Abstract
When exposed to stressful conditions, plants accumulate an array of metabolites, particularly amino acids. Amino acids have traditionally been considered as precursors to and constituents of proteins, and play an important role in plant metabolism and development. A large body of data suggests a positive correlation between proline accumulation and plant stress. Proline, an amino acid, plays a highly beneficial role in plants exposed to various stress conditions. Besides acting as an excellent osmolyte, proline plays three major roles during stress, i.e., as a metal chelator, an antioxidative defense molecule and a signaling molecule. Review of the literature indicates that a stressful environment results in an overproduction of proline in plants which in turn imparts stress tolerance by maintaining cell turgor or osmotic balance; stabilizing membranes thereby preventing electrolyte leakage; and bringing concentrations of reactive oxygen species (ROS) within normal ranges, thus preventing oxidative burst in plants. Reports indicate enhanced stress tolerance when proline is supplied exogenously at low concentrations. However, some reports indicate toxic effects of proline when supplied exogenously at higher concentrations. In this article, we review and discuss the effects of exogenous proline on plants exposed to various abiotic stresses. Numerous examples of successful application of exogenous proline to improve stress tolerance are presented. The roles played by exogenous proline under varying environments have been critically examined and reviewed.
Keywords: :
Introduction
Throughout their life cycle, plants are subjected to various types of environmental stresses, which include salinity, water deficit, temperature extremes, toxic metal ion concentration and UV radiations. These environmental factors limit the growth and productivity of plants to varying degrees, depending upon severity of stress. One of the stress response in the plants is the stimulated production of reactive oxygen species (ROS) e.g., OH·, O2·, H2O2 etc. These species cause considerable damage through peroxidation of membrane lipid components and also through direct interaction with various macromolecules. Cells have adapted different mechanisms to keep the ROS level in check. However, low ROS concentration participates in signal transduction mechanism.Citation1 These ROS are scavenged by low molecular weight antioxidative metabolites e.g., glutathione, ascorbic acid, α-tocopherol and antioxidative enzymes e.g., catalase, ascorbate peroxidase and superoxide dismutase. However, under different stress conditions the free radical generation exceeds the overall cellular antioxidative potential leading to oxidative stress, which contributes to adverse effects on plant growth.
In response to different stresses plants accumulate large quantities of different types of compatible solutes.Citation2 Compatible solutes are low molecular weight, highly soluble organic compounds that are usually non-toxic at high cellular concentrations. These solutes provide protection to plants from stress by contributing to cellular osmotic adjustment, ROS detoxification, protection of membrane integrity and enzymes/protein stabilization.Citation3-Citation5 These include proline, sucrose, polyols, trehalose and quaternary ammonium compounds (QACs) such as glycine betaine, alinine betaine, proline betaine and pipecolate betaine.Citation6,Citation7
The phenomenon of proline accumulation is known to occur under water deficit,Citation8 salinity,Citation9,Citation10 low temperature,Citation11 heavy metal exposureCitation12-Citation15 and UV radiations, etc. Apart from acting as osmolyte for osmotic adjustment, proline contributes to stabilizing sub-cellular structures (e.g., membranes and proteins), scavenging free radicals and buffering cellular redox potential under stress conditions.Citation3 It may also act as protein compatible hydrotrope,Citation16 alleviating cytoplasmic acidosis and maintaining appropriate NADP+/NADPH ratios compatible with metabolism.Citation17 In many plant species, proline accumulation under salt stress has been correlated with stress tolerance, and its concentration has been shown to be generally higher in salt tolerant than in salt sensitive plants.Citation18-Citation21 Its accumulation normally occurs in cytoplasm where it functions as molecular chaperons stabilizing the structure of proteins and its accumulation buffer cytosolic pH and maintains cell redox status. It has also been proposed that its accumulation may be part of stress signal influencing adaptive responses. Keeping in view the diverse roles of proline in plants, a comprehensive note on biosynthesis, transport, signaling and role in stress has been reviewed in this article.
Proline Biosynthesis, Signaling and Transport
Proline biosynthetic pathway had long been outlined before 40 y in Escherichia coli.Citation22 In plants, proline is synthesized by two pathways viz. glutamate pathway and orinithine pathway. The glutamate pathway accounts for major proline accumulation during osmotic stress. The proline is synthesized from glutamatic acid via intermediate ∆'-pyrroline-5-carboxylate (P5C). The reaction is being catalyzed by ∆'-pyrroline-5-carboxylate synthetase (P5CS) and ∆'-pyrroline-5-carboxylate reductase (P5CR) ().Citation23 P5CS is encoded by two genes whereas P5CR is encoded by only one in most plant species.Citation16,Citation24,Citation25 Proline catabolism occurs in mitochondria by means of the chronological action of proline dehydrogenase or proline oxidase (PDH or POX) producing P5C from proline and P5C dehydrogenase (P5CDH) converts P5C to glutamate. Two genes encode PDH, whereas a single P5CDH gene has been identified in Arabidopsis and tobacco (Nicotiana tabacum).Citation26-Citation29 PDH transcription is activated by rehydration and proline but repressed by dehydration, thus preventing proline degradation during abiotic stress.Citation27,Citation28 In an alternative pathway, proline can be synthesized from ornithine, which is transaminated to P5C by orinithine-δ-aminotransferase.Citation30 It has been suggested that ornithine pathway is important during seedling development and in some plants for stress-induced proline accumulation.Citation24,Citation31,Citation32 Accumulation of proline has been suggested to contribute to stress tolerance in many ways. As proline acts as the molecular chaperons it is able to maintain the protein integrity and enhancing the activities of different enzymes.Citation33 Numerous studies have reported proline as an antioxidant suggesting its role as ROS scavenger and singlet oxygen quencher.Citation34,Citation35
Figure 1. Proline metabolism in higher plants. Solid lines represents the biosynthetic pathway while catabolic pathways are shown with dashed lines. BAC, basic amino acid transporter (for arginine and ornithine exchange); Glu, glutamate; G/P, mitochondrial glutamate/proline antiporter; KG, α-ketoglutarate; P, mitochondrial proline transporter; Pi, inorganic phosphate; ProT, proline transporter; ?, unknown transporters. (Figure adapted from Szabados and Savoure [151])
![Figure 1. Proline metabolism in higher plants. Solid lines represents the biosynthetic pathway while catabolic pathways are shown with dashed lines. BAC, basic amino acid transporter (for arginine and ornithine exchange); Glu, glutamate; G/P, mitochondrial glutamate/proline antiporter; KG, α-ketoglutarate; P, mitochondrial proline transporter; Pi, inorganic phosphate; ProT, proline transporter; ?, unknown transporters. (Figure adapted from Szabados and Savoure [151])](/cms/asset/c9929eaf-fd5e-4a60-a83a-f75423da3af4/kpsb_a_10921949_f0001.gif)
Exogenous proline application reduces ROS levels in fungi and yeast, thus preventing programmed cell death,Citation36 and also prevents lipid peroxidation in alga cells exposed to heavy metals.Citation37 Pretreatment of proline also mitigated Hg2+ toxicity in rice (Oryza sativa) through ROS scavenging, such as H2O2.Citation38 Damaging effects of ROS on Photosystem II (PSII) can be reduced by proline in isolated thylakoid membranes (PSII).Citation39
Internal proline content can be determined by biosynthesis, catabolism and transport between cells and different cellular compartments. The biosynthetic enzymes (P5CS1, P5CS2 and P5CR) are predicted to be localized in the cytosol whereas a mitochondrial localization is predicted for the enzymes involved in proline catabolism (such as PDH1/ERD5, PDH2, P5CDH and OAT).Citation40
