
Abstract
Interleukin-21 (IL-21) is a type I four-helical bundle cytokine that exerts a variety of significant effects on many hematopoietic cells, including T and B lymphocytes and natural killer cells. IL-21 is produced predominantly by CD4+ T cells and natural killer T cells and, when aberrantly overexpressed, appears to play important roles in a wide variety of autoimmune disorders. To generate potential therapeutic reagents capable of inhibiting IL-21 for clinical use, we immunized human immunoglobulin transgenic mice with IL-21 and then identified and cloned a panel of human anti-human IL-21 binding monoclonal antibodies. IL-21 neutralizing and IL-21-binding, non-neutralizing antibodies were assigned to distinct epitope “bins” based on surface plasmon resonance competition studies. The most potent neutralizing antibodies had extremely high (sub pM) affinity for IL-21 and were able to block IL-21 activity in various biological assays using either an IL-21R-transfected pre-B-cell line or primary human B cells, and their neutralizing activity was, in some cases, superior to that of a soluble form of the high affinity heterodimeric IL-21 receptor. Characterization of this panel of IL-21 antibodies provided the basis for the selection of a therapeutic candidate antibody capable of inhibiting IL-21 activity for the treatment of autoimmune and inflammatory diseases.
Disclosure of Potential Conflicts of Interest
All authors are current or former employees and shareholders of ZymoGenetics, Inc.
Acknowledgments
We gratefully acknowledge the helpful discussions, generation of reagents, and other contributions to this project by many of our past and present ZymoGenetics colleagues, including: Ty Brender, Jennifer Brody, Chung-Leung Chan, Mike Dodds, Frank Grant, Jane Gross, Matt Holdren, Tracey (Pownder) Jurista, Cecile Krejsa, Joe Kuijper, Megan Lantry, Katherine Lewis, Hong Liu, Pat McKernan, Kathy Mink, Julie Parrish-Novak, Scott Presnell, Fred Ramsdell, Brian Reardon, P.V. Sivakumar, Marrissa Smith, Mike Stamm, Kim Waggie, Jim West, Rob West and Eugene Yi. This work was funded by ZymoGenetics, Inc.
Figures and Tables
Figure 1 Affinity and epitope bin distribution. (A) The mAbs were screened for binding affinity to rhIL-21 via surface plasmon resonance (BIAcore). Association rate constants (ka) and dissociation rate constants (kd) were plotted in an affinity isotherm plot for mAbs in Bin 1 (filled circles), Bin 2 (open squares), Bin 1/2 (asterisks), Bin 3 (filled triangles) and the soluble heterodimeric receptor (open triangle). Values for a specific subset of the mAb clones are indicated with letters. (B) Epitope bin assignment of the mAbs.
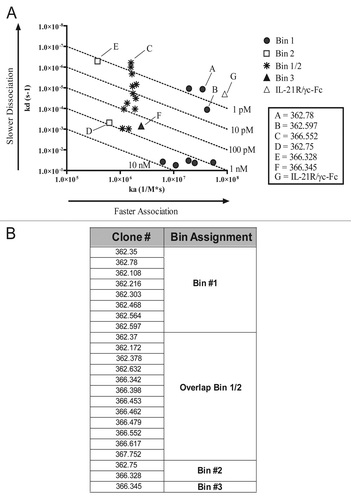
Figure 2 Neutralization of IL-21-mediated activity in vitro. (A) Neutralization of STAT3 phosphorylation by IL-21 mAbs and the soluble receptor. Thirty-two pM rhIL-21 was incubated with three CHO-expressed recombinant IL-21 neutralizing mAb clones 362.78 (filled squares), 366.328 (open triangles), 366.552 (X's), or soluble IL-21R/γc-Fc (asterisks) prior to stimulation of BaF3 cells expressing IL-21R. Phospho-STAT3 was measured following a 15 min incubation. (B) Neutralization of STAT-mediated luciferase activity. rhIL-21 (641 pM) was incubated with IL-21 mAbs or soluble IL-21R/γc for 30 min prior to culture with BaF3/IL-21R cells. After 20 h, STAT-mediated luciferase activity was measured. (C) Neutralization of IL-21-mediated B cell proliferation. Primary human B cells were incubated for 4 d with 3.21 nM (50 ng/mL) IL-21, 100 ng/mL anti-CD40 or a titration of IL-21 mAb or soluble IL-21R/γc-Fc. Incorporation of 3H-thymidine during the final 16 h was measured. Results shown are representative of three independent experiments.
