Abstract
Aurora kinase A (Aur-A), a mitotic kinase, regulates initiation of mitosis through centrosome separation and proper assembly of bipolar spindles. LIM kinase 1 (LIMK1), a modulator of actin and microtubule dynamics, is involved in the mitotic process through inactivating phosphorylation of cofilin. Phosphorylated LIMK1 is recruited to the centrosomes during early prophase, where it colocalizes with γ-tubulin. Here, we report a novel functional cooperativity between Aur-A and LIMK1 through mutual phosphorylation. LIMK1 is recruited to the centrosomes during early prophase and then to the spindle poles, where it colocalizes with Aur-A. Aur-A physically associates with LIMK1 and activates it through phosphorylation, which is important for its centrosomal and spindle pole localization. Aur-A also acts as a substrate of LIMK1, and the function of LIMK1 is important for its specific localization and regulation of spindle morphology. Taken together, the novel molecular interaction between these two kinases and their regulatory roles on one another’s function may provide new insight on the role of Aur-A in manipulation of actin and microtubular structures during spindle formation.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Author Contributions
L.R. conducted all kinase assays, mutagenesis experiments, pull down assays and involved in writing the manuscript. R.O. performed all microscopy and knockdown experiments. M.R. performed coimmunoprecipitation experiments. R.C. conceived the idea, analyzed data and wrote the manuscript.
Acknowledgements
We are thankful to the National Cancer Institute of National Institute of Health and the Prostate Cancer Research Program of the Department of Defense for funding this research.
Figures and Tables
Figure 1 LIMK1 and Aur-A colocalized to the centrosomes in mitotic cells. (A) Upper and lower parts: immunofluorescence analysis of pLIMK-pT508 (red) and Aur-A (green) in PC-3 cells. DNA was counterstained with DAPI. Colocalization of pLIMK1 with Aur-A in early prophase was mainly to the centrosomes (white arrows). (B) Pearson's correlation of colocalization between LIMK1 and Aur-A from (A). (C) Immunofluorescence of LIMK1 (red) and Aur-A (green) during mitosis. A distinct colocalization of pLIMK1 (only pLIMK1 localizes to the centrosomes) and Aur-A at the centrosomes during prophase was evident. Strong association of Aur-A with the microtubules also could be noted. At prometaphase, continued localization of pLIMK1 at the separating centrosomes along with Aur-A could be noted (white arrows). In metaphase, redistribution of pLIMK1 at the spindle poles but not at the spindle microtubules and its sustained colocalization with Aur-A was seen (white arrows). In anaphase, pLIMK1 remained colocalized with Aur-A to the poles of the elongated spindle. All pictures are representatives of at least three independent experiments. Scale bar 5 µm.
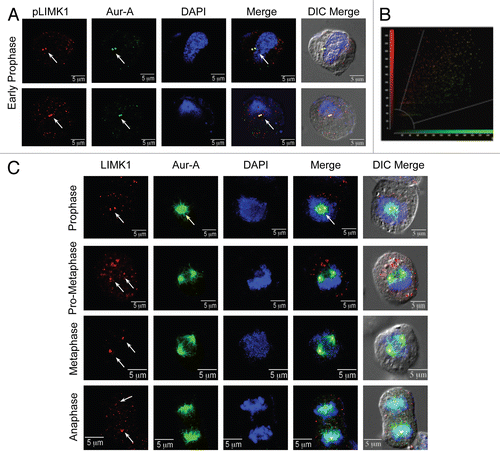
Figure 2 LIMK1 acts as a substrate of Aur-A. (A and B) Kinase assays with inactive GST-LIMK1 (500 ng) and GST-Aur-A (50 ng) kinases and His-cofilin (1 µg) or MBP (0.5 µg) as respective substrates. (A) Coomassie stained SDS-PAGE showing location of the peptide bands. MW, molecular weight marker. (B) Autoradiogram showing no phosphorylation of His-cofilin by GST-LIMK1 (lane 2), which confirms its inactivity. Strong phosphorylation of MBP and LIMK1 by Aur-A (lanes 4 and 5) and autophosphorylation of Aur-A (lane 4 and 5) could be seen. Aur-A autophosphorylation seemed to be enhanced in the presence of MBP and LIMK1. (C and D) Phosphorylation of His-LIMK1K by active His-Aur-A. (C) Coomassie stained SDS-PAGE. (D) Autoradiogram showing increased phosphorylation intensity with increasing amounts of His-LIMK1K (0.25 µg, 0.5 µg and 1 µg) by active His-Aur-A (0.22 µg) (lanes 4–6) but not by inactive His-Aur-AK162M kinase (0.22 µg) (lanes 7–9). His-Aur-AK162M was also unable to phosphorylate MBP (1 µg) (lane 2), which confirms catalytic inactivity of Aur-AK162M mutant. SDS-PAGE images are representatives of three to five independent experimental repeats.