Intercellular transport of proline occurs between cytosol, chloroplasts and mitochondria as implied by compartmentalization of proline metabolism (). It has been reported that uptake of proline in mitochondria is an active process suggesting the existence of specific amino acid transporters.Citation41 These transporters have been identified in Arabidopsis thalianaCitation42 and in tomato pollen.Citation43 At least three transporters (Pro T1, Pro T2 and AAP6) of proline were identified in Arabidopsis thaliana based on C-DNA technology.Citation42 These transporters belong to amino acid permease (AAP) family and are expressed during stressful conditions. Pro T1 express ubiquitously but in Arabidopsis thaliana plants exposed to salinity stress, higher levels of Pro T1 were recorded in roots, stems and flowers. Young flowers showed highest expression particularly in floral stalk phloem. Under water or salinity stress, strong expression of Pro T2 was recorded whereas, AAP6 transcripts were detected mainly in sink tissues (roots, cauline leaves).Citation42 In the halophyte species Limonium latifolium, proline was sequestered to vacuoles in non-stressed plants, whereas, high proline content was detected in the cytosol of salt-stressed plants, suggesting the importance of de novo proline biosynthesis as well as transport for proline accumulation.Citation44
Proline metabolism has been studied for more than 40 y in plants, but little is known about the signaling pathways involved in its regulation. Proline biosynthetic pathway is activated and its catabolism repressed during dehydration, whereas rehydration regulates in opposite direction.Citation16,Citation26-Citation28,Citation32,Citation45 Chen et al.Citation46 indicated that proline accumulation in detached rice leaves upon exposure to excess Cu was due to proteolysis and increased activities of Δ1-pyrroline-5-carboxylate reductase or ornithine-δ-aminotransferase, which are enzymes of proline metabolism. It has also been revealed that Cu-induced proline synthesis and accumulation in detached rice leaves was mediated by ABA.Citation46 Zhang et al.Citation47 reported that Cu-induced proline synthesis is associated with NO generation. In this study the authors reported that exposure of Chlamydomonas reinhardtii to increasing concentration of Cu resulted in an increased synthesis of proline and a concomitant increase of intracellular NO level. The authors argued that this intracellular NO generation was involved in Cu-induced proline accumulation and signaling and this notion was largely based on the fact that the application of sodium nitroprusside (a potent NO donor) increased the activity and transcript amount of P5CS (a key enzyme of proline biosynthesis) in Cu-treated algae which was blocked if a NO scavenger instead of NO donor was used.Citation47 Further, it was reported in Senedesmus that the exogenous proline acts by detoxifying the ROS generated in response to the heavy metal (Cu or Zn) treatment rather than by improving the antioxidative defense system.Citation48 Similarly, Wang et al.Citation38 also demonstrated that the protective effect of proline against Hg toxicity in rice was through detoxifying ROS, generated in response to metal treatment.
Effect of Exogenous Proline on Growth of Plants under Varying Environment
When exposed to abiotic stress, plants experience growth inhibition or retardation. However, exogenous application of proline provided osmoprotection and also enhanced the growth of plants exposed to salt stress.Citation5,Citation49 Roy et al.Citation50 reported that proline, applied exogenously at a low concentration, ameliorated the adverse effects of salinity in rice. When added to the culture medium at low concentrations proline effectively alleviated salinity-induced decline in fresh weight and also reduced peroxidative damage to the lipid membranes in ground nut (Arachis hypogea); in contrast, higher proline concentrations did not prove beneficial.Citation51 In a similar study, Ehsanpour and FatahianCitation52 reported that exogenous application of proline to culture medium subjected to salinity stress resulted in an increase in dry weight and also increased the free proline content in callus cells of alfalfa (Medicago sativa). When applied exogenously to immature embryos of maize (Zea mays) proline stimulated somatic embryogenesis.Citation53-Citation55 Ali et al.Citation56 reported that the exogenous proline applied as spray treatment at seedling and/or at vegetative stage of Zea mays resulted in an enhanced growth under water deficient environment. Proline applied as pre-sowing seed soaking treatment alleviated the adverse effects generated by drought stress in Triticum aestivum resulting in an enhanced growth and yield characteristics.Citation57 Exogenous application of proline enhanced growth and also maintained the nutrient status by promoting the uptake of K+, Ca+, P and N in Zea mays plants exposed to drought stress.Citation58 Exogenous application of proline mitigated the cadmium induced inhibitory effects on the growth of cultured tobacco Bright Yellow-2 (BY-2) cells ().Citation59
Figure 2. Proline mediated intracellular redox-regulation as a multifaceted convergent strategy of different stresses. A balance of exogenous/endogenous molecules sets the required internal concentration of proline.
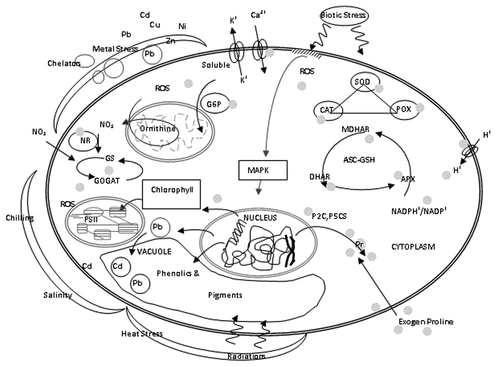
Effect of Exogenous Proline on Plant-Water Relations and Photosynthesis
Stress, in general, is known to alter plant-water relationsCitation60 which may affect water uptake, ascent of sap, stomatal functioningCitation61 and retardation of chlorophyll biosynthesisCitation62 and ultimately result in decreased photosynthesis. Decrease in leaf water potential is also associated with stress. The disturbance in plant-water relations due to heavy metal exposure triggers proline accumulation; this effect has been observed, for example, in response to Cd in Lectuca sativa.Citation63 Exogenous proline application to Vicia faba significantly increased leaf water potential during salinity stress.Citation64 The exogenous proline supplemented mitigated the reduction in photosynthetic activity and leaf water relations under salt stress in Olea europaea L. cv Chemlali, and the mitigating effect of proline was concentration dependent.Citation65 It is well-established that proline protects plants against stress by stabilizing the mitochondrial electron transport complex II,Citation66 membranes and proteinsCitation8,Citation67-Citation70 and enzymes such as RUBISCO.Citation71 When compared with other osmolytes such as glycine betaine, proline applied exogenously was highly effective in alleviating NaCl-generated stress in tobacco cells.Citation3 Both upper and lower stomata in Vicia faba responded to different concentrations of proline supplied exogenously either to detached leaves or to intact leaves.Citation72 The stomata on abaxial surfaces exhibited higher resistance than those on adaxial surfaces when treated exogenously with proline. Furthermore, lower concentrations of exogenous proline were even more effective in increasing stomatal resistance than that of ABA spray.Citation72 In the study by Rajagopal and Sinha,Citation73 exogenously applied proline maintained turgidity in leaves of barley and wheat undergoing stress.
Effect of Exogenous Proline on Oxidative Stress and the Antioxidant System
Plants continuously synthesize the reactive oxygen species (ROS) as a byproduct of various metabolic pathways.Citation74 ROS play a significant role in providing protection against harmful pathogens.Citation75-Citation77 They are also important in trachiary element formation, lignification and several other developmental processes.Citation78-Citation80 However, excessive levels of ROS result in oxidative damage to plants, e.g., nucleic acid damage, oxidation of proteins and lipids and degradation of chlorophyll pigments.Citation81-Citation84 Therefore, ROS generation should remain within plant-compatible limits. Under normal conditions ROS are scavenged by different antioxidant defense compounds.Citation85 However, when plants are exposed to various biotic and abiotic stresses increased generation of ROS occurs.Citation86-Citation88 This increased ROS level, besides causing the aforesaid damage, also cause K+ efflux from cells ().Citation89
Reports indicate that proline is responsible for scavenging the ROS and other free radicals.Citation35,Citation36,Citation90-Citation94 Proline, when applied exogenously to roots of Arabidopsis, resulted in a reduced level of ROS, indicating the ROS scavenging potential of proline.Citation95 Further, exogenous proline application also reduced ROS-induced K+ efflux.Citation95 Hoque et al.Citation96 reported that the activities of antioxidative enzymes viz. catalase (CAT), peroxidase (POX) and superoxide dismutase (SOD) were significantly enhanced when proline was applied exogenously in tobacco suspension cultures exposed to salinity stress.
