
Abstract
Cattle establish firm dominance relationships through ritualised fights. This study aimed at investigating behaviours involved in dominance relationships and effect of factors such as weight, age and repeated fighting experience in fighting dynamics. Subject of the study was the Valdostana breed, whose cows assess dominance relationships in traditional competitions. Tournaments consist in rounds in which cows interact in pairs to assess dominance. Only winners participate in subsequent rounds. An amount of 120 fights involving 145 cows was retained, and winners (51 cows) were considered as focal individuals. An ethogram of agonistic interactions was established, including behaviours of different agonistic intensity as physical interactions (pushes, clashes), displays (threats, vocalisations), and non agonistic approaches. A transition diagram of behaviours showed a tendency to express firstly non agonistic approaches and lastly more aggressive clashes. A mixed linear model analysis on traits like competition intensity, duration, and type of behaviours expressed showed a significant effect of age difference on behaviours. However, the most important factor was the number of rounds performed: from the first to subsequent fights agonistic intensity and physical contacts increased, and displays reduced. This may be due either to the fact that more aggressive individuals were likely to be the winners, or that in higher rounds the opponents were more similar regarding fighting ability or aggressiveness and thus more intense fights occurred. The increased aggressiveness after repeated situations of competition suggests suggests that careful attention should be paid to welfare when animals are exposed to situations of high competition, like regrouping.
Introduction
Domestic animals are in large part social species showing frequent interactions among conspecifics, to establish affiliative relationships or to gain the right over resources as feeding and lying spaces (Pusey and Packer, Citation1997; Stricklin, Citation2001; Clutton-Brock et al., Citation2006). Firm dominance relationships characterise groups of cattle (Bos taurus) and other ungulates, as bison, ibex and muskoxen, both under intensive rearing system and in free-ranging conditions (Oberosler et al., Citation1982; Willisch and Neuhaus, Citation2010; Sárová et al., Citation2013). As long as group composition is constant, only few competitions between pairs of animals occur to define social dominance, and most of them involve rank-close animals (Reinhardt et al., Citation1986; Bouissou et al., Citation2001). Interactions for dominance occur in cattle at beginning of summer grazing, when different herds are conveyed in the same pasture area, or whenever cows of an herd are regrouped in relation to milk yield or stage of lactation (Phillips, Citation2002; Bøe and Færevik, Citation2003; von Keyserlingk et al., Citation2008). Fights are ritualised and involve behaviours as threats, butts, and head to head or head to neck contacts (Bouissou et al., Citation2001). The different behaviours that competitors perform permit to evaluate their own fighting ability or resource-holding potential (Parker, Citation1974) or both of them, and therefore to decide whether to continue the fight or withdraw (Arnott and Elwood, Citation2009). Competitors are used to send and receive signals of fighting ability as body size, antler length (in species where present), vocalisations or body displays (Maynard Smith and Harper, Citation2003), that often increase in intensity in the course of the interaction (i.e., escalating fight; Parker, Citation1974). The more similar are the competitors, the longer usually is the assessment (Enquist and Leimar, Citation1983; Taylor and Elwood, Citation2003). Previous agonistic experiences (winner-loser effects; Hsu et al., Citation2006) sometimes play a role in interactions, as an experience of defeat may induce to avoid a subsequent fight, and a recent victory may make individuals more likely to attack and increase the agonistic effort (Hsu and Wolf, Citation2001). Negative consequences on physiology, production and welfare, e.g. weight loss, reduced milk yield and stress increment, have been observed in cattle (as in other species) after experience of regrouping or other situations where social status needs to be settled (Grant and Albright, Citation2001; Phillips and Rind, Citation2001; von Keyserlingk et al., Citation2008; Schirmann et al., Citation2011; Castro et al., Citation2012). A number of studies have described which behaviours cows exhibit during agonistic interactions (Bouissou et al., Citation2001; Gibbons et al., Citation2009), but no work has evaluated how behaviours evolve in the course of the encounters, and how they change when a series of fights is played. Fighting cows from Alpine breeds Heréns, Aosta Chestnut and Aosta Black Pied (Plusquellec and Bouissou, Citation2001; Sartori and Mantovani, Citation2010, Citation2013; Castro et al., Citation2011) provide good opportunities to analyse the dynamics of repeated agonistic interactions in cattle. Cows of these breeds are famous for the vigorous temperament they show during dominance fights at pasture. Farmers revive these agonistic interactions in traditional cow battles that have been taken place for decades, and involve about 3000 cows per year.
Moving from the observation of cow behaviour at tournaments involving Aosta Chestnut and Aosta Black Pied, the present work firstly aimed to describe how cow interactions take place, what behaviours are involved and the sequence of the behaviours showed in the course of a conflict. Then, the first, third and fifth rounds of fights were considered to find out possible differences in behaviours exhibited and agonistic intensity as the tournament progresses.
Materials and methods
Data have been collected by video recording at folkloristic competition Batailles de Reines, that is organised following the guidelines of the farmers’ association responsible for the battle organisation. These guidelines are formulated in respect of the Italian legislation on animal care (legislative decree 116/1992) and include the control and supervision of professional veterinarians for the whole competition. Moreover, the study has been officially approved by the Ethics Committee of the University of Padova (Protocol no. 67798) that operates within the European directive 86/609/CEE regarding the protection of animals used for experimental and other scientific purposes.