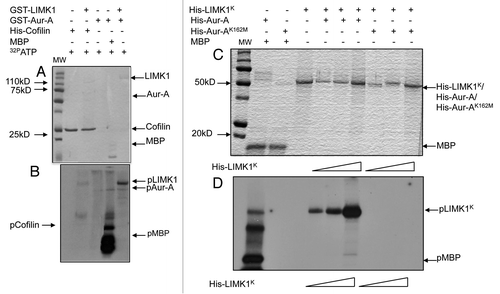
Figure 3 Aur-A physically associates with LIMK1. (A) Immunoprecipitation and reverse immunoprecipitation of FLAG-LIMK1 or Aur-A from BPHV and BPHLCA cell extracts. 500 µg total extracts were immunoprecipitated using either anti-FLAG or anti-Aur-A antibodies. Rabbit IgG was used as the negative control. Data shows strong immunoprecipitated FLAG-LIMK1 or Aur-A bands from cells expressing FLAG-LIMK1 (lanes 2 and 4). No bands could be seen in lanes for BPHV control cell extracts (lanes 3, 5, 7 and 8). No nonspecific bands were noted with rabbit antibodies (lanes 6 and 7). (B and C) Interaction between His-Aur-A and endogenous LIMK1 affinity precipitated from PC-3 extracts. (B) Immunoblots showing binding of LIMK1 with both His-Aur-A and His-Aur-AK162M (lanes 3 and 4). No nonspecific binding was noted with the beads (lane 2). (C) Densitometric analysis of the binding affinity of Aur-A and Aur-AK162M to LIMK1 from equal amounts of extracts. Data shows a 50% reduction in the affinity of binding of Aur-AK162M with LIMK1. Data represents mean ± SD of three independent experiments. (D) Interaction of Aur-A with different domains of LIMK1. Total extracts of RWPE-1 cells transiently transfected with pCMVLIMK1-FLAG, pCMVLIMK1LD-FLAG or pCMVLIMK1K-FLAG were used for affinity precipitation with His Aur-A. Data shows that in addition to wild-type LIMK1, Aur-A was capable of binding both LIM domains and kinase domain independently. The lane for bead control (lane 1) shows some nonspecific binding, but the intensity was much lower than the beads with bound Aur-A. Data shows representative images from three independent experiments.
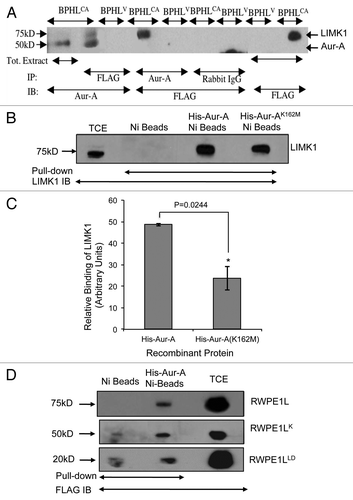
Figure 4 Aur-A phosphorylated LIMK1 at a site different than its activating phosphorylation site. (A) Phosphopeptide analysis of GST-LIMK1 after incubation with GST-Aur-A in kinase assay buffer showing phosphorylation of S307 at the linking region between the PDZ, and the kinase domain, which shows a partial motif for Aur-A phosphorylation (L at +1 position after phosphorylating residue). (B and C) Kinase assay showing loss of phosphorylation by His-Aur-A of His-LIMK1 with S307A mutation. (B) Coomassie stained SDS-PAGE. (C) Autoradiogram of the kinase assay with His-LIMK1 (0.5 µg), His-LIMK1S307A (0.5 µg), His-Aur-A (0.22 µg) and His-cofilin (1 µg). Both His-LIMK1 and His-LIMK1S307A were inactive, as no His-cofilin phosphorylated polypeptide was detected (lanes 4 and 7). While strong phosphorylation of the wild-type LIMK1 by His-Aur-A was noted (lane 4), very weak to no phosphorylation could be seen with LIMK1S307A mutant protein (lane 8), which further confirms the unique Aur-A phosphorylation site on LIMK1. (D) Phosphorylation of His-LIMK1 at S307 by Aur-A was essential for phosphorylation at T508. Immunoblot analysis of nonradioactive kinase assays using phosphospecific antibodies (pT508) showing strong phosphorylation of wild-type His-LIMK1 at T508 by His-Aur-A (lane 2), which was not noted with Aur-AK162M (lane 3). No phosphorylation at T508 could be seen when His-LIMK1S307A was incubated with His-Aur-A (lane 5). Some nonspecific signals were noted in lanes 1, 3 and 5. Data presented are representatives of at least three separate experiments.