Another important defense system of plants to protect cells against the destructive ROS (i.e., those generated in response to stress) is the ascorbate-glutathione (ASC-GSH) cycle.Citation93 Exogenous proline application upregulates the activities of enzymes in the ASC-GSH cycle. The activities of APX (ascorbate peroxidase), MDHAR (monohydro ascorbate reductase) and DHAR (dihydro ascorbate reductase) enzymes, which are the components of ASC-GSH cycle, were significantly enhanced by exogenous proline application in tobacco cultures exposed to salinity stress (Hoque et al. 2007). Kaul et al.Citation97 using in vitro studies, showed that exogenously applied l-proline proved to be a potent free radical (particularly ROS) scavenger. Hong et al.Citation91 concluded that the role of proline as a free radical scavenger is more important in alleviating stress than its role as a simple osmolyte. Islam et al.Citation59 showed that proline and betaine confer tolerance to cadmium stress in cultured tobacco cells by increasing the activities of SOD and CAT and also decreased the lipid peroxidation rate ().
Effect of Exogenous Proline on Plants Exposed to Salinity Stress
High salinity is a major problem faced by plants worldwide, which results in serious metabolic perturbations reducing crop productivity and yield. Salinity stress reduced growth and protein content in Pancratium maritium. This effect was, however, significantly reversed when proline was exogenously supplied.Citation98 Furthermore, the salinity-induced reduction in ubiquitin conjugate content and inhibition of the antioxidative enzymes catalase and peroxidase was significantly overcome in Pancratium maritimum when proline was supplied exogenously.Citation98 In a study performed by Gadallah,Citation64 exogenous proline application completely alleviated salinity-induced injury in Vicia faba. The membrane disruptions induced by salinity were also alleviated by exogenous proline in Vicia faba.Citation64 In the same study, exogenous proline application increased leaf chlorophyll content, leaf relative water content and overall plant growth. Exogenous proline application also increased percentage germination and root length in pea exposed to salinity stress.Citation99 In a study by Ehsanpour and FatahianCitation52 on callus cells of Medicago sativa, proline supplied exogenously to the culture medium subjected to salinity stress resulted in an increase in dry weight and also increased free proline content in the callus cells. Exogenous addition of proline to nutrient medium drastically decreased the oxidative damage to membranes caused by salinity in Mesembryanthemum crystallinum L. thus resulting in reduced lipid peroxidation rate but increased the chlorophyll content in the leaves of salt stressed plants.Citation100
Effect of Exogenous Proline on Plants Exposed to Radiation Stress
Harmful radiation such as UV-B (280–320 nm) is an important environmental factor which at higher levels adversely affects photosynthesis and other physiological processes.Citation71,Citation101 In response to UV-B radiation plants develop a variety of protective mechanisms, e.g., production of UV-B screening pigments and synthesis of protective compounds including flavonoids and proline.Citation80,Citation102,Citation103 The free radicals generated in response to UV-B exposure are scavenged by proline.Citation103,Citation104 The study by Arora and SaradhiCitation104 was further supported by the finding that barley seedlings pretreated with NaCl were more resistant to UV-B radiation. This increased tolerance to UV-B exposure was probably due to proline accumulation induced by salt stress conditions.Citation105 In addition, exogenous proline application to barley seedlings followed by UV-B exposure resulted in reduced chlorophyll/carotenoid ratio, oxygen evolution rate and photochemical efficiency of PS II, and also increased proline accumulation.Citation106 The reduced chlorophyll / carotenoid ratio by exogenous proline application was due to synthesis of pigments that provided protection to cells against UV-B radiation exposure ().Citation106
Effect of Exogenous Proline on Plants Exposed to Temperature Stress
Deviation from optimum temperature results in serious perturbations in plant growth and development. These perturbations include disruption of membranes due to lipid peroxidation, metabolic modifications, changes in protein content and enzymatic activity and electrolyte and amino acid leakage from cells. Application of chilling treatment to tropical and subtropical plants such as mung bean and soybean resulted in serious physiological and biochemical dysfunctions, most of which are mediated by active oxygen species.Citation88
Chilling sensitive seeds which are vulnerable to low temperatures during the early phase of imbibition result in decrease in percent germinationCitation107-Citation109; poor seedling growth and reduced plant productivity.Citation108 Hare et al.Citation110 observed that seed germination in Arabidopsis thaliana was enhanced by proline applied exogenously. An increase in plant growthCitation111 and crop productivityCitation112 under chilling stress conditions was also observed when proline was applied exogenously ().
Studies have revealed that the oxidative pentose phosphate pathway (OPPP) plays a critical role in triggering seed germination in various plant species. Botha et al.Citation113 and ShettyCitation114 suggested a link between OPPP and proline biosynthesis. This was further confirmed by Posmyk and Janas,Citation115 who found a positive correlation between endogenous proline content in seeds and germination when exposed to chilling stress.
Seed germination in Vigna radiata, inhibited by application of chilling stress of 5°C, was overcome when seeds were hydroprimed with proline. Further exogenous proline application had a dose-dependent stimulatory effect on germination of Vigna radiata seeds.Citation115 This effect of exogenous proline was attributed to membrane stabilizing potentialCitation34 which was changed from porous and leaky to stable and non-leaky.Citation116
Lipid peroxidation induced by chillingCitation117 was effectively overcome by exogenous proline application in Vigna radiata.Citation115 Exogenous proline acted as an active oxygen scavenger thereby overcoming the oxidative stress induced by chilling.Citation115 Van Swaaij et al.Citation118 determined that exogenous proline application resulted in increased frost tolerance in leaves of Solanum. Exogenous proline treatment also increased leaf proline content, thereby alleviating chill-induced stress. Besides acting as a free radical scavenger and stabilizing membranes, exogenous proline also acted as a source of nitrogen and carbon, thereby improving seedling growth and regeneration in Vigna radiata exposed to chilling stress.Citation115
Effect of Exogenous Proline on Plants Exposed to Heavy Metal Stress
Heavy metals such as Cd, Cu, Pb, Ni and Zn are common and destructive environmental threats encountered by plants. Plant uptake of excessive concentrations of heavy metals generates stress resulting in serious physiologic and structural disturbances. In response to heavy metal stress plants accumulate a large quantity of proline. Many plants have been shown to accumulate proline when exposed to heavy metal stress.Citation12,Citation13,Citation63,Citation119,Citation120 Besides acting as an osmoprotectant and ROS quencher, proline also acts as a heavy metal chelator, thereby alleviating heavy metal stress.Citation121 Proline induces the formation of phytochelatins which chelate with heavy metals like Cd thereby decreasing their toxicity.Citation122 Xu et al.Citation123 reported that the proline pre-treatment reduced the ROS level and also protected the plasma membrane of callus subjected to cadmium stress, thereby improving the cadmium tolerance resulting in regeneration of shoots in Solanum nigrum. Exogenous application of proline resulted in increase of its endogenous levels that antagonized the toxic effects of Selenium by improving the growth of seedlings. The damage caused by stress was reduced significantly with simultaneous increase in the activities of enzymatic and non- enzymatic antioxidants.Citation124 Heavy metal-tolerant populations of Deschampsia and Silene have been shown to contain a higher constitutive content of proline as compared with non-tolerant counterparts.Citation14,Citation125 In a study performed by Sharma et al.Citation126 exogenous proline protected the activity of glucose-6-phosphate dehydrogenase and nitrate reductase in vitro against inhibition by Cd and Zn. This protection was due to the formation of a proline-metal complex.Citation126 Similar complex-forming properties of proline were observed by Farago and MullenCitation121 where proline formed a complex with Cu in metal-tolerant Armaria ().