Description of the subject and data collection
The traditional Batailles de Reines is a bloodless competition existing since decades in Aosta Valley, Italian North-West Alps, and aimed at reviving the competitions for dominance occurring at pasture (Mantovani et al., Citation2007). Participants are pregnant lactating cows belonging to Aosta Chestnut and Aosta Black Pied, rustic breeds from West Alps also known with the close Aosta Red Pied population as Valdostana cattle, and possible direct descendants of primitive aurochs (Bos primi-genius; Del Bo et al., Citation2001; Forabosco and Mantovani, Citation2011). The competition attracts many tourists (about 15,000 at the annual final competition) and awards the best fighters and their offspring a considerable money prize. Renowned traditional cow tournaments are also organised in the French region Haute-Savoie and in Canton Valais, Switzerland and involve cows from Heréns breed (Plusquellec, Citation2001; Plusquellec and Bouissou, Citation2001; Castro et al., Citation2011). A detailed description of Batailles de Reines has been provided elsewhere (Sartori and Mantovani, Citation2010). Briefly, annual competitions consist in tournaments (20 eliminatory and a final battle) lasting 21 days from late March to mid-October, each involving about 200 to 300 cows. A tournament takes place in a grass arena of about 50 m in diameter where 6 small patches of soil (i.e., about 10 m in diameter) are previously prepared. Here, up to 6 pairs of cows that meet for the first time are contemporarily brought to the patches to make they settle dominance relationship. Cows perceive the patch as a resource to defend, which enhances the beginning of the dominance assessment. Cows assess dominance through a series of behaviours of different agonistic intensity and involving body, horns and vocalisations. provides an example of tournament arena with four pairs of cows. Before the tournament each cow is weighed, checked for milk production and pregnancy, and assigned to one of three weight categories (category 1, over 560 kg; category 2, 510 to 560 kg; category 3, up to 510 kg). In one day three tournaments occur, one for each weight category. A first round of matches is engaged by all the participants randomly divided in pairs within the respective weight category. Cows belonging to the lightest weight category compete as first, then followed by participants of the middle category, and finally by members of the heaviest one. Cows fight in pairs, and only the winners are allowed to go to a subsequent round of match, whereas the losers have to leave the whole daily competition. During a day about 4 to 7 rounds of matches (depending on the tournament size) are played. Cows compete only with participants of the same weight category, and within round the resolution of the matches always follows the same order (i.e., category 3 first, then category 2 and finally category 1). The knockout battle scheme makes sure that within weight category participants are halved at each round until a pair disputes a final match. At the end of a daily competition the winners from each weight category achieve the title of Queen and the right to participate to the final day of competition in the year. For the present study, data on agonistic performances among cows were collected in four consecutive days of the 2009 edition of Batailles de Reines, which took place in the grass arenas of the following locations: Gressan, Nus, Saint Christophe and Aosta. Target competitions had a similar setting, as they were performed in a limited period of time (from 27 September to 18 October), and they were substantially equal in terms of competition ground. All matches were recorded using four digital video cameras with different digital supports (miniDV, mini-DVD and SDHC), for the duration of about 6 to 7 h of recording per day. All devices were set to obtain the best quality of recording and used at the same moment to catch all contemporary matches in the arena. Additional information on animal identities, individual weight and age at the day of the tournament, herd and pedigree were obtained from the farmer organisation that manages tournaments (Amis des Batailles de Reines), the regional breeders Association of Aosta Valley (AREV) and the National Breeders Association of the Valdostana breed (ANABoRaVa).
Behavioural observations
Video recordings were analysed using AvidemuxTM and JWatcherTM (Blumstein and Daniel, Citation2007) software by one single observer. Behavioural data were collected through a focal sampling of continuous recording [i.e. each interaction was traced for its whole duration; (Altmann, Citation1974)]. An amount of 120 matches chosen randomly from the first (48 matches), third (48 matches) and fifth rounds (24 matches) were retained for analysis to obtain a representative dataset of the whole tournament. Rounds after the fifth were discarded as they occurred quite rarely. Some descriptive statistics on data are reported in . The winner of each fight was chosen as focal individual (n=51 individuals), whereas behavioural information from the loser was discarded. This was done to avoid non-independent information about an interaction, and to retain at least two matches per cow: since losers must leave the whole competition when defeated, only one individual observation per match was available for them, whereas once a winner of the first or third level was observed, this cow was followed also in the subsequent fights, thus obtaining repeated observations.
An ethogram was established based on information available from the literature (e.g., Collis, Citation1976; Bouissou et al., Citation2001; Gibbons et al., Citation2009) and including all the behaviours observed in the course of the competitions and video-recorded. Observed behaviours were then grouped into six main behaviours (). The duration of each behaviour was measured considering the gap, in seconds, between the beginning of that behaviour and the start of the subsequent. Repeated behaviours in the course of a fight were summed up to acquire the individual total time spent performing that action. A score of agonistic intensity using a 1 to 6 scale was assigned to each main behaviour moving from positive/affiliative interactions to very agonistic patterns (). Main behaviours with the respective intensity score, in parenthesis, were: affiliative/non-agonistic (1) (the focal individual gently touches the other with the head, and the receiver does the same; alternatively, animals mutually lick their muzzles, or interact through sniffing and glances without any recognisable affiliative or agonistic intent); passive (1) (an animal stands in the arena in a relaxed posture and being careless of the opponent); defence of resource (2) (a participant lasts in rubbing the muzzle in the soil and scraping the ground, indicating a right over the resource); vocalisation (2) (an animal raises the nose and bellows); visual display (3) [a participant stands in front of the opponent and assumes a threatening posture by turning the head and showing the neck to opponent, or by standing up and showing the whole body. No contact occurs between individuals, and they may exhibit at the same moment, assuming a parallel (heads in the same direction) or an anti-parallel (head-back) orientation]; looking in eyes (4) (the two opponents stand very close and opposite one another, with the heads leaning towards the rival and the eyes fixed on those of the opponent. Muscles are stiff and ready to be released and used in a physical fight); pushing (5) (focal individual and opponent push each other with their heads, leveraging on the ground with their legs. As a variation, an animal push the body of the opponent with head, neck or the own body, and the opponent stands, attempting to contrast the attacker); vigorous clash (6) (forceful conflict in which an animal moves inside the arena and uses the head, the horns or the whole body to overpower the opponent, that attempts to do the same. Sometimes individuals are capable of lifting up the rival with their head).