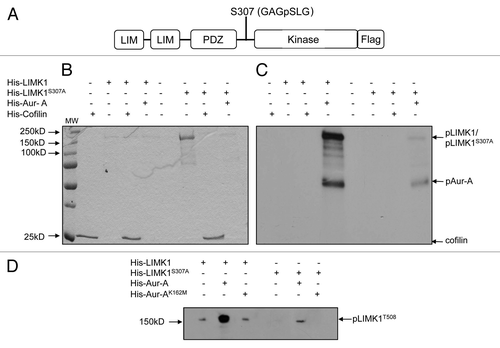
Figure 5 Aur-A allows T508 phosphorylation on endogenously expressed LIMK1. (A) Nonradioactive kinase assays using calf intestinal phosphatase (CIP) (5 units) treated extracts (50 µg) of RWPE-1 cells transfected with pCMVLIMK1-FLAG. CIP-treated extracts were incubated with His-Aur-A (0.22 µg) with phosphatase inhibitor (PPI) (lane 3), and phosphopeptide band was detected by western blotting using anti-p508-LIMK1 antibodies. Data show strong phosphorylation at T508 by His-Aur-A but not without Aur-A. No phosphorylated LIMK1 (T508) could be seen in the absence of PPI (lane 1). GAPDH was used as the loading control. (B) and (C) Nonradioactive kinase assays using immunoprecipitated FLAG-tagged LIMK1 from CIP treated (100 units) transfected RWPE-1 cell extracts (500 µg) or LIMK1 from CIP treated (100 units) PC-3 cell extracts (500 µg) and His-Aur-A (0.22 µg). Phosphorylated LIMK1 at T508 was detected by immunoblotting using anti-pT508-LIMK1 antibodies. (B) A strong phosphorylated band of FLAG-LIMK1 at T508 was evident upon incubation with His-Aur-A but not in the lane without Aur-A. (C) A similar phosphorylation at T508 of immunoprecipitated LIMK1 by His-Aur-A was noted (lane 3) which was not seen upon incubation with Aur-AK162M, which confirms the requirement of active Aur-A to achieve phosphorylation at T508 of LIMK1. Data show a representative image of at least three independent experiments.
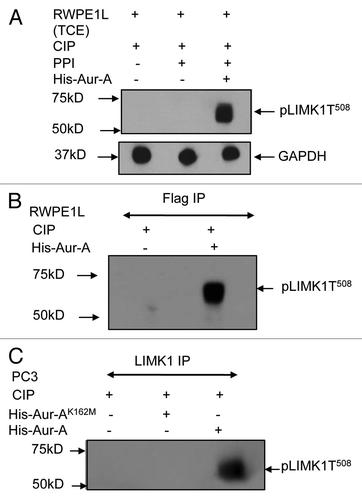
Figure 6 LIMK1 mutant did not colocalize with pAur-A. (A) Upper two parts: immunofluorescence analysis of LIMK1 (red) using anti-FLAG antibodies and pAur-A (green) in P69 cells transfected with LIMK1-FLAG constructs. Lower two parts: localization of LIMK1S307A using anti-FLAG antibodies and pAur-A in transfected P69 cells. Mitotic spindle and asters are shown by α-tubulin staining (magenta). DNA was counterstained with DAPI. Scale bar: 10 µm. (B) Relative correlation coefficient of the colocalized proteins as determined using the average value of two centrosomes per sample/cell. Results presented as mean ± SD of 10 individual cells from at least three independent experiments.
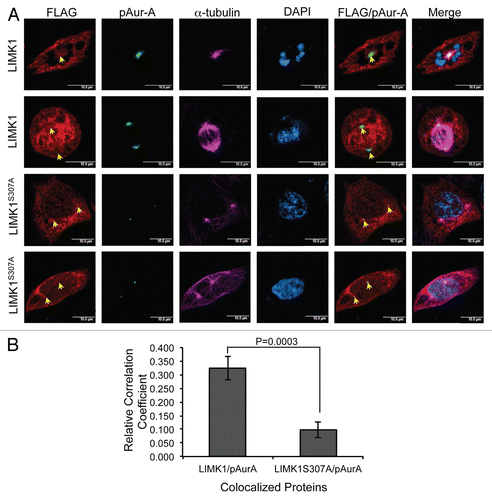
Figure 7 Aur-A inhibition led to mislocalization of pLIMK1. (A and B) Immunofluorescence analysis of nocodazole synchronized mitotic shake-off PC-3 and RWPE-1 cells following treatment with MLN8237 or vehicle. (A and B) Mitotic spindles are shown by α-tubulin staining in red, and spindle poles localization of pAur-A-pT288 is shown in green. Treatment with MLN8237 showed a distinct deformation of the spindles and altered localization of pAur-A in the mitotic cells (lower parts) compared with vehicle-treated cells. (C and D) Centrosomes are shown by staining of centrosomal protein centrin in red and pLIMK1 (T508) in green. Upper parts: in vehicle-treated cells, pLIMK1 was colocalized with centrin at the centrosomes. Inhibitor treatment (lower parts) showed a diffused staining of pLIMK1 at the periphery and mislocalization pLIMK1 from the centrosomes (orange arrow). Some diffused staining of centrin at the cell periphery was also noted in MLN8237-treated cells, although some centrosomal localization could be seen (white arrows). Scale bar: 5 µm.