Research has demonstrated heavy metal-induced proline accumulation in higher as well as in lower plants. Enhanced proline accumulation was noted in response to heavy metals like Cd, Co, Zn and Pb in Cajanus cajan and Vigna mungoCitation119; Pb, Cd, Cu and Zn in Helianthus annuusCitation127; and Zn and Cu in Triticum aestivum.Citation13 Proline accumulation in response to heavy metal exposure has been observed in lower plants, e.g., algae. Proline accumulated when different algae were exposed to metals such as Cu in Anacystis nidulans,Citation128 Chlorella sp.Citation129 and Chlorella vulgaris.Citation37 Exogenous proline application to Chlorella vulgaris was found to counteract lipid peroxidation as well as K+ efflux observed after exposure to Cu, Cr, Ni and Zn.Citation37 Regardless of the above data, a survey of the literature indicates that generally little information is available on the effect of exogenous proline on plants exposed to heavy metal stress.
Effect of Exogenous Proline on Other Enzymes and Metabolites
Exogenous proline application besides enhancing the activity of antioxidative enzymes (CAT, POX and SOD)Citation96 is also known to enhance the activity of other enzymes. Nitrogenase activity in drought-stressed soybean nodules was significantly enhanced when proline (an osmolyte) was applied exogenously. However, when other osmolytes such as malate were tested there was no significant enhancement in drought-stressed nodule nitrogenase activity.Citation130 Proline is known to act as an enzyme protectant during abiotic stress conditions.Citation131,Citation132 This effect is further supported by the finding that exogenous proline application alleviated salt stress by upregulating the stress protective proteins in Pancratium maritinmumCitation98 and reducing oxidation of lipid membranes in tobacco.Citation94
It is well-established that stress results in increased proline accumulation in root nodules. Kohl et al.Citation133 reported that drought stress induced the activity of the enzymes of proline metabolism such as proline dehydrogenase (pro-DH) in bacteroids, suggesting that proline may be imported to the symbiosomes as a substrate for bacteroids during periods of stress. This finding was further confirmed by the observation that proline, when applied exogenously to the drought-stressed soybean nodules, was imported across the symbiosome membrane at a relatively faster rate, which was metabolized by the bacteroids and used to enhance nitrogenase activity in the nodules.Citation130 In addition proline protected the enzymes against heat, salinity or chilling stress under in vitro conditions.Citation134,Citation135 This is due to the fact that the 3-D structure of proteins is governed by hydrophobic/hydrophilic, ionic interactions and interactions between side chains of constituent amino acids. Proline could interfere with these side chain bonds and induce conformational changes in the enzyme protein and thus affect their activity ().Citation135,Citation136
GadallahCitation64 reported that the contents of soluble sugars, hydrolysable sugars and soluble proteins of salt-stressed Vicia faba significantly increased when proline was supplied exogenously. Posmyk and JanasCitation115 reported that Vigna radiata seedlings exposed to chilling stress when supplied exogenously with proline resulted in an increase in phenolics content. Phenolics like endogenous proline act as a free radical scavenger thereby overcoming oxidative stress.Citation137,Citation138
Exogenous Proline — A Comparison with Other Amino Acids
Accumulation of large quantities of proline is an adaptive response of plants to various biotic and abiotic stresses.Citation139-Citation145 Singh et al.Citation146 was probably the first to assign a correlation between proline accumulation and drought resistance in barley cultivars.Citation147 However, reports have shown accumulation of other free amino acids under stress conditions e.g., aspartic acid, glutamic acid and glutamine in cottonCitation148; asparagine, aspartic acid, serine and glycine in maizeCitation149,Citation150 and ornithine, arginine and glutamic acid in detached rice leaves.Citation151
Proline administered exogenously in nutrient solution to wheat exposed to osmotic stress delayed wilting.Citation73 Subsequently RajagopalCitation72 showed that in Vicia faba exogenous proline decreased stomatal conductance and also increased uptake of exogenous proline into leaves. However, Thakur and RaiCitation152 observed that exogenous application of alanine, serine and asparagine also delayed wilting under stress conditions in maize. Exogenous proline inhibited stomatal opening in Vicia faba whereas other amino acids such as histidine, methionine, aspartic acid, glutamic acid, asparagine and gutamine promoted stomatal opening.Citation153 Rai and KumariCitation154 monitored the passage of H+ or OH- ions across the Vinca petal membrane and observed that exogenous proline decreased membrane permeability for H+ or OH- ions. A similar response was generated when arginine, asparagine, glutamine, alanine and leucine were tested.
Exogenous proline application to Phaseolus seedlings promoted calcium uptake whereas alanine, aspartic acid, glutamic acid and tryptophan inhibited Ca uptake.Citation155 Like proline the amino acids histidine, γ-amino butyric acid, hydroxy proline, leucine, glutamine and phenylalanine enhanced K+ uptake. However, threonine, aspartic acid and alanine did not affect K+ uptake.
Santos et al.Citation156 reported that exogenous proline acts as a nitrogen source. They concluded that diverse morphogenetic processes such as embryogenesis and organogenesis vary in their demand of exogenous nitrogen (especially in the form of proline) which would also act as an osmoregulator. Seed germination in Sinapis alba was least inhibited by proline when applied exogenously at a concentration of 3.5x10−2 M whereas, at the same concentration, alanine, glutamic acid and aspartic acid proved highly inhibitory.Citation157 The authors also indicated that proline was about 300 times more soluble in water than other amino acids and thus acted as a comparatively non-toxic osmolyte. Proline level declined in response to nitrogen deficiency in Phaseolus vulgaris because of the stimulation of proline dehydrogenase. However, under the conditions of adequate nitrogen, proline level increased due to the activation of ornithine δ-aminotransferase.Citation158
Proline Toxicity in Plants
Despite the beneficial effects of exogenous proline application, it imparts toxic effects as well if over-accumulated or applied at excessive concentrations. Such negative effects of exogenous proline were observed in tomato, where an imbalance in inorganic ions was observed.Citation159 Proline at a low concentration (e.g., 30 mM) when applied exogenously, ameliorated the adverse effects of salinity on early seedling growth in rice, whereas at higher concentrations (40–50 mM) proline resulted in toxic effects and poor plant growth.Citation50 In a study by Hare et al.,Citation160 it was shown that proline applied exogenously at a low concentration enhanced in vitro shoot organogenesis in Arabidopsis hypocotyls explants, whereas growth was inhibited at higher concentrations. An explanation for this toxic effect of exogenous proline is attributed to the fact that lower concentrations activated a cycle of cytosolic proline synthesis from glutamate and mitochondrial proline degradation, which simultaneously provided NADP+ to drive cytosolic purine biosynthesis and reducing equivalents for mitochondrial ADP phosphorylation.Citation161 An induction by exogenous proline of the Arabidopsis gene which encodes proline dehydrogenase (PDH)Citation27 is consistent with this hypothesis. However, at higher levels of exogenous proline, feedback inhibition of Δ1-pyroline-5-carboxylate synthetase (P5CS)Citation162,Citation163 blocked the biosynthetic portion of this cycle and thereby inhibited organogenesis, as in Arabidopsis.Citation160 The toxic effects of exogenous proline were also observed by Rodriguez and HeyserCitation164 where growth in suspension culture of saltgrass (Distichlis spicata) was inhibited when proline was applied exogenously at a high concentration. This treatment also decreased proline biosynthesis.
Chen and KaoCitation165 suggest that high concentrations of proline mimic the toxic effects of Cd in rice seedlings. Nanjo et al.Citation166 evaluated proline toxicity in Arabidopsis T-DNA tagged mutant pdh that was defective in pro dehydrogenase (At ProDH), responsible for catalyzing the first step of proline catabolism. This pdh mutant was hypersensitive to exogenous l-proline at concentrations < 10 mM whereas the wild type grew normally at such concentrations.
Besides the above toxic effects of exogenous proline, it has been shown to destabilize the DNA helix, lower the DNA melting point, increase susceptibility to S1 nuclease and increase insensitivity to DNAase1, when supplied at high concentrations.Citation33
Conclusions
1. Proline, an amino acid, plays an important role in plants. It protects the plants from various stresses and also helps plants to recover from stress more rapidly.
2. When applied exogenously to plants exposed to stress, Proline results in increased growth and other physiological characteristics of plants.
3. Exogenous proline scavenges the ROS generated in plants under various biotic and abiotic stresses.
4. Exogenous proline application affects plant-water relations by maintaining turgidity of cells under stress, and also increases the rate of photosynthesis.