A match starts when both contenders are brought to a patch of ground, and ends when one of them retreats and leaves the patch. Sometimes a cow decides to retreat even before meeting the opponent or just at the beginning of the mutual assessment. However, more commonly the match ends once a cow recognises that the opponent has a greatest fighting ability, and that may occur both after a non-physical assessment (behaviours of intensity 3 and 4) or during a physical struggle (behaviours of intensity 5 and 6). When the competitor leaves, the winner typically stands straight in a posture of dominance ().
Analysis of data
A total of 120 matches, performed by 145 cows and belonging to the first (n=48 matches), third (n=48) and fifth (n=24) rounds of matches, were considered. Data of only the winners (n=51 individuals) were analysed, as mentioned above. Due to technical problems of recording sometimes occurring at the beginning of tournaments, the first match of cows in some cases was missed, and for some individuals it was only possible to retain just the third and the fifth matches. A transition diagram of behaviours occurred during an interaction was established, and then a linear analysis on factors that may affect the dynamics of conflicts was carried out.
The diagram of behaviours was built using information from the individual ethograms of the selected winners. The sequential analysis tool of JWatcherTM (Blumstein and Daniel, Citation2007) permitting to produce a transitional probability matrix of behaviours was used (data not shown). Values in the matrix were obtained dividing the observed frequency for an event pair (e.g., vocalisation and looking in eyes) by the number of times the first behaviour occurred within one of the possible event pairs (Blumstein and Daniel, Citation2007).
Then, five variables were defined to describe each match (): i) the total duration of the match (DUR), in seconds; ii) the overall intensity of the match (INT), a score computed as , where d is the duration of the i-th behaviour observed during the match, and s is the intensity score assigned to the behaviour; iii) the ratio between the amount of the non-agonistic interactions and the whole duration of the match (NA); iv) the ratio between the displays and the whole duration of the match (DPY); v) the ratio between the duration of the physical contacts throughout the match and the whole duration of the match (PHY). All the ratios were expressed as frequencies. The NA included behaviours with an agonistic intensity of 1, i.e., Affiliative/non-agonistic and Passive, the DPY accounted for Defence of resource, Vocalisation, Visual display, and Looking in eyes (agonistic intensity 2 to 4); and PHY considered Pushing and Clash (agonistic intensity 5 and 6). All the five traits were analysed through single-trait mixed model for repeated measurements (MIXED procedure; SAS, Citation2009), and a Student-t test was performed to check the normality of residuals distribution (UNIVARIATE procedure; SAS, Citation2009). A series of preliminary analyses (GLM procedure; SAS, Citation2009) was performed to find out the fixed effects to include within the model. Among them, the character score, routinely scored by breeders to measure the level of aggressiveness/docility of a cow, was considered as factor to correct for a possible influence of individual aggressiveness. Since no significant differences in cow characters were detected, the effect was discarded. Other effects accounted in preliminary analyses were the breed (Aosta Chestnut or Aosta Black Pied), the absolute weight and age of the focal cow, and the number of times both contenders participated in Batailles de Reines. Homogeneity of variance was tested for each fixed effect through single-trait one-way ANOVAs (GLM Procedure; SAS, Citation2009). Model fitting statistics [i.e., akaike information criterion (AIC) coefficient; Akaike, Citation1974)] and variance components estimates retain just the factors included in the following linear model:
where yijkl is the individual observation on the k-th cow in l-th match (2 or 3 observations/cow), µ is the intercept, T is the i-th day of tournament (i=four levels), C the j-th weight category (j=three levels), T*C is the interaction between T and C, ak:ijl is the random effect of the individual within T*C, ~N (0, σ2a), b1 and b2 are the regression coefficients of Wk and Ak, respectively the individual covariates of the difference in weight (kg) and in age (years) of the target k-th individual minus the opponent, M is the effect of the l-th progressive number of matches (l=3: first, third or fifth match) in which a cow participated in a given tournament and category, and eijk is the random residual term, ~N (0, σ2e). A preliminary rank correlation between Wk and Ak (CORR procedure, SPEARMAN option; SAS, Citation2009) showed a fair and not significant correlation for participants in categories 2 and 3 (i.e., 0.13 and 0.16, respectively), and a moderate correlation of 0.48 just for cows in category 1. This allowed to include both factors in the model. A preliminary analysis of the co-variance structure among repeated measurements using the AIC coefficient indicated a non-homogeneous co-variance structure among the five analysed variables. Therefore, the co-variance structures chosen were: a variance component (Littell et al., Citation1998) for DUR and INT, an autoregressive and a heterogeneous autoregressive matrices (Ware, Citation1985) for DPY and NA respectively, and a Huynh-Feldt covariance structure (Huynh and Feldt, Citation1970) for PHY. Differences among least squares means (LSMs) of the three M levels were estimated using the Bonferroni adjustment method and compared using the CONTRAST Statement of MIXED procedure (SAS, Citation2009). The linearity of the M effect was tested also using the CONTRAST Statement. The regression curves obtained from the solutions for the fixed effects of weight difference and age difference (MIXED procedure; SAS, Citation2009) were considered in order to analyse how the five behavioural traits under study varied in relation to the increase or the reduction of age and weight differences.