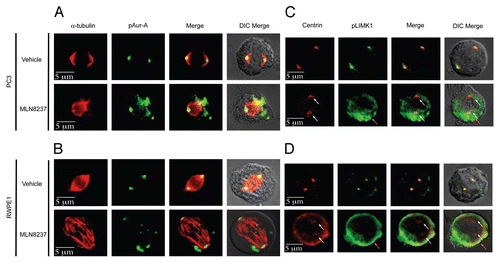
Figure 8 LIMK1 phosphorylates Aur-A. (A and B) Kinase assays using immunoprecipitated LIMK1 and His-Aur-AK162M and His-cofilin as the substrates. (A) Coomassie stained SDS-PAGE of the kinase assays. (B) Autoradiogram of the SDS-PAGE showing phosphorylation of His-Aur-AK162M (lane 3) and His-cofilin (lane 1) by LIMK1. LIMK1 was immunoprecipitated from PC-3 cells using anti-LIMK1 antibodies and incubated with His-Aur-AK162M or His-cofilin in kinase assay buffer with γ-32P-ATP. (C) Knockdown of LIMK1 leads to decreased levels of pAur-A. PC-3 cells were transiently transfected with either scrambled or LIMK1 shRNA vectors. Cells were harvested 48 hr post-transfection and levels of LIMK1, pAur-AT288 and Aur-A were detected by western blotting using anti-LIMK1, anti-pAur-AT288 or anti-Aur-A antibodies. A significant reduction in the levels of pAur-A-pT288 but not total Aur-A was evident in cells with reduced levels of LIMK1. GAPDH was used as the loading control. (D) Quantification of the relative pAur-AT288 levels from (C) by densitometry. Data show mean ± SD of three separate experiments.
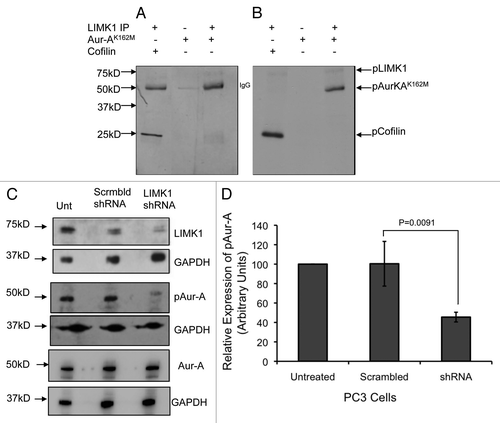
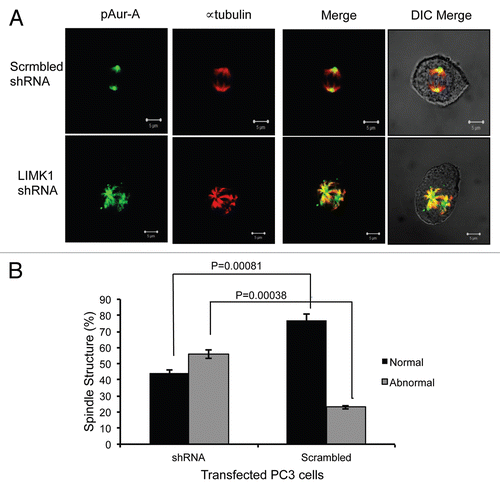
Figure 9 Knockdown of LIMK1 caused altered localization of pAur-AT288 and spindle disorganization. (A) Immunofluorescence analysis of pAur-AT288 (green) localization and α-tubulin staining (red) showing mitotic spindle structures in LIMK1 shRNA transfected PC-3 cells. A diffused staining of pAur-A in the aster microtubules with multiple speckles could be seen in cells with reduced expression of LIMK1 compared with scrambled shRNA-expressing cells. Lack of normal spindle structures was also evident in shRNA-transfected cells. (B) Quantification of spindle abnormalities of scrambled and LIMK1 shRNA transfected PC-3 cells showing a significant increase in the number of cells with abnormal spindles. A total of 200 transfected cells were counted. Data show mean ± SD of the percent change in the spindle structure of three separate experiments.