5. Exogenous proline application to plants protects them from harmful radiation such as. UV-B.
6. Low concentrations of exogenous proline protect plants from salinity, drought and temperature stress. However, higher doses will impart toxic effects.
Despite the diverse physio-morphological effects of exogenous proline on plants, much work is still required for a more complete understanding of its effects on plant response to external stress. Keeping in view the potential of exogenous proline as a ROS scavenger, it may become a useful tool to counter the adverse effect of stressful environments thereby decreasing annual losses to agriculture.
Related Research Data
References
- Foyer CH, Noctor G. Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell 2005; 17:1866 - 75; 10.1105/tpc.105.033589; PMID: 15987996
- Serraj R, Sinclair TR. Osmolyte accumulation: can it really help increase crop yield under drought conditions?. Plant Cell Environ 2002; 25:333 - 41; 10.1046/j.1365-3040.2002.00754.x; PMID: 11841674
- Ashraf M, Foolad MR. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 2007; 59:206 - 16; 10.1016/j.envexpbot.2005.12.006
- Bohnert HJ, Jensen RG. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol 1996; 14:89 - 97; 10.1016/0167-7799(96)80929-2
- Yancey PH. Compatible and counteracting solutes In: Strange K ed. Cellular and Molecular Physiology of Cell Volume Regulation. Boca Raton, FL: CRC Press, 1994:81-109.
- Ashraf M, Harris PJC. Potential biochemical indicators of salinity tolerance in plants. Plant Sci 2004; 166:3 - 16; 10.1016/j.plantsci.2003.10.024
- Rhodes D, Hanson AD. Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu Rev Plant Physiol Plant Mol Biol 1993; 44:357 - 84; 10.1146/annurev.pp.44.060193.002041
- Hare PD, Cress WA, Van Staden J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ 1998; 21:535 - 53; 10.1046/j.1365-3040.1998.00309.x
- Munns R. Genes and salt tolerance: bringing them together. New Phytol 2005; 167:645 - 63; 10.1111/j.1469-8137.2005.01487.x; PMID: 16101905
- Rhodes D, Nadolska-Orczyk A, Rich PJ. Salinity, osmolytes and compatible solutes In: Lauchli A, Luttge U, eds. Salinity, Environment, Plant, Molecules. Netherlands: Al-Kluwer Academic Publishers, 2002:181-204.
- Naidu BP, Paleg LG, Aspinall D, Jennings AC, Jones GP. Amino acid and glycine betaine accumulation in cold-stressed wheat seedlings. Phytochem 1991; 30:407 - 9; 10.1016/0031-9422(91)83693-F
- Bassi R, Sharma SS. Changes in proline content accompanying the uptake of zinc and copper by Lemna minor.. Ann Bot (Lond) 1993; 72:151 - 4; 10.1006/anbo.1993.1093
- Bassi R, Sharma SS. Proline accumulation in wheat seedlings exposed to zinc and copper. Phytochem 1993; 33:1339 - 42; 10.1016/0031-9422(93)85086-7
- Schat H, Sharma SS, Vooijs R. Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris.. Physiol Plant 1997; 101:477 - 82; 10.1111/j.1399-3054.1997.tb01026.x
- Sharma SS, Dietz KJ. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J Exp Bot 2006; 57:711 - 26; 10.1093/jxb/erj073; PMID: 16473893
- Strizhov N, Abrahám E, Okrész L, Blickling S, Zilberstein A, Schell J, et al. Differential expression of two P5CS genes controlling proline accumulation during salt-stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis.. Plant J 1997; 12:557 - 69; 10.1046/j.1365-313X.1997.00557.x; PMID: 9351242
- Hare PD, Cress WA. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul 1997; 21:79 - 102; 10.1023/A:1005703923347
- Fougère F, Le Rudulier D, Streeter JG. Effects of salt stress on amino acid, organic acid, and carbohydrate composition of roots, bacteroids, and cytosol of alfalfa (Medicago sativa L.). Plant Physiol 1991; 96:1228 - 36; 10.1104/pp.96.4.1228; PMID: 16668324
- Gangopadhyay G, Basu S, Mukherjee B, Gupta S. Effects of salt and osmotic shocks on unadapted and adapted callus lines of tobacco. Plant Cell Tissue Organ Cult 1997; 49:45 - 52; 10.1023/A:1005860718585
- Madan S, Nainawatee HS, Jain RK, Chowdhury JB. Proline and proline metabolising enzymes in in-vitro selected NaCl-tolerant Brassica juncea L. under salt stress. Ann Bot (Lond) 1995; 76:51 - 7; 10.1006/anbo.1995.1077
- Petrusa LM, Winicov I. Proline status in salt tolerant and salt sensitive alfalfa cell lines and plants in response to NaCl. Plant Physiol Biochem 1997; 35:303 - 10
- Vogel HJ, Davis BD. Glutamic- d-semialdehyde and Δ'-pyrroline-5-carboxylic acid, intermediates in the biosynthesis of proline. J Am Chem Soc 1952; 74:109 - 12; 10.1021/ja01121a025
- Sekhar PN, Amrutha RN, Sangam S, Verma DP, Kishor PB. Biochemical characterization, homology modeling and docking studies of ornithine delta-aminotransferase--an important enzyme in proline biosynthesis of plants. J Mol Graph Model 2007; 26:709 - 19; 10.1016/j.jmgm.2007.04.006; PMID: 17604199
- Armengaud P, Thiery L, Buhot N, Grenier-De March G, Savouré A. Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol Plant 2004; 120:442 - 50; 10.1111/j.0031-9317.2004.00251.x; PMID: 15032841
- Verbruggen N, Villarroel R, Van Montagu M. Osmoregulation of a pyrroline-5-carboxylate reductase gene in Arabidopsis thaliana.. Plant Physiol 1993; 103:771 - 81; 10.1104/pp.103.3.771; PMID: 8022935
- Deuschle K, Funck D, Hellmann H, Däschner K, Binder S, Frommer WB. A nuclear gene encoding mitochondrial Delta-pyrroline-5-carboxylate dehydrogenase and its potential role in protection from proline toxicity. Plant J 2001; 27:345 - 56; 10.1046/j.1365-313X.2001.01101.x; PMID: 11532180
- Kiyosue T, Yoshiba Y, Yamaguchi-Shinozaki K, Shinozaki K. A nuclear gene encoding mitochondrial proline dehydrogenase, an enzyme involved in proline metabolism, is upregulated by proline but downregulated by dehydration in Arabidopsis.. Plant Cell 1996; 8:1323 - 35; PMID: 8776899
- Verbruggen N, Hua XJ, May M, Van Montagu M. Environmental and developmental signals modulate proline homeostasis: evidence for a negative transcriptional regulator. Proc Natl Acad Sci U S A 1996; 93:8787 - 91; 10.1073/pnas.93.16.8787; PMID: 8710950
- Ribarits A, Abdullaev A, Tashpulatov A, Richter A, Heberle-Bors E, Touraev A. Two tobacco proline dehydrogenases are differentially regulated and play a role in early plant development. Planta 2007; 225:1313 - 24; 10.1007/s00425-006-0429-3; PMID: 17106685