Results
Some descriptive statistics about participants and matches considered in the study are reported in . Focal cows were on average 6.53±1.47 years old, i.e. 0.01±2.16 years younger than their opponent. Furthermore, they weighed 601±80 kg, i.e., 9±47 kg more than their rivals, and 2.4±0.6 of their matches were included in the present study. In the course of a dyadic interaction, cows spent most of the time in defending the resource, like the patch of soil (34 to 51% of the total time; ). Visual displays in front of the opponent (19 to 26%) and vigorous clashes (14 to 22%) followed this behaviour. About 1% of the time was spent in affiliative/non-agonistic interactions between contenders, e.g., gently touches with the heads, whereas ca. 5.16% of the time cows showed a passive posture towards the opponent. The transitional diagram obtained from the individual ethograms showed a pattern typical of an escalated conflict (). As soon as they met, cows were in some cases not interested on the opponent and expressed a passive behaviour, or, in other cases, they started to interact through non-agonistic or affiliative contacts, or through vocalisations. More commonly, cows started to defend the resource represented by the patch of earth as soon as they arrived in the arena. After this behaviour, cows usually (46% of times) engaged a visual assessment of reciprocal strength by assuming threatening postures, and this behaviour culminated in a quick glance into the opponent’s eyes (18% of times). In some cases (25% of times), after defending the resource, the cows directly moved to look in eyes, avoiding the visual display. Looking in eyes may represent a possible starting signal for a physical interaction. In half of the cases (46%), after glancing the cows engaged a physical assessment where vigorous clashes alternated with head to head or body pushing (25%). In most cases (42%) the physical assessment concluded the match. Conversely, after looking in each other’s eyes, the opponents sometimes decided to persist in defending the resource (22% of times), or to carry on a visual display (18% of times). In few cases (7%), one participant recognised the opponent as stronger, and thus decided to stop and retreat.
Among the different factors considered in the mixed model ANOVA (), tournament, weight category and interaction showed little influence on the five variables related to fighting. Specifically, the day of tournament affected significantly only DUR (P<0.05; ), showing on average longer matches on first day of video recording (at Gressan; LSM of the effect: 314.81 sec, P<0.05; data not shown) and shorter on third day (at Saint Christophe; LSM of the effect: 152.42 sec, P<0.01; data not shown). Conversely, the weight category did not show any significant effect (P>0.05) on any of the measured variables. Only transient significant effects (P<0.05; PHY variable, ) were observed for the interaction between day of tournament and weight category. The weight difference between contenders exerted no significant effects on the variables, and a P value close to the statistical significance (P=0.052) was found just for DPY. Conversely, difference in age played a significant effect (P<0.05) on NA and PHY. Regression curves obtained from the solutions of the mixed model for NA and PHY on the age difference (data not shown) showed an increment for PHY (+0.022% per +1 year of age that focal cow had respect to the opponent; P<0.05), and a negative variation for NA (-0.013% per +1 year of age difference; P<0.05). A trend of significance (P<0.10) was also found for INT, which resulted greater (+0.074 of score per +1 year of age difference) for focal individuals older than opponents. No significance was found for the regression curves of age difference on the other traits and for the regression curves of the weight difference (data not shown). The progressive number of matches disputed (M) turned out to be a significant source of variation affecting all behavioural traits (P<0.05 for DPY; P<0.01 for NA and PHY; P<0.001 for INT), except the duration of the matches (). Considering the LSMs estimated for the levels of M effect (i.e., 1st, 3rd and 5th; ), an increment in the intensity from the first to the subsequent encounters was observed (LSM for INT: 2.69 points of score vs 3.32 and 3.66 in the 1st, 3rd and 5th match, respectively). This increased intensity was linked to a percentage reduction of non-agonistic (LSM for NA: 0.11 vs 0.11 vs 0.03 in the 1st, 3rd and 5th match, respectively) and display behaviours (LSM for DPY: 0.74 vs 0.58 vs 0.59 in the 1st, 3rd and 5th match, respectively). Consistently, pushing and vigorous clashes (i.e., physical behaviour) increased from the first to the subsequent matches (LSM for PHY: 0.15 vs 0.31 vs 0.37 in the 1st, 3rd, and 5th match, respectively). Significant differences (P<0.05 and P<0.01) after Bonferroni adjustment for multiple comparisons were found between the first match and both the third and fifth ones considering INT and PHY as variables, and between the first and the third matches for DPY and the third and the fifth for NA (). A significant (P<0.01) linear variation on M effect was found for all the variables excluding the duration of the match (data not shown).


