed with a Cl and expressed as log10(CI) versus fractional effect Where calculable, 95% confidence intervals are shown.
Gemcitabine synergized the cytotoxicity of carboplatin in cervical cancer cells by enhancing DNA damage and cell apoptosis
To investigate the mechanism of the synergistic effect observed with the gemcitabine and carboplatin combination, we detected γ-H2AX expression in SiHa cells by immunofluorescence assay. The results show that the number of γ-H2AX-positive cells increased after carboplatin exposure, which was significantly promoted by the combined treatment of gemcitabine ( and ). Western blotting analysis of γ-H2AX expression also demonstrated that gemcitabine plus carboplatin significantly increased γ-H2AX protein expression in SiHa cells. All these results suggest that combined gemcitabine and carboplatin treatment resulted in increasing DNA damage in SiHa cells ().
Figure 5 Gemcitabine enhanced carboplatin-induced DNA damage and apoptosis in cervical cancer cells. (A) SiHa cells were treated with gemcitabine or/and carboplatin for 6 hours. Immunofluorescence analysis was used to detect γ-H2AX expression. (B) The quantitative data from and γ-H2AX-positive cells were calculated as (*P<0.05). (C) SiHa cells were treated with the indicated drugs for 6 hours. γ-H2AX protein expression was analyzed by Western blotting. (D) SiHa cells were treated with the drugs indicated for 72 hours. Apoptosis was analyzed by Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) double staining assay. (E) The degree of apoptotic cell death was quantified for each condition. Data represented the mean ± standard deviation of three individual experiments (*P<0.05).
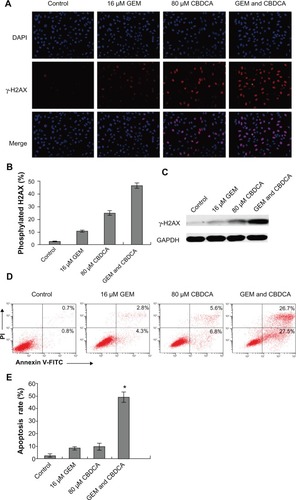
The combined effect of gemcitabine plus carboplatin was further evaluated by Annexin V/FITC staining followed by flow cytometry analysis. As shown in and , the percentage of Annexin V-positive cells was synergistically increased by the combination of gemcitabine and carboplatin (P<0.05). The data indicate that gemcitabine and carboplatin enhance cell apoptosis in cervical cancer cells. Taken together, gemcitabine and carboplatin exhibit synergistic cytotoxicity in cervical cancer cells by increasing DNA damage and cell apoptosis.
Discussion
The uncontrolled proliferation and growth of cancer cells requires extra dNTPs for DNA synthesis. Therefore, an increase in RR activity is necessary because it helps to provide excess dNTPs for DNA replication in cancer cells. Clinical studies based on quantitative RT-PCR, immunohistochemistry, and Western blotting have demonstrated that RR subunits overexpression were involved in many types of tumors.Citation21–Citation24 Kunos et al investigated the immunohistochemical expression of three RR subunits in 18 paired benign and cancerous tissues of the uterine cervix. They found that cervical cancer tissues uniformly overexpress RRM2, and that cancer relapse rates are high and disease-free intervals are shorter when RRM1 and RRM2B are simultaneously overexpressed.Citation25 Here, we not only showed that the expression levels of the three RR subunits were increased, but also, for the first time, suggest that the enzyme activity of RR is substantially upregulated in cancerous tissues in comparison with adjacent normal tissues in patients with cervical cancer (). The results suggest that upregulation of RR is probably involved in the development of cervical cancer.
It has been reported that overexpression of RRM1 is negatively related to tumorigenesis and metastasis in human and mouse lung cancer cell lines by inducing PTEN expression and reducing focal adhesion kinase phosphorylation.Citation26 RRM2 has been shown to play a critical role in enhancing invasion and metastasis in colorectal, bladder, gastric, and other cancers.Citation22,Citation27,Citation28 However, the role of RRM2B in the development of cancer is controversial. RRM2B has been shown to suppress invasion and metastasis in colorectal cancer, and its expression is associated with a better survival prognosis in patients with colorectal cancer.Citation29 However, in squamous cell carcinoma of the esophagus, RRM2B expression was significantly correlated with depth of invasion, lymph node metastasis, stage, and poor prognosis.Citation30 Accordingly, these findings reveal that subunit expression of RR is closely associated with tumor progression, invasion, and metastasis.
Platinum compounds are generally regarded as first-line chemotherapy in recurrent or metastatic cervical cancer. Clinical evidence suggests that a platinum-based combination improves response rates and prolongs survival when compared with platinum alone.Citation31 Previously, Hernández et al reported that the combination of cisplatin and gemcitabine increased cytotoxicity in cervical cancer cells,Citation32 and clinical trials showed that the combination of gemcitabine and cisplatin was tolerated, achieved enhanced response rates, and prolonged survival in patients with cervical squamous carcinoma.Citation14,Citation15,Citation33 The National Comprehensive Cancer Network guidelines (version 2.2013) recommend first-line combination therapies including cisplatin/gemcitabine (category 2B) for recurrent or metastatic cervical cancer. However, the effect of combination of gemcitabine and carboplatin on cervical cancer cells has not been experimentally investigated before. Here, we demonstrated that gemcitabine plus carboplatin had a synergistic inhibitory effect on cervical cancer cells (log10[CI] <0, ). Used separately gemcitabine inhibited DNA synthesis and carboplatin induced DNA damage ( and ), and gemcitabine plus carboplatin enhanced the DNA-damaging effects and cell apoptosis in cervical cancer cells ().