- Verbruggen N, Hermans C. Proline accumulation in plants: a review. Amino Acids 2008; 35:753 - 9; 10.1007/s00726-008-0061-6; PMID: 18379856
- Roosens NH, Thu TT, Iskandar HM, Jacobs M. Isolation of the ornithine-delta-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana.. Plant Physiol 1998; 117:263 - 71; 10.1104/pp.117.1.263; PMID: 9576796
- Xue X, Liu A, Hua X. Proline accumulation and transcriptional regulation of proline biosynthesis and degradation in Brassica napus.. BMB Rep 2009; 42:28 - 34; 10.5483/BMBRep.2009.42.1.028; PMID: 19192390
- Rajendrakumar CSV, Suryanarayana T, Reddy AR. DNA helix destabilization by proline and betaine: possible role in the salinity tolerance process. FEBS Lett 1997; 410:201 - 5; 10.1016/S0014-5793(97)00588-7; PMID: 9237629
- Matysik J. Alia., Bhalu B, Mohanty P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr Sci 2002; 82:525 - 32
- Smirnoff N, Cumbes QJ. Hydroxyl radical scavenging activity of compatible solutes. Phytochem 1989; 28:1057 - 60; 10.1016/0031-9422(89)80182-7
- Chen C, Dickman MB. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii.. Proc Natl Acad Sci U S A 2005; 102:3459 - 64; 10.1073/pnas.0407960102; PMID: 15699356
- Mehta SK, Gaur JP. Heavy-metal-induced proline accumulation and its role in ameliorating metal toxicity in Chlorella vulgaris. New Phytol 1999; 143:253 - 9; 10.1046/j.1469-8137.1999.00447.x
- Wang F, Zeng B, Sun Z, Zhu C. Relationship between proline and Hg2+-induced oxidative stress in a tolerant rice mutant. Arch Environ Contam Toxicol 2009; 56:723 - 31; 10.1007/s00244-008-9226-2; PMID: 18787889
- Alia PPS, Mohanty P. Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J Photochem Photobiol 1997; 38:253 - 7; 10.1016/S1011-1344(96)07470-2
- Szabados L, Savouré A. Proline: a multifunctional amino acid. Trends Plant Sci 2010; 15:89 - 97; 10.1016/j.tplants.2009.11.009; PMID: 20036181
- Yu C, Claybrook DL, Huang AH. Transport of glycine, serine, and proline into spinach leaf mitochondria. Arch Biochem Biophys 1983; 227:180 - 7; 10.1016/0003-9861(83)90361-2; PMID: 6416178
- Rentsch D, Hirner B, Schmelzer E, Frommer WB. Salt stress-induced proline transporters and salt stress-repressed broad specificity amino acid permeases identified by suppression of a yeast amino acid permease-targeting mutant. Plant Cell 1996; 8:1437 - 46; PMID: 8776904
- Schwacke R, Grallath S, Breitkreuz KE, Stransky E, Stransky H, Frommer WB, et al. LeProT1, a transporter for proline, glycine betaine, and gamma-amino butyric acid in tomato pollen. Plant Cell 1999; 11:377 - 92; PMID: 10072398
- Gagneul D, Aïnouche A, Duhazé C, Lugan R, Larher FR, Bouchereau A. A reassessment of the function of the so-called compatible solutes in the halophytic plumbaginaceae Limonium latifolium.. Plant Physiol 2007; 144:1598 - 611; 10.1104/pp.107.099820; PMID: 17468212
- Yoshiba Y, Kiyosue T, Katagiri T, Ueda H, Mizoguchi T, Yamaguchi-Shinozaki K, et al. Correlation between the induction of a gene for delta 1-pyrroline-5-carboxylate synthetase and the accumulation of proline in Arabidopsis thaliana under osmotic stress. Plant J 1995; 7:751 - 60; 10.1046/j.1365-313X.1995.07050751.x; PMID: 7773306
- Chen CT, Chen L, Lin CC, Kao CH. Regulation of proline accumulation in detached rice leaves exposed to excess copper. Plant Sci 2001; 160:283 - 90; 10.1016/S0168-9452(00)00393-9; PMID: 11164600
- Zhang LP, Mehta SK, Liu ZP, Yang ZM. Copper-induced proline synthesis is associated with nitric oxide generation in Chlamydomonas reinhardtii.. Plant Cell Physiol 2008; 49:411 - 9; 10.1093/pcp/pcn017; PMID: 18252734
- Tripathi BN, Gaur JP. Relationship between copper- and zinc-induced oxidative stress and proline accumulation in Scenedesmus sp. Planta 2004; 219:397 - 404; 10.1007/s00425-004-1237-2; PMID: 15014994
- Csonka LN, Hanson AD. Prokaryotic osmoregulation: genetics and physiology. Annu Rev Microbiol 1991; 45:569 - 606; 10.1146/annurev.mi.45.100191.003033; PMID: 1741624
- Roy D, Basu N, Bhunia A, Banerjee S. Counteraction of exogenous L-proline with NaCl in salt-sensitive cultivar of rice. Biol Plant 1993; 35:69 - 72; 10.1007/BF02921122
- Jain MJ, Mathur GM, Koul SK, Sarin NS. Ameliorative effects of proline on salt stress-induced lipid peroxidation in cell lines of groundnut Arachis hypogaea L.). Plant Cell Rep 2001; 20:463 - 8; 10.1007/s002990100353
- Ehsanpour AA, Fatahian N. Effects of salt and proline on Medicago sativa callus. Plant Cell Tissue Organ Cult 2003; 73:53 - 6; 10.1023/A:1022619523726
- Armstrong CL, Green CE. Establishment and maintenance of friable, embryogenic maize callus and the involvement of L-proline. Planta 1985; 164:207 - 14; 10.1007/BF00396083
- Claparols I, Santos MA, Torné JM. Influence of some exogenous amino acids on the production of maize embryogenic callus and on endogenous amino acid content. Plant Cell Tissue Organ Cult 1993; 34:1 - 11; 10.1007/BF00048457
- Duncan DR, Widholm JM. Proline accumulation and its implication in cold tolerance of regenerable maize callus. Plant Physiol 1987; 83:703 - 8; 10.1104/pp.83.3.703; PMID: 16665311
- Ali Q, Ashraf M, Athar HUR. Exogenously applied proline at different growth stages enhances growth of two maize cultivars grown under water deficit conditions. Pak J Bot 2007; 39:1133 - 44
- Kamran M, Shahbaz M, Ashraf M, Akram NA. Alleviation of drought- induced adverse effects in spring wheat (Triticum aestivum L.) using proline as a pre-sowing seed treatment. Pak J Bot 2009; 41:621 - 32
- Ali Q, Ashraf M, Shahbaz M, Humera H. Ameliorating effect of foliar applied proline on nutrient uptake in water stressed maize (Zea mays L.) plants. Pak J Bot 2008; 40:211 - 9
- Islam MM, Hoque MA, Okuma E, Banu MN, Shimoishi Y, Nakamura Y, et al. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J Plant Physiol 2009; 166:1587 - 97; 10.1016/j.jplph.2009.04.002; PMID: 19423184
- Barcelo J, Poschenrieder C. Plant water relations as affected by heavy metal stress: A review. J Plant Nutr 1990; 13:1 - 37; 10.1080/01904169009364057
- Poschenrieder CH, Barcelo J. Water relations in heavy metal stressed plants In: Prasad MNV ed. Heavy metal stress in plants: from biomolecules to ecosystems. New Delhi: Narosa Publishing House, 2004:249-70.