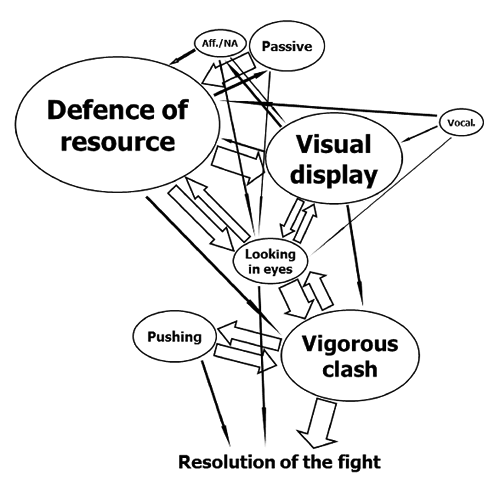
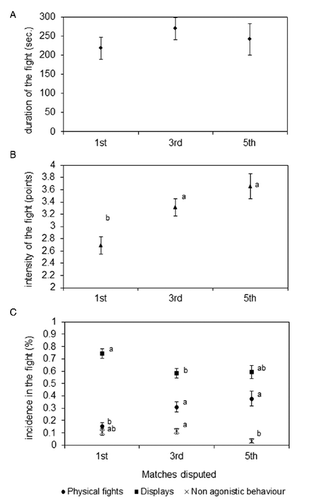
Table 1. Descriptive statistics of data of Batailles de Reines retained for analysis.
Table 2. Descriptive statistics of number of matches selected and behaviours observed.
Table 3. Results of the mixed model analyses for the fixed factors included within the model.
Discussion
The fighting behaviours we observed in Aosta Chestnut and Aosta Black Pied cattle have been previously described in related species such as bison (Bison bison; Roden et al., Citation2005) or buffalo (Syncerus caffer; Sinclair, Citation1977), and other cattle breeds [i.e., Holstein Friesian (Collis, Citation1976; Gibbons et al., Citation2009) or Scottish Highland cattle (Reinhardt et al., Citation1986)]. The ritualisation of cattle agonistic interactions and the progression of the behaviours described in the present study have been already mentioned by Bouissou and Boissy (Citation1995) and Bouissou et al. (Citation2001), in similar conditions, i.e., the traditional cow tournaments involving Heréns cows.
The agonistic interactions observed in the present study followed the dynamics of an escalated fight (Parker, Citation1974; Clutton-Brock and Albon, Citation1979) in which competitors provide each other consistent indications about their own fighting ability through honest signals with increasing costs (Zahavi, Citation1975; Enquist and Leimar, Citation1983). Other examples of escalated agonistic interactions have been found elsewhere in natural situations, e.g., in Cervids as roe deer Capreolus capreolus (Hoem et al., Citation2007), red deer Cervus elaphus (Clutton-Brock et al. Citation1979), fallow deer Dama dama (McElligott et al., Citation1998; Bartos et al., Citation2007), and in a number of other species, as Norway rat Rattus norvegicus (Lehner et al., Citation2011) or whip spider Phrynus marginemaculatus (Fowler-Finn and Hebets, Citation2006).
Regarding the effects included in the analysis, a significant effect on some variables was played by the age difference of contenders. Some influences of age in dominance disputes were also found in ungulates, i.e., roe deer (Hoem et al., Citation2007) and red deer (Jennings et al., Citation2005). Age usually plays a main role in dominance relationships of cattle (O’Connell et al., Citation1989; Murphy and Duarte, Citation1990; Phillips, Citation2002), since the social skills necessary for gaining a high rank need to be learnt (Phillips, Citation2002). Animals close in age are expected to engage agonistic interactions of greater intensity, because they generally have a closer fighting ability (Hsu et al., Citation2006). As suggested by the regression curves of age difference on PHY and INT, cows of the present study have shown an agonistic intensity greater when older than opponents, and lower when younger. This could be due to different tactics adopted by animals to face the opponent: when older than competitors, animals could win by engaging in enduring physical fights because they are generally heavier than the opponents (Arnott and Elwood, Citation2009). On the contrary, when younger, cows could win through visual assessment and quick physical fights. The use of different fighting tactics in relation to factors as contenders’ size, age or experience is broadly known among animals (Hsu et al., Citation2006; Kemp, Citation2003; Jennings et al., Citation2010), and results of present study could be explained in this perspective.
A decisive parameter for dominance in cattle as in many other species is also weight (Murphy and Duarte, Citation1990; Jennings et al., Citation2004; Arnott and Elwood, Citation2009), with heavier animals likely to gain an higher rank. However, the difference in body size did not affect interactions under study, resulting almost close to zero the regression of behaviours analysed on weight. This could be due to the organisation of competitions, dividing contenders a priori into three different weight categories, and therefore maybe reducing the effect of the weight with respect to other sources of variation. The effect of weight may also be confounded with the influence of age, but preliminary correlations on weight and age differences indicated that these two factors have not overlapped. Additionally, even the occurrence of pregnancy could be supposed to influence matches through some hormonal effects, but due to the seasonality in inseminations and calving typical of the target breeds, all cows were about at the same stage of gestation.
