
Abstract
Fragmentation is a degradation pathway ubiquitously observed in proteins despite the remarkable stability of peptide bond; proteins differ only by how much and where cleavage occurs. The goal of this review is to summarize reports regarding the non-enzymatic fragmentation of the peptide backbone of monoclonal antibodies (mAbs). The sites in the polypeptide chain susceptible to fragmentation are determined by a multitude of factors. Insights are provided on the intimate chemical mechanisms that can make some bonds prone to cleavage due to the presence of specific side-chains. In addition to primary structure, the secondary, tertiary and quaternary structures have a significant impact in modulating the distribution of cleavage sites by altering local flexibility, accessibility to solvent or bringing in close proximity side chains that are remote in sequence. This review focuses on cleavage sites observed in the constant regions of mAbs, with special emphasis on hinge fragmentation. The mechanisms responsible for backbone cleavage are strongly dependent on pH and can be catalyzed by metals or radicals. The distribution of cleavage sites are different under acidic compared to basic conditions, with fragmentation rates exhibiting a minimum in the pH range 5 to 6; therefore, the overall fragmentation pattern observed for a mAb is a complex result of structural and solvent conditions. A critical review of the techniques used to monitor fragmentation is also presented; usually a compromise has to be made between a highly sensitive method with good fragment separation and the capability to identify the cleavage site. The effect of fragmentation on the function of a mAb must be evaluated on a case-by-case basis depending on whether cleavage sites are observed in the variable or constant regions, and on the mechanism of action of the molecule.
Acknowledgments
The authors are grateful to Dr. Steve L. Cohen for insightful discussions and comments on the manuscript.
Figures and Tables
Figure 1 pH-dependence of the major fragmentation sites in a mAb.
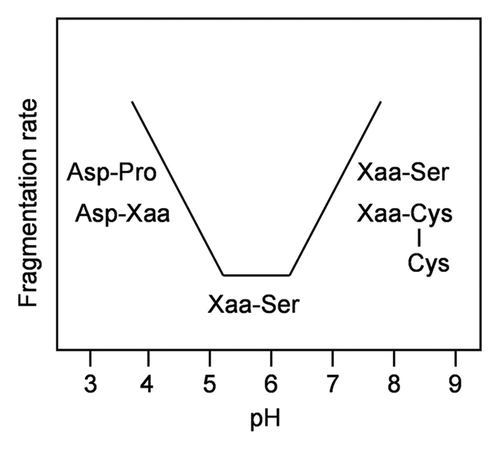
Figure 2 Reducing SDS PAGE analysis of a stressed mAb (2 weeks at 45°C, pH 9) and its SEC-purified monomer. Compared to the whole sample, the monomeric fraction lacks bands corresponding to the hinge cleavage (Fc HC and Fab HC) but contains similar bands corresponding to several cleavages within the immunoglobulin domains. Cleavage in the CH1 domain loop K133STSGGT yields two fragments of approximately 35 and 15 kDa.
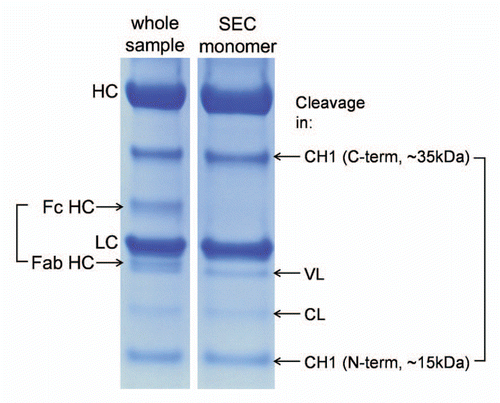
Figure 3 Frequently observed cleavage sites in mAbs. Only one heavy chain and one light chain are shown. Dotted lines-disulfide bridges, shaded boxes-cleavage sites. Glycosylated Asn297 is indicated.
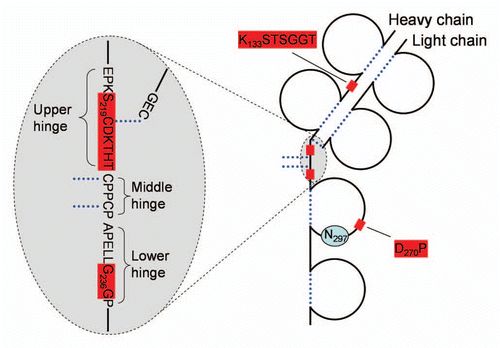
Figure 4 (A) Schematic drawing of a portion of IgG1 and IgG2 hinge region indicating sites of fragmentation. (B) pH-dependence of IgG1 hinge fragmentation monitored by SEC. Fragmentation between S219 and C220 in IgG1 that proceeds via β-elimination is shown as a solid line. The rest of the cleavages are shown as dotted lines. Disulfide bridges are indicated. The samples in (B) were incubated for approximately two weeks at 45°C. To identify the cleavage sites, the Fab fragments were purified by SEC and then analyzed by LC/MS. Considering the relatively wide pH range of 4–10, the cleavage sites can be divided into three groups. (1) Ser-Cys bond, where cleavage occurs via beta elimination, becomes dominant at higher pH. (2) Asp-Lys bond, where cleavage dramatically increases at pH < 5, presumably involving Asp side chain. (3) The rest of the cleavage sites in the hinge, where pH dependence is not as remarkable.