- Singh PK, Tewari RK. Cadmium toxicity induced changes in plant water relations and oxidative metabolism of Brassica juncea L. plants. J Environ Biol 2003; 24:107 - 12; PMID: 12974420
- Costa G, Morel JL. Water relations, gas exchange and amino acid content in Cd-treated lettuce. Plant Physiol Biochem 1994; 32:561 - 70
- Gadallah MAA. Effects of proline and glycinebetaine on Vicia faba responses to salt stress. Biol Plant 1999; 42:249 - 57; 10.1023/A:1002164719609
- Ben Ahmed C, Ben Rouina B, Sensoy S, Boukhriss M, Ben Abdullah F. Exogenous proline effects on photosynthetic performance and antioxidant defense system of young olive tree. J Agric Food Chem 2010; 58:4216 - 22; 10.1021/jf9041479; PMID: 20210359
- Hamilton EW 3rd, Heckathorn SA. Mitochondrial adaptations to NaCl. Complex I is protected by anti-oxidants and small heat shock proteins, whereas complex II is protected by proline and betaine. Plant Physiol 2001; 126:1266 - 74; 10.1104/pp.126.3.1266; PMID: 11457977
- Holmström KO, Somersalo S, Mandal A, Palva TE, Welin B. Improved tolerance to salinity and low temperature in transgenic tobacco producing glycine betaine. J Exp Bot 2000; 51:177 - 85; 10.1093/jexbot/51.343.177; PMID: 10938824
- Mansour MMF. Protection of plasma membrane of onion epidermal cells by glycinebetaine and proline against NaCl stress. Plant Physiol Biochem 1998; 36:767 - 72; 10.1016/S0981-9428(98)80028-4
- McNeil SD, Nuccio ML, Hanson AD. Betaines and related osmoprotectants. Targets for metabolic engineering of stress resistance. Plant Physiol 1999; 120:945 - 50; 10.1104/pp.120.4.945; PMID: 10444077
- Paleg LG, Stewart GR, Bradbeer JW. Proline and glycine betaine influence protein solvation. Plant Physiol 1984; 75:974 - 8; 10.1104/pp.75.4.974; PMID: 16663771
- Allen DJ, McKee IF, Farage PK, Baker NR. Analysis of limitations to CO2 assimilation on exposure of leaves of two Brassica napus cultivars to UV-B. Plant Cell Environ 1997; 20:633 - 40; 10.1111/j.1365-3040.1997.00093.x
- Rajagopal V. The influence of exogenous proline on the stomatal resistance in Vicia faba.. Physiol Plant 1981; 52:292 - 6; 10.1111/j.1399-3054.1981.tb08508.x
- Rajagopal V, Sinha SK. Influence of exogenously supplied proline on relative water content in wheat and barley. Indian J Exp Biol 1980; 18:1523 - 4
- Foyer CH, Harbinson JC. Oxygen metabolism and the regulation of photosynthetic electron transport. In: Foyer CH, Mullineaux PM eds. Causes of photooxidative stress and amelioration of defense systems in plants., Boca Raton, FL, USA: CRC Press; 1994:1-42.
- Alvarez ME, Lamb C. Oxidative burst-mediated defense responses in plant disease resistance. In: Scandalios JG ed. Oxidative stress and the molecular biology of antioxidant defenses. Cold Spring Harbor: Cold spring Harbor Press, 1997:815-39.
- Bolwell GP, Bindschedler LV, Blee KA, Butt VS, Davies DR, Gardner SL, et al. The apoplastic oxidative burst in response to biotic stress in plants: a three-component system. J Exp Bot 2002; 53:1367 - 76; 10.1093/jexbot/53.372.1367; PMID: 11997382
- Doke N. The oxidative burst: Role in signal transduction and plant stress. In: Scandalios JG ed. Oxidative stress and the molecular biology of antixoidant defenses. Cold Spring Harbor: Cold Spring Harbor Press, 1997:785-813.
- Fath A, Bethke P, Beligni V, Jones R. Active oxygen and cell death in cereal aleurone cells. J Exp Bot 2002; 53:1273 - 82; 10.1093/jexbot/53.372.1273; PMID: 11997375
- Jacobson MD. Reactive oxygen species and programmed cell death. Trends Biochem Sci 1996; 21:83 - 6; PMID: 8882579
- Tevini M, Braun J, Fieser G. The protective functions of epidermal layer of rice seedlings against ultraviolet-B radiation. Photochem Photobiol 1991; 53:329 - 33; 10.1111/j.1751-1097.1991.tb03636.x
- Davies KJ. Protein damage and degradation by oxygen radicals. I. general aspects. J Biol Chem 1987; 262:9895 - 901; PMID: 3036875
- Fridovich I. Superoxide dismutases. Adv Enzymol Relat Areas Mol Biol 1986; 58:61 - 97; PMID: 3521218
- Imlay JA, Linn S. DNA damage and oxygen radical toxicity. Science 1988; 240:1302 - 9; 10.1126/science.3287616; PMID: 3287616
- Schützendübel A, Polle A. Plant responses to abiotic stresses: heavy metal-induced oxidative stress and protection by mycorrhization. J Exp Bot 2002; 53:1351 - 65; 10.1093/jexbot/53.372.1351; PMID: 11997381
- Alscher RG, Donahue JL, Cramer CL. Reactive oxygen species and antioxidants: Relationships in green cells. Physiol Plant 1997; 100:224 - 33; 10.1111/j.1399-3054.1997.tb04778.x
- Dat J, Vandenabeele S, Vranová E, Van Montagu M, Inzé D, Van Breusegem F. Dual action of the active oxygen species during plant stress responses. Cell Mol Life Sci 2000; 57:779 - 95; 10.1007/s000180050041; PMID: 10892343
- Mano J. Early events in environmental stresses in plants: Induction mechanisms of oxidative stress. In: Inze D, Montagu MV eds. Oxidative Stress in Plants. London: Taylor & Francis, 2002:217-46.
- Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 2002; 7:405 - 10; 10.1016/S1360-1385(02)02312-9; PMID: 12234732
- Shabala S. Non-invasive microelectrode ion flux measurements in plant stress physiology. In: Volkov A ed. Plant Electrophysiology – Theory and Methods. Berlin: Springer-Verlag, 2006:35–71.
- Bohnert HJ, Nelson DE, Jensen RG. Adaptation to environmental stresses. Plant Cell Rep 1995; 7:1099 - 111
- Hong Z, Lakkineni K, Zhang Z, Verma DP. Removal of feedback inhibition of delta(1)-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol 2000; 122:1129 - 36; 10.1104/pp.122.4.1129; PMID: 10759508
- Niyogi KK. PHOTOPROTECTION REVISITED: Genetic and Molecular Approaches. Annu Rev Plant Physiol Plant Mol Biol 1999; 50:333 - 59; 10.1146/annurev.arplant.50.1.333; PMID: 15012213
- Noctor G, Foyer CH. Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 1998; 49:249 - 79; 10.1146/annurev.arplant.49.1.249; PMID: 15012235
- Okuma E, Murakami Y, Shimoishi Y, Tada M, Murata Y. Effects of exogenous application of proline and betaine on the growth of tobacco cultured cells under saline conditions. Soil Sci Plant Nutr 2004; 50:1301 - 5; 10.1080/00380768.2004.10408608
- Cuin TA, Shabala S. Compatible solutes reduce ROS-induced potassium efflux in Arabidopsis roots. Plant Cell Environ 2007; 30:875 - 85; 10.1111/j.1365-3040.2007.01674.x; PMID: 17547658
- Hoque MA, Banu MN, Okuma E, Amako K, Nakamura Y, Shimoishi Y, et al. Exogenous proline and glycinebetaine increase NaCl-induced ascorbate-glutathione cycle enzyme activities, and proline improves salt tolerance more than glycinebetaine in tobacco Bright Yellow-2 suspension-cultured cells. J Plant Physiol 2007; 164:1457 - 68; 10.1016/j.jplph.2006.10.004; PMID: 17223225
- Kaul S, Sharma SS, Mehta IK. Free radical scavenging potential of L-proline: evidence from in vitro assays. Amino Acids 2008; 34:315 - 20; 10.1007/s00726-006-0407-x; PMID: 17086481
- Khedr AHA, Abbas MA, Wahid AAA, Quick WP, Abogadallah GM. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J Exp Bot 2003; 54:2553 - 62; 10.1093/jxb/erg277; PMID: 14512386
- Bar-Nun N, Poljakoff-Mayber A. Salinity stress and the content of proline in roots of Pisum sativum and Tamarix tetragyna.. Ann Bot (Lond) 1977; 41:173 - 9
- Shevyakova N, Bakulina E, Kuznetsov V. Proline antioxidant role in the common ice plant subjected to salinity and paraquat treatment inducing oxidative stress. Russ J Plant Physiol 2009; 56:663 - 9; 10.1134/S1021443709050124
- Rajendiran K, Ramanujam MP. Alleviation of ultraviolet-B radiation-induced growth inhibition of green gram by triadimefon. Biol Plant 2003; 46:621 - 4; 10.1023/A:1024840301092
- Day TA, Vogelmann TC. Alterations in photosynthesis and pigment distributions in pea leaves following UV-B exposure. Physiol Plant 1995; 94:433 - 40; 10.1111/j.1399-3054.1995.tb00950.x
- Saradhi PP, Alia, Arora S, Prasad KV. Proline accumulates in plants exposed to UV radiation and protects them against UV induced peroxidation. Biochem Biophys Res Commun 1995; 209:1 - 5; 10.1006/bbrc.1995.1461; PMID: 7726821
- Arora S, Saradhi PP. Light induced enhancement in proline levels under stress is regulated by non-photosynthetic events. Biol Plant 2002; 45:629 - 32; 10.1023/A:1022355721123
- Fedina IS, Georgieva K, Grigorova I. Light-dark changes in proline content of barley leaves under salt stress. Biol Plant 2002; 45:59 - 63; 10.1023/A:1015175802847
- Fedina IS, Grigorova ID, Georgieva KM. Response of barley seedlings to UV-B radiation as affected by NaCl. J Plant Physiol 2003; 160:205 - 8; 10.1078/0176-1617-00760; PMID: 12685037
- Bramlage WJ, Leopold AC, Parrish DJ. Chilling stress to soybeans during imhibition. Plant Physiol 1978; 61:525 - 9; 10.1104/pp.61.4.525; PMID: 16660329
- Larcher W. Effects of low temperature stress and frost injury on plant productivity. In: Johnson CB, ed. Physiological processes limiting plant productivity. London: Butterworths, 1981:253-69.