The synergistic effect of this combined drug regime could be attributed to the different but complementary mechanisms of action of the drugs involved. Carboplatin causes cellular DNA strand breaks and activates the DNA damage-responsive ATM/ATR signaling pathways,Citation34 which induces cell cycle arrest and DNA repair or cell apoptosis depending on the extent of the DNA damage.Citation35 On the other hand, gemcitabine inhibits DNA synthesis by reducing RR activity and blocking DNA strand elongation, which simultaneously prevents repair of carboplatin-induced DNA damage. Ultimately, the combined treatment synergistically suppresses cell proliferation and leads to cell death.
Other RR inhibitors, such as triapine, are also reported to increase both radiation and chemotherapy sensitivity through a mechanism involving blocking RR activity which results in sustained DNA damage in cervical cancer cells.Citation36 Further, the synergistic cytotoxicity between cisplatin and gemcitabine has been observed in human ovarian cancer cell lines and head and neck cancer xenografts,Citation13,Citation37 and one clinical study has shown that carboplatin plus gemcitabine is an active combination for patients with metastatic breast cancer.Citation38 Thus, it is probably common for RR inhibition to enhance chemoradiation sensitivity by blocking repair of DNA damage in treated cells.
In a Phase III clinical trial, the toxicity and efficacy of cisplatin doublet combinations have been already evaluated in advanced and recurrent cervical carcinoma with a Gynecologic Oncology Group study (GOG 204).Citation39 Although the doublet of cisplatin/gemcitabine has not been shown to be superior to the control arm (cisplatin/paclitaxel) in GOG 204. However, if a patient has contraindications or is not a candidate for taxanes, carboplatin and gemcitabine could be a reasonable alternative option.
In summary, we demonstrated that subunit expression and enzyme activity of RR were increased in most cervical cancer tissues, and the RR inhibitor gemcitabine synergized the cytotoxicity of carboplatin by enhancing DNA damage and cell apoptosis in cervical cancer cells. Our study might provide evidence for the combined use of gemcitabine and carboplatin in the treatment of cervical cancer.
Acknowledgments
This work was partly supported by the Preferential Society Development Program of Ningbo Municipal Science and Technology Bureau (2009C50001), the National Natural Science Foundation of China (30873094 and 30770831), the 863 National High Technology Research and Development Program of China (2012AA020206), and the KC Wong Magna Fund at Ningbo University.
Disclosure
The authors report no conflicts of interest in this work.
References
- JemalABrayFCenterMMFerlayJWardEFormanDGlobal cancer statisticsCA Cancer J Clin201161699021296855
- PecorelliSRevised FIGO staging for carcinoma of the vulva, cervix, and endometriumInt J Gynaecol Obstet200910510310419367689
- YangLDoupleEBO’HaraJACrabtreeRAEastmanAEnhanced radiation-induced cell killing by carboplatin in cells of repair-proficient and repair-deficient cell linesRadiat Res19951442302367480650
- MarkmanMChemoradiation in the management of cervix cancer: current status and future directionsOncology20138424625023392268
- BrockAPragerWBohmeRPohlmannSThe methods and results of simultaneous radiochemotherapy with carboplatin in advanced cervical carcinomasStrahlenther Onkol1994170264268 German8197548
- LeathCA3rdStraughnJMJrChemotherapy for advanced and recurrent cervical carcinoma: results from cooperative group trialsGynecol Oncol201312925125723280089
- PerezRPCellular and molecular determinants of cisplatin resistanceEur J Cancer199834153515429893624
- ShaoJZhouBChuBYenYRibonucleotide reductase inhibitors and future drug designCurr Cancer Drug Targets2006640943116918309
- SmithBDKarpJERibonucleotide reductase: an old target with new potentialLeuk Res2003271075107612921942
- JordheimLPSevePTredanODumontetCThe ribonucleotide reductase large subunit (RRM1) as a predictive factor in patients with cancerLancet Oncol20111269370221163702
- HeinemannVXuYZChubbSInhibition of ribonucleotide reduction in CCRF-CEM cells by 2′,2′-difluorodeoxycytidineMol Pharmacol1990385675722233693
- HuangPChubbSHertelLWGrindeyGBPlunkettWAction of 2′,2′-difluorodeoxycytidine on DNA synthesisCancer Res199151611061171718594
- BraakhuisBJRuiz van HaperenVWWeltersMJPetersGJSchedule-dependent therapeutic efficacy of the combination of gemcitabine and cisplatin in head and neck cancer xenograftsEur J Cancer199531A233523408652266