The main outcome of the present study is the evidence that agonistic interactions significantly changed and increased with the number of fights experienced (i.e., M effect), which is the main factor included in the analysis. Perhaps, as cows experienced a series of competitions, possible variations in hormone levels induced an increase in agonistic effort (Wingfield, Citation2005). An increased motivation to fight could have come from the experience of winning previous matches (possible winner effect; Hsu et al., Citation2009). However, this effect cannot be disentangled from the effect of a intrinsic selection for fighting ability. Indeed, the series of knock-out rounds performed could have selected more capable or aggressive opponents, and therefore an increase in agonistic effort or a longer assessment could have occurred (Enquist and Leimar, Citation1983). Some temporal variations in fighting behaviours have been observed in fishes, when new colonies are established (Oliveira and Almada, Citation1998), and when individuals ascend in ranks during hierarchic assessments (Wong and Balshine, Citation2010), but information in the literature is scarce. As for ungulates, some variations in the type of agonistic interaction performed have been found. For instance, male fallow deer (Dama dama; Mattiangeli et al., Citation1999) usually prefer lower-intensity fights called sparring in young age (Barrette and Vandal, Citation1990), and more forceful fights when older. In cattle, more frequent and longer agonistic displacements are observed in Heréns cows when they are re-introduced within their herd after longer periods of separation for calving (Castro et al., Citation2012). The interval occurring between two chances to engage agonistic interactions may affect the agonistic intensity of contenders, as observed in Heréns cows; indeed, during their regular outdoor winter exercise, cows perform a greater number of agonistic behaviours when longer periods between subsequent days of exercise occur, due to the necessity to test and re-establish the former relationships (Castro et al., Citation2011). Another situation of increased agonistic behaviour in cattle was found in pre-partum and lactating dairy cows in the first three days after regrouping (von Keyserlingk et al., Citation2008, Schirmann et al., Citation2011). In general, cows experience situations of repeated agonistic interactions at regrouping (Hasegawa et al., Citation1997) and under extremely large group sizes, because animals have not the possibility to maintain an adequate individual distance, and may have difficulties in memorising the social status of all the others (Kondo et al., Citation1989; Fregonesi and Leaver, Citation2002).
Results of the present study are referred to cows engaging interactions in traditional tournaments, but the evidence they show may be of interest in normal cow management in herds too. As amply found in cattle literature (Wierenga, Citation1990; DeVries et al., Citation2004; Huzzey et al., Citation2006), more aggressive interactions may occur when cows are exposed to prolonged competitions, both temporary (e.g., regrouping) and permanent (e.g., high density husbandry). Such situations may increase the social stress and the occurrence of physiological responses to chronic stress (Bouissou et al., Citation2001; Bøe and Færevik, Citation2003).
Conclusions
Results of this study have permitted to delineate the dynamics of agonistic interactions (e.g., displays followed by physical contact) in cow traditional tournaments, reviving dominance interactions occurring at pasture and showing how the agonistic dynamics in cows vary within a single competition and when repeated agonistic experiences occur. Differences in body weight of contenders did not play a significant role in agonistic interactions, maybe due to an aprioristic division of participants in weight category. Conversely, a greater proportion of physical interactions was found when the winner was older than the competitor, maybe due to different strategies adopted in relation to the different ages. The main factor that influenced the behaviours expressed in agonistic interactions was the occurrence of repeated agonistic experiences. With the number of fights performed, the agonistic intensity of interactions as well as the proportion of physical contacts between contenders significantly increased in comparison to the other behavioural patterns analysed.
Acknowledgments
The authors are grateful to the Regional Breeders Association of Valle d’Aosta (AREV, Aosta, Italy), the National Breeders Association of Valdostana breed (ANABoRaVa, Gressan-Aosta, Italy), and the Regional Association organising the battles (Amis de Reines, Aosta, Italy) for providing data and their technical support to the study. Authors are grateful to postgraduates and PhD students who gave a precious contribution in terms of data collection and analysis: Alberto Boldrin, Elisa Boldrin, Alex Coletti, Nina Fehlbaum, Linda Giosmin, Meriam Mrad, Raffaele Petronilli, Massimiliano Pisapia, Joy Reding, Giovanni Svigelj, Laura Treu, Giuseppe Valvo. A particular thank goes to Dr. Massimiliano Pisapia for his key role in organising data collection and to Prof. Marina Canapero for her careful linguistic revision. Special thanks to the anonymous reviewers for their help and suggestions to improve the quality of the manuscript.
References
- Akaike H., 1974. A new look at the statistical model identification. IEEE T. Automat. Contr. 19:716-723.
- Altmann J., 1974. Observational study of behaviour: sampling methods. Behaviour 49:227-267.
- Arnott G. Elwood R.W., 2009. Assessment of fighting ability in animal contests. Anim. Behav. 77:991-1004.
- Barrette C. Vandal D., 1990. Sparring, relative antler size, and assessment in male caribou. Behav. Ecol. Sociobiol. 26:383-387.
- Bartos L. Fricová B. Bartosová-VíchováJ. Panamá J. Sustr P. Smídová E., 2007. Estimation of the probability of fighting in fallow deer (Dama dama) during the rut. Aggressive Behav. 33:7-13.
- Blumstein D.T. Daniel J.C., 2007. Quantifying behaviour the J Watcher way. Sinauer Associates, Sunderland, MA, USA.
- Bøe K.E. Færevik G., 2003. Grouping and social preference in calves heifers and cows. Appl. Anim. Behav. Sci. 80:175-190.
- Bouissou M.F. Boissy A., 1995. Le comportement social des bovins et ses conséquences en élevage. INRA Prod. Anim. 18:87-99.
- Bouissou M.F. Boissy A. Le NeindreP. Veissier I., 2001. The social behaviour of cattle. In: KeelingL.J. GonyouH.W. ( eds.) Social behaviour in farm animals. CAB Int. Publ., Wallingford, UK, pp 113-145.
- Castro I.M.L. Gygax L. Wechsler B. Hauser R., 2011. Increasing the interval between winter outdoor exercise aggravates agonistic interactions in Hérens cows kept in tie-stalls. Appl. Anim. Behav. Sci. 129:59-66.