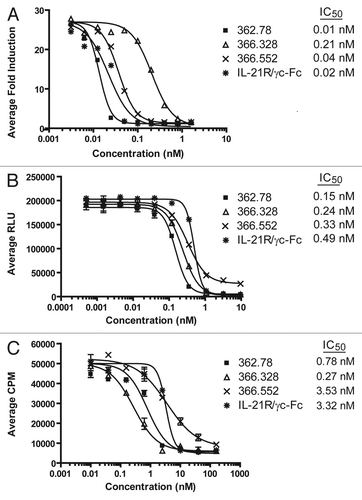
Figure 3 Neutralization of IL-21-mediated B cell differentiation and IgG1 production. (A) Inhibition of B-cell differentiation to a PC phenotype. A subset of primary human B cells, when cultured for 8 d with 25 ng/mL IL-21, 100 ng/mL anti-CD40 and 10 ng/mL IL-4, differentiate to an IgDlo, CD38+ PC phenotype. Addition of IL-21 neutralizing mAbs (produced from 1st round hybridoma clones) or soluble IL-21R blocked PC differentiation in a dose-dependent manner. Data for clone 362.78 is shown, though similar results were obtained with clones 362.328 and 366.552. Results shown are representative of two separate experiments. (B) IL-21 mAbs reduced IgG1 production. PC supernatants were collected at the end of the 8 d differentiation period and concentration (ng/mL) of total IgG1 measured. Inhibition of IL-21-mediated PC differentiation by the IL-21 mAbs correlated with a reduction in the amount of IgG1 produced.
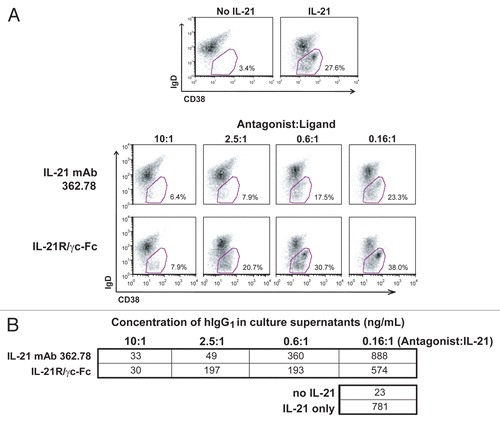
Figure 4 IL-21 mAb binds to and neutralizes native IL-21. (A) APC-labeled IL-21 mAb (362.78; purified from the 2nd round hybridoma clone) binds intracellular IL-21 in PMA+ionomycin-stimulated (upper panels), but not unstimulated (lower panels) human T cells. Total PBMC were stained with anti-CD3-FITC and counterstained with either mAb 362.78 (left panels), a commercially available APC-labeled positive control IL-21 mAb (IgG4) (right panels), or an APC-labeled IgG4 isotype control mAb (not shown). The APC staining from the isotype control mAb was used to set the quadrants shown. Results shown are representative of three independent experiments. (B) IL-21 mAb (clone 362.78) neutralized native IL-21-induced STAT3 phosphorylation. CD4+ T cells were isolated from four healthy donors and stimulated for 16 h with PMA+ionomycin to generate conditioned media containing native IL-21. Media was then incubated with a titration of IL-21 mAbs, cultured with BaF3/IL-21R cells for 15 min and pSTAT3 was measured. IL-21-mediated pSTAT3 from all four donors was blocked in a dose-dependent manner by IL-21 mAb.
