Abstract
The limb is one of the premier models for studying how a simple embryonic anlage develops into complex three-dimensional form. One of the key issues in the limb field has been to determine how the limb becomes patterned along its proximal (shoulder/hip) to distal (digits) axis. For decades it has been known that the apical ectodermal ridge (AER) plays a crucial role in distal outgrowth and patterning of the vertebrate embryonic limb. Most studies have explored the relationship between the AER and the progressive assignment of cell fates to mesenchyme along the proximal to distal (PD) axis. Comparatively few, however, have examined the additional role of the AER to regulate distal outgrowth of the limb and how this growth may also influence pattern along the PD axis. Here, I will review key studies that explore the role of growth in limb development. In particular, I will focus on a recent flurry of papers that examine the role of the Wnt/planar cell polarity (PCP) pathway in regulating directed growth of the limb mesenchyme. Finally, I will discuss a potential mechanism that relates the AER to the Wnt/PCP pathway and how directed growth can play a role in shaping the limb along the PD axis.
Acknowledgments
I thank Michael Stark, Marc Hansen and Aaron Smith for assistance with the manuscript. J.R.B. is supported by a grant from the NIH (HD060087).
Figures and Tables
Figure 1 Wnt5a signals through a Ror2/Vangl2 receptor complex. (A) In the absence of a Wnt5a signal, Ror2 and Vangl2 are inactive. Vangl2 is unphosphorylated. (B) In the presence of Wnt5a, a ternary complex with Ror2 and Vangl2 forms, which recruits CKiδ that, in turn, phosphorylates critical serine and threonine residues in Vangl2. Phosphorylated Vangl2 is critical for asymmetric localization of Vangl2 and could underlie polarized growth. See reference Citation51.
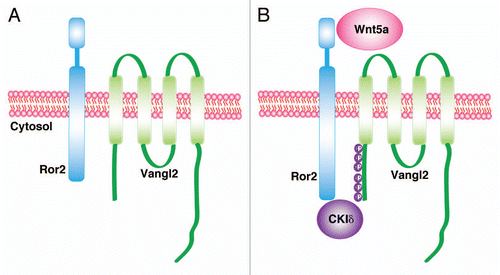
Figure 2 The dimensions of the AER change dramatically over time and underlie morphogenetic changes in the limb bud. (A) The pre-AER (orange), as indicated through Fgf8 expression, is short along the anteroposterior (AP) axis [anterior (a), left; posterior (p), right] and thick along the dorsoventral (DV) axis [dorsal (d), up; ventral (v), down], approaching a circular shape. Fgf8 emanating from the pre-AER elicits directed growth of the adjacent mesenchyme through the induction of Wnt5a/PCP signaling and through canonical Fgf/MAPK signaling. The “circular” AER recruits a cylindrical mass of mesenchyme that prefigures the shape of the stylopod. (B) Over time, the AER lengthens along the AP axis and thins along the DV, recruiting an oval solid mass of mesenchyme that presages the zeugopod. (C) At late stages, the AER extends dramatically along the AP axis and thins along the DV axis, recruiting a thin paddle of mesenchyme that prefigures the autopod. a, anterior; p, posterior; d, dorsal; v, ventral with respect to the limb.
![Figure 2 The dimensions of the AER change dramatically over time and underlie morphogenetic changes in the limb bud. (A) The pre-AER (orange), as indicated through Fgf8 expression, is short along the anteroposterior (AP) axis [anterior (a), left; posterior (p), right] and thick along the dorsoventral (DV) axis [dorsal (d), up; ventral (v), down], approaching a circular shape. Fgf8 emanating from the pre-AER elicits directed growth of the adjacent mesenchyme through the induction of Wnt5a/PCP signaling and through canonical Fgf/MAPK signaling. The “circular” AER recruits a cylindrical mass of mesenchyme that prefigures the shape of the stylopod. (B) Over time, the AER lengthens along the AP axis and thins along the DV, recruiting an oval solid mass of mesenchyme that presages the zeugopod. (C) At late stages, the AER extends dramatically along the AP axis and thins along the DV axis, recruiting a thin paddle of mesenchyme that prefigures the autopod. a, anterior; p, posterior; d, dorsal; v, ventral with respect to the limb.](/cms/asset/0d457c87-89dd-4610-a8e0-95d4f8761f6b/kogg_a_10919049_f0002.gif)
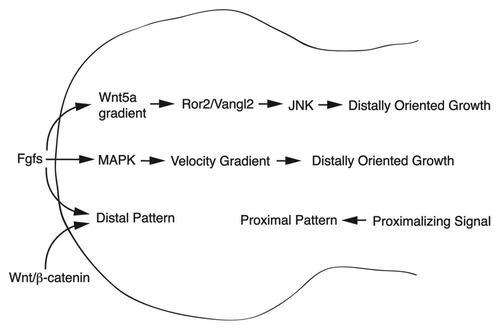
Figure 3 Coordination of growth and patterning in the limb. Fgfs that emanate from the AER play a central role in growth and patterning of the limb. First, they elicit a gradient of cell migration velocity (cells closest to the source of Fgf migrate with higher velocity than those further away), which generates a net distal bias in growth.Citation26 Fgfs (and perhaps other signals from the AER )Citation52,Citation53 are required for a gradient of Wnt5a expression in the distal limb, which is critical for a graded Vangl2 phosphorylation. A gradient of Vangl2 activity appears to underlie distally oriented growth. Fgf and canonical Wnt signals from the AER are required for the establishment of distal fates along the PD axis. According to this model, loss of Fgf signaling from the AER would disrupt both outgrowth and patterning of the limb. The loss of Wnt5a/PCP signaling would result in a severe disruption in outgrowth; however, some residual outgrowth and relatively normal patterning would occur.