- Castro I.M.L. Gygax L. Wechsler B. Hauser R., 2012. Effect of short and long periods of separation on agonistic behaviour injuries and stress in Hérens cows kept in loose housing. Appl. Anim. Behav. Sci. 136:96-103.
- Clutton-Brock T.H. Albon S.D., 1979. The roaring of red deer and the evolution of honest advertisement. Behaviour 69:146-170.
- Clutton-Brock T.H. Albon S.D. Gibson R.M. Guinness F.E., 1979. The logical stag: Adaptive aspects of fighting in red deer (Cervus elaphus L.). Anim. Behav. 27:211-225.
- Clutton-Brock T.H. Hodge S.J. Spong G. Russell A.F. Jordan N.R. Bennett N.C. Sharpe L.L. Manser M.B., 2006. Intrasexual competition and sexual selection in cooperative mammals. Nature 444:1065-1069.
- Collis K.A., 1976. An investigation of factors related to the dominance order of a herd of dairy cows of similar age and breed. Appl. Anim. Ethol. 2:167-173.
- Del Bo L. Polli M. Longeri M. Ceriotti G. Looft C. Barre-Dirie A. Dolf G. Zanotti M., 2001. Genetic diversity among some cattle breeds in the Alpine area. J. Anim. Breed. Genet. 118:317-325.
- DeVries T.J. Von KeyserlingkM.A.G. Weary D.M., 2004. Effect of feeding space on the inter-cow distance, aggression, and feeding behavior of free-stall housed lactating dairy cows. J. Dairy Sci. 87:1432-1438.
- Enquist M. Leimar O., 1983. Evolution of fighting behaviour: decision rules and assessment of relative strength. J. Theor. Biol. 102:387-410.
- Fregonesi J.A. Leaver J.D., 2002. Influence of space allowance and milk yield level on behaviour performance and health of dairy cows housed in straw yard and cubicle systems. Livest. Prod. Sci. 78:245-257.
- Forabosco F. Mantovani R., 2011. European and indigenous cattle breeds in Italy. Schiel and Denver Publ., Houston, TX, USA.
- Fowler-Finn K.D. Hebets E.A., 2006. An examination of agonistic interactions in the whip spider Phrynus marginemaculatus (Arachnida Amblypygi). J. Arachnol. 34:62-76.
- Gibbons J.M. Lawrence A.B. Haskell M.J., 2009. Consistency of aggressive feeding behaviour in dairy cows. Appl. Anim. Behav. Sci. 121:1-7.
- Grant R.J. Albright J.L., 2001. Effect of animal grouping on feeding behaviour and intake of dairy cattle. J. Dairy Sci. 84:156-163.
- Hasegawa N. Nishiwaki A. Sugawara K. Ito I., 1997. The effects of social exchange between two groups of lactating primiparous heifers on milk production dominance order behaviour and adrenocortical response. Appl. Anim. Behav. Sci. 51:15-27.
- Hoem S.A. Melis C. Linnell J.D.C. Andersen R., 2007. Fighting behaviour in territorial male roe deer Capreolus capreolus: the effects of antler size and residence. Eur. J. Wildlife Res. 53:1-8.
- Hsu Y. Earley R.L. Wolf L.L., 2006. Modulation of aggressive behaviour by fighting experience: mechanisms and contest outcomes. Biol. Rev. 81:33-74.
- Hsu Y. Lee I.H. Lu C.K., 2009. Prior contest information: mechanisms underlying winner and loser effects. Behav. Ecol. Sociobiol. 63:1247-1257.
- Hsu Y. Wolf L.L., 2001. The winner and loser effect: what fighting behaviours are influenced? Anim. Behav. 61:777-786.
- Huynh H. Feldt L.S., 1970. Conditions under which mean square ratios in repeated measurement designs have exact F-Distributions. J. Am. Stat. Assoc. 65:1582-1589.
- Huzzey J.M. DeVries T.J. Valois P. Von KeyserlingkM.A.G., 2006. Stocking density and feed barrier design affect the feeding and social behavior of dairy cattle. J. Dairy Sci. 89:126-133.
- Jennings D.J. Carlin C.M. Hayden T.J. Gammell M.P., 2010. Investment in fighting in relation to body condition, age and dominance rank in the male fallow deer, Dama dama. Anim. Behav. 79:1293-1300.
- Jennings D.J. Gammell M.P. Carlin C.M. Hayden T.J., 2004. Effect of body weight antler length resource value and experience on fight duration and intensity in fallow deer. Anim. Behav. 68:213-221.
- Jennings D.J. Gammell M.P. Payne R.J.H. Hayden T.J., 2005. An investigation of assessment games during fallow deer fights. Ethology 111:511-525.
- Kemp D.J., 2003. Twilight fighting in the evening brown butterfly, Melanitis leda (L.)(Nymphalidae): age and residency effects. Behav. Ecol. Sociobiol. 54:7-13.
- Kondo S. Sekine J. Okubo M. Asahida Y., 1989. The effect of group size and space allowance on the agonistic and spacing behaviour of cattle. Appl. Anim. Behav. Sci. 24:127-135.
- Lehner S.R. Rutte C. Taborsky M., 2011. Rats benefit from winner and loser effects. Ethology 117:949-960.
- Littell R.C. Henry P.R. Ammerman C.B., 1998. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 76:1216-1231.