- BurnettAFRomanLDGarciaAAMuderspachLIBraderKRMorrowCPA phase II study of gemcitabine and cisplatin in patients with advanced, persistent, or recurrent squamous cell carcinoma of the cervixGynecol Oncol200076636610620443
- BrewerCABlessingJANagourneyRAMcMeekinDSLeleSZweizigSLCisplatin plus gemcitabine in previously treated squamous cell carcinoma of the cervix: a phase II study of the Gynecologic Oncology GroupGynecol Oncol200610038538816271750
- SteeperJRSteuartCDA rapid assay for CDP reductase activity in mammalian cell extractsAnal Biochem1970341231305440901
- ShaoJZhouBZhuLDetermination of the potency and subunit-selectivity of ribonucleotide reductase inhibitors with a recombinant-holoenzyme-based in vitro assayBiochem Pharmacol20056962763415670581
- ShaoJZhouBZhuLIn vitro characterization of enzymatic properties and inhibition of the p53R2 subunit of human ribonucleotide reductaseCancer Res2004641614729598
- ChouTCMotzerRJTongYBoslGJComputerized quantitation of synergism and antagonism of taxol, topotecan, and cisplatin against human teratocarcinoma cell growth: a rational approach to clinical protocol designJ Natl Cancer Inst199486151715247932806
- KangMHWanZKangYHSpostoRReynoldsCPMechanism of synergy of N-(4-hydroxyphenyl)retinamide and ABT-737 in acute lymphoblastic leukemia cell lines: Mcl-1 inactivationJ Natl Cancer Inst200810058059518398104
- VilmarAGarcia-FoncillasJHuarrizMSantoni-RugiuESorensenJBRT-PCR versus immunohistochemistry for correlation and quantification of ERCC1, BRCA1, TUBB3 and RRM1 in NSCLCLung Cancer20127530631221996087
- LiuXZhouBXueLRibonucleotide reductase subunits M2 and p53R2 are potential biomarkers for metastasis of colon cancerClin Colorectal Cancer2007637438117311703
- KimDJungWKooJSThe expression of ERCC1, RRM1, and BRCA1 in breast cancer according to the immunohistochemical phenotypesJ Korean Med Sci20112635235921394302
- WangLMLuFFZhangSYYaoRYXingXMWeiZMOverexpression of catalytic subunit M2 in patients with ovarian cancerChin Med J (Engl)20121252151215622884145
- KunosCARadivoyevitchTKresakAElevated ribonucleotide reductase levels associate with suppressed radiochemotherapy response in human cervical cancersInt J Gynecol Cancer2012221463146923051959
- GautamALiZRBeplerGRRM1-induced metastasis suppression through PTEN-regulated pathwaysOncogene2003222135214212687015
- MorikawaTMaedaDKumeHHommaYFukayamaMRibonucleotide reductase M2 subunit is a novel diagnostic marker and a potential therapeutic target in bladder cancerHistopathology20105788589221166702
- MorikawaTHinoRUozakiHExpression of ribonucleotide reductase M2 subunit in gastric cancer and effects of RRM2 inhibition in vitroHum Pathol2010411742174820825972
- LiuXLaiLWangXRibonucleotide reductase small subunit M2B prognoses better survival in colorectal cancerCancer Res2011713202321321415168
- OkumuraHNatsugoeSYokomakuraNExpression of p53R2 is related to prognosis in patients with esophageal squamous cell carcinomaClin Cancer Res2006123740374516778101
- ScatchardKForrestJLFlubacherMCornesPWilliamsCChemotherapy for metastatic and recurrent cervical cancerCochrane Database Syst Rev201210CD00646923076924
- HernándezPOliveraPDueñas-GonzalezAGemcitabine activity in cervical cancer cell linesCancer Chemother Pharmacol20014848849211800030
- TermrungruanglertWTresukosolDVasuratnaASittisomwongTLertkhachonsukRSirisabyaNNeoadjuvant gemcitabine and cisplatin followed by radical surgery in (bulky) squamous cell carcinoma of cervix stage IB2Gynecol Oncol20059757658115863162
- SmithJThoLMXuNGillespieDAThe ATM-Chk2 and ATR-Chk1 pathways in DNA damage signaling and cancerAdv Cancer Res20101087311221034966
- FurgasonJMBahassi elMTargeting DNA repair mechanisms in cancerPharmacol Ther201313729830823107892
- KunosCARadivoyevitchTPinkJRibonucleotide reductase inhibition enhances chemoradiosensitivity of human cervical cancersRadiat Res201017457458120954859
- MoufarijMAPhillipsDRCullinaneCGemcitabine potentiates cisplatin cytotoxicity and inhibits repair of cisplatin-DNA damage in ovarian cancer cell linesMol Pharmacol20036386286912644587
- NasrFLChahineGYKattanJGGemcitabine plus carboplatin combination therapy as second-line treatment in patients with relapsed breast cancerClin Breast Cancer2004511712215245614
- MonkBJSillMWMcMeekinDSPhase III trial of four cisplatin-containing doublet combinations in stage IVB, recurrent, or persistent cervical carcinoma: a Gynecologic Oncology Group studyJ Clin Oncol2009274649465519720909