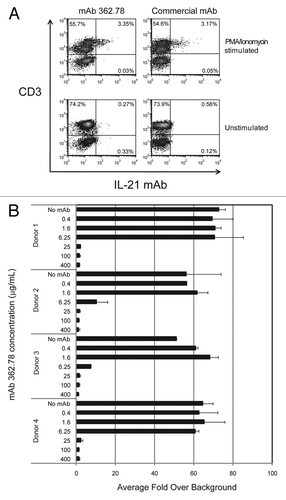
Figure 5 Binding kinetics correlate with efficacy in the IL-21-mediated pSTAT3 assay. Serial dilutions of IL-21 antagonists were pre-incubated with 96 pM IL-21 for 0 (filled squares), 5 (open triangles), 15 (filled inverted triangles) or 30 (open diamonds) min prior to initiation of the pSTAT3 assay. Recombinant CHO-expressed mAbs were used in these experiments. (A) With a relatively slow association rate, clone 366.552 requires a longer pre-incubation period than clone 362.78 or IL-21R to completely neutralize IL-21-mediated STAT3 phosphorylation. (B) IL-21 mAb clone 362.78 (very fast association rate) demonstrated the most potent inhibition of IL-21 mediated pSTAT3 with complete neutralization following 5, 15 and 30 min pre-incubation periods. (C) Soluble IL-21R/γc neutralized IL-21-mediated pSTAT3 in a time-dependent manner, but not as effectively as clone 362.78.
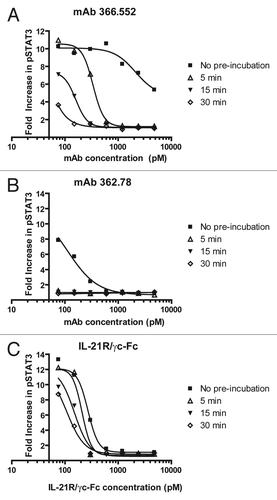
Figure 6 Representative mAbs from the neutralizing epitope bins (Bin #1, Bin #1/2 and Bin #2) were tested for competition with soluble homodimeric or heterodimeric IL-21R. Sensorgrams are shown for co-binding with (A) mAb 362.78 (Bin #1), (B) mAb 366.552 (Bin #1/2) and (C) mAb 366.328 (Bin #2). The lists of competitors tested against each mAb are shown in the boxes on the right.
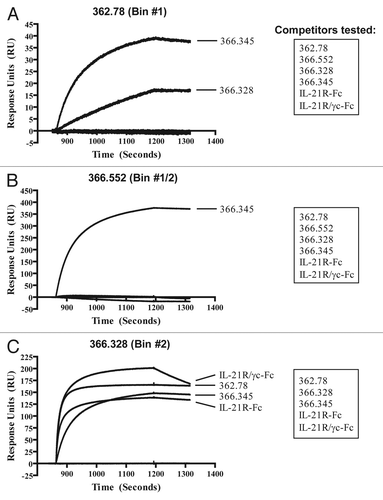
Figure 7 Amino acid sequence of mature hIL-21. Residues 1–133 of the mature hIL-21 polypeptide, lacking the signal sequence, are shown. Helices A–D are boxed, and the four synthetic peptides used to assess binding of the IL-21 mAbs by ELISA and western blot () are indicated. The two arrows denote the residues mutated in helix D (Q116D and I119D) of the IL-21 mutein; these two mutations do not affect the ability of the mutein to bind IL-21R, but prevent signaling through the IL-21R/γc complex, and are thought to be critical residues for γc binding.Citation57,Citation58
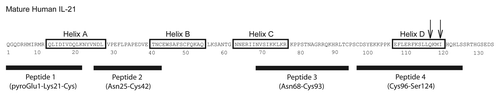
Table 1 Characteristics of key IL-21 mAbs and the soluble IL-21R/γc-Fc heterodimeric receptor
Table 2 Affinity and neutralizing activity of key IL-21 mAbs and the soluble IL-21R/γc-Fc heterodimeric receptor
Table 3 Summary of binding of key IL-21 mAbs to various forms of IL-21 and to peptides derived from IL-21