- Mantovani R. Contiero B. Vevey M., 2007. Genetic evaluation of cow fighting ability in the Valdostana breed. Ital. J. Anim. Sci. 6(Suppl.1):156-158.
- Mattiangeli V. Mattiello S. Verga M., 1999. The fighting technique of male fallow deer (Dama dama): an analysis of agonistic interactions during the rut. J. Zool. 249:339-346.
- Maynard SmithJ. Harper D., 2003. Animal signals. Oxford University Press, Oxford, UK.
- McElligott A.G. Mattiangeli V. Mattiello S. Verga M. Reynolds C.A. Hayden T.J., 1998. Fighting tactics of fallow bucks (Dama dama Cervidae): reducing the risks of serious conflict. Ethology 104:789-803.
- Murphy R.M. Duarte F.A., 1990. Social aggregations in cattle: II. Contributions of familiarity and genetic similarity. Behav. Genet. 20:355-368.
- Oberosler R. Carenzi C. Verga M., 1982. Dominance hierarchies of cows on Alpine pastures as related to phenotype. Appl. Anim. Ethol. 8:67-77.
- O’Connell J. Giller P.S. Meaney W., 1989. A comparison of dairy cattle behavioural patterns at pasture and during confinement. Irish J. Agr. Food Res. 28:65-72.
- Oliveira R.F. Almada V.C., 1998. Dynamics of social interactions during group formation in males of the cichlid fish Oreochromis mossambicus. Acta Ethol. 1:57-70.
- Parker G.A., 1974. Assessment strategy and the evolution of fighting behaviour. J. Theor. Biol. 47:223-243.
- Phillips C.J.C., 2002. Cattle behaviour and welfare. Blackwell, Oxford, UK.
- Phillips C.J.C. Rind M.I., 2001. The effects on production and behaviour of mixing uniparous and multiparous cows. J. Dairy Sci. 84:2424-2429.
- Plusquellec P., 2001. Influence d’une sélection pour la combativité et l’aptitude à la dominance sur le comportement social et la réactivité émotionnelle des bovins domestiques femelles de la race d’Hérens. PhD Diss., University of Paris XIII, France.
- Plusquellec P. Bouissou M.F., 2001. Behavioural characteristics of two dairy breeds of cows selected (Hérens) or not (Brune des Alpes) for fighting and dominance ability. Appl. Anim. Behav. Sci. 72:1-21.
- Pusey A.E. Packer C., 1997. The ecology of relationships. In: KrebsJ.R. DaviesN.B. ( eds.) Behavioural ecology: an evolutionary approach. Blackwell, Oxford, UK, pp 254-283.
- Reinhardt C. Reinhardt A. Reinhardt V., 1986. Social behaviour and reproductive performance in semi-wild Scottish Highland cattle. Appl. Anim. Behav. Sci. 15:125-136.
- Roden C. Vervaecke H. Van ElsackerL., 2005. Dominance age and weight in American bison males (Bison bison) during non-rut in semi-natural conditions. Appl. Anim. Behav. Sci. 92:169-177.
- ŠárováR. Špinka M. Stěhulová I. Ceacero F. ŠimečkováM. Kotrba R., 2013. Pay respect to the elders: age, more than body mass, determines dominance in female beef cattle. Anim. Behav. 86:1315-1323.
- Sartori C. Mantovani R., 2010. Genetic of fighting ability in cattle using data from the traditional battle contest of Valdostana breed. J. Anim. Sci. 88:3206-3213.
- Sartori C. Mantovani R., 2013. Indirect genetic effects and the genetic bases of social dominance: evidence from cattle. Heredity 110:3-9.
- SAS, 2009. SAS user’s guide: statistics. Version 9.2. SAS Inst. Inc., Cary, NC, USA.
- Schirmann K. Chapinal N. Weary D.M. Heuwieser W. von KeyserlingkM.A.G., 2011. Short-term effects of regrouping on behaviour of prepartum dairy cows. J. Dairy Sci. 94:2312-2319.
- Sinclair A.R.E., 1977. The African buffalo: a study of resource limitation of populations. University of Chicago Press, Chicago, IL, USA.
- Stricklin W.R., 2001. The evolution and domestication of social behaviour. In: KeelingL.J. GonyouH. ( eds.) Social behaviour in farm animals. CAB Int. Publ., Wallingford, UK, pp 83-110.
- Taylor P.W. Elwood R.W., 2003. The mismeasure of animal contests. Anim. Behav. 65:1195-1202.
- von KeyserlingkM.A.G. Olenick D. Weary D.M., 2008. Acute behavioural effects of regrouping dairy cows. J. Dairy Sci. 91:1011-1016.
- Ware J.H., 1985. Linear models for the analysis of longitudinal studies. Am. Stat. 39:95-101.
- Wierenga H.K., 1990. Social dominance in dairy cattle and the influences of housing and management. Appl. Anim. Behav. Sci. 27:201-229.
- Willisch C.S. Neuhaus P., 2010. Social dominance and conflict reduction in rutting male Alpine ibex, Capra ibex. Behav. Ecol. 21:372-380.
- Wingfield J.C., 2005. A continuing saga: the role of testosterone in aggression. Horm. Behav. 48:253-255.
- Wong M. Balshine S., 2010. Fight for your breeding right: hierarchy re-establishment predicts aggression in a social queue. Biol. Lett. 7:190-193.
- Zahavi A., 1975. Mate selection. A selection for a handicap. J. Theor. Biol. 53:205-214.