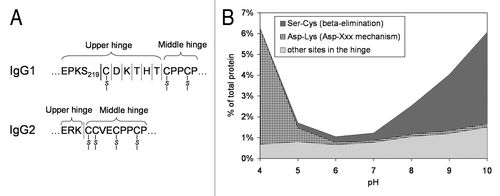
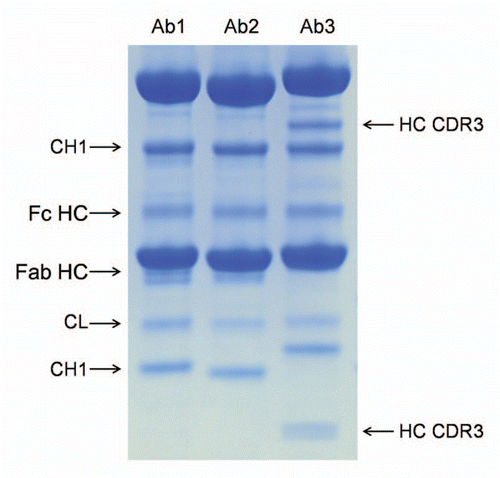
Figure 5 Comparison of the cleavage pattern in three monoclonal antibodies. The three antibodies were stored in PBS at pH 7.0 for three months at 45°C. All antibodies show comparable cleavage in the hinge (Fc HC band, the Fab HC band is not apparent in Ab3 due to co-migration with the light chain), comparable cleavage in the K133STSGGT loop (in agreement with the sequence of the variable domain the N-terminal fragment (CH1 band) showed different molecular weight), comparable cleavage in the CL domain. Ab3 has two additional bands that correspond to two cleavages (seen as a doublet on the small N-terminal fragment) in heavy chain CDR3.
Schematic 1 pH-dependence of peptide bond hydrolysis.Citation8
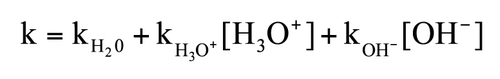
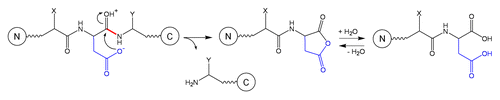
Schematic 2 Fragmentation at Asp-Xaa site.Citation3,Citation15
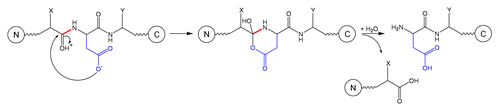
Schematic 3 Fragmentation at Xaa-Asp site.Citation3
Schematic 4 Fragmentation at Xaa-Ser site.Citation3
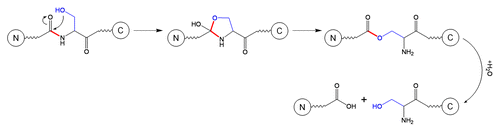
Schematic 5 β-elimination at Ser residue.Citation11
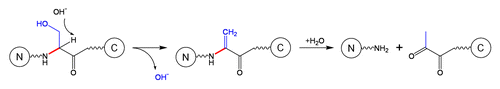
Schematic 6 Cleavage of a disulfide bond followed by fragmentation at dehydroalanine.Citation11
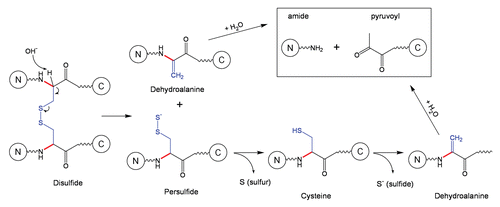
Schematic 7 Fragmentation at Asn-Xaa sites.Citation27
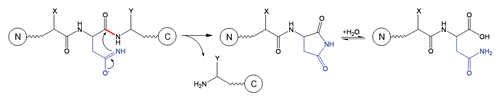
Table 1 Summary of residues involved in peptide bond cleavage, conditions for testing and references that discuss mechanisms of cleavage