- Leopold AC. Temperature effects on soybean imbibition and leakage. Plant Physiol 1980; 65:1096 - 8; 10.1104/pp.65.6.1096; PMID: 16661338
- Hare PD, Cress WA, van Staden J. A regulatory role for proline metabolism in stimulating Arabidopsis thaliana seed germination. Plant Growth Regul 2003; 39:41 - 50; 10.1023/A:1021835902351
- Fedina LS, Tsonev T, Guleva EI. The effect of pretreatment with proline on the response of Pisum sativum to salt stress. Photoynthetica 1993; 29:521 - 7
- Itai C, Paleg LG. Responses of water-stressed Hordeum distichum L. and Cucumis sativus to proline and betaine. Plant Sci Lett 1982; 25:329 - 35; 10.1016/0304-4211(82)90163-8
- Botha FC, Potgieter GP, Botha AM. Respiratory metabolism and gene expression during seed germination. Plant Growth Regul 1992; 11:211 - 24; 10.1007/BF00024560
- Shetty K. Role of proline-linked pentose phosphate pathway in biosynthesis of plant phenolics for functional food and environmental applications: a review. Process Biochem 2004; 39:789 - 804; 10.1016/S0032-9592(03)00088-8
- Posmyk MM, Janas KM. Effects of seed hydropriming in presence of exogenous proline on chilling injury limitation in Vigna radiata L. seedlings. Acta Physiol Plant 2007; 29:509 - 17; 10.1007/s11738-007-0061-2
- Webster BD, Leopold AC. The ultrastructure of dry and imbibed cotyledons of soybean. Am J Bot 1977; 64:1286 - 93; 10.2307/2442492
- Parkin KL, Kuo SJ. Chilling-induced lipid degradation in cucumber (Cucumis sativa L. cv Hybrid C) fruit. Plant Physiol 1989; 90:1049 - 56; 10.1104/pp.90.3.1049; PMID: 16666850
- van Swaaij AC, Jacobsen E, Feenstra WJ. Effect of cold hardening, wilting and exogenously applied proline on leaf proline content and frost tolerance of several genotypes of Solanum.. Physiol Plant 1985; 64:230 - 6; 10.1111/j.1399-3054.1985.tb02341.x
- Alia, Saradhi PP. Proline accumulation under heavy metal stress. J Plant Physiol 1991; 138:554 - 8; 10.1016/S0176-1617(11)80240-3
- Talanova VV, Titov AF, Boeva NP. Effect of increasing concentrations of lead and cadmium on cucumber seedlings. Biol Plant 2000; 43:441 - 4; 10.1023/A:1026735603890
- Farago ME, Mullen WA. Plants which accumulate metals. IV. A possible copper-proline complex from the roots of Armeria maritime.. Inorg Chim Acta 1979; 32:L93 - 4; 10.1016/S0020-1693(00)91627-X
- De Knecht JA, Van Dillen M, Koevoets PLM, Schat H, Verkleij JAC, Ernst WHO. Phytochelatins in cadmium-sensitive and cadmium-tolerant Silene vulgaris.. Plant Physiol 1994; 104:255 - 61; PMID: 12232077
- Xu J, Yin H, Li X. Protective effects of proline against cadmium toxicity in micropropagated hyperaccumulator, Solanum nigrum L. Plant Cell Rep 2009; 28:325 - 33; 10.1007/s00299-008-0643-5; PMID: 19043719
- Aggarwal M, Sharma S, Kaur N, Pathania D, Bhandhari K, Kaushal N, et al. Exogenous proline application reduces phytotoxic effects of selenium by minimising oxidative stress and improves growth in bean (Phaseolus vulgaris L.) seedlings. Biol Trace Elem Res 2011; 140:354 - 67; 10.1007/s12011-010-8699-9; PMID: 20455031
- Smirnoff N, Stewart GR. Nitrogen assimilation and zinc toxicity to zinc-tolerant and non-tolerant clones of Deschampsia cespitosa (L.) Beauv. New Phytol 1987; 107:671 - 80; 10.1111/j.1469-8137.1987.tb00905.x
- Sharma SS, Schat H, Vooijs R. In vitro alleviation of heavy metal-induced enzyme inhibition by proline. Phytochemistry 1998; 49:1531 - 5; 10.1016/S0031-9422(98)00282-9; PMID: 11711061
- Kastori R, Petrović M, Petrović N. Effect of excess lead, cadmium, copper, and zinc on water relations in sunflower. J Plant Nutr 1992; 15:2427 - 39; 10.1080/01904169209364485
- Wu J-T, Chang S-C, Chen K-S. Enhancement of intracellular proline level in cells of Anacystis nidulans (cyanobacteria) exposed to deleterious concentrations of copper. J Phycol 1995; 31:376 - 9; 10.1111/j.0022-3646.1995.00376.x
- Wu J-T, Hsieh M-T, Kow L-C. Role of proline accumulation in response to toxic copper in Chlorella sp. (chlorophyceae) cells. J Phycol 1998; 34:113 - 7; 10.1046/j.1529-8817.1998.340113.x
- Pedersen AL, Feldner HC, Rosendahl L. Effect of proline on nitrogenase activity in symbiosomes from root nodules of soybean (Glycine max L.) subjected to drought stress. J Exp Bot 1996; 47:1533 - 9; 10.1093/jxb/47.10.1533
- Okuma E, Soeda K, Tada M, Murata Y. Exogenous proline mitigates the inhibition of growth of Nicotiana tabacum cultured cells under saline conditions. Soil Sci Plant Nutr 2000; 46:257 - 63; 10.1080/00380768.2000.10408781
- Sharma P, Dubey RS. Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: role of osmolytes as enzyme protectant. J Plant Physiol 2005; 162:854 - 64; 10.1016/j.jplph.2004.09.011; PMID: 16146311
- Kohl DH, Kennelly EJ, Zhu Y, Schubert KR, Shearer G. Proline accumulation, nitrogenase (C2H2 reducing) activity and activities of enzymes related to proline metabolism in drought-stressed soybean nodules. J Exp Bot 1991; 42:831 - 7; 10.1093/jxb/42.7.831
- Krall JP, Edwards GE, Andreo CS. Protection of pyruvate, Pi dikinase from maize against cold lability by compatible solutes. Plant Physiol 1989; 89:280 - 5; 10.1104/pp.89.1.280; PMID: 16666527
- Paleg LG, Doughlas TJ, Van Daal A, Keech DB. Proline and betaine protect enzymes against heat inactivation. Aust J Plant Physiol 1981; 8:107 - 14
- Schobert B. Is there an osmotic regulatory mechanism in algae and higher plants?. J Theor Biol 1977; 68:17 - 26; 10.1016/0022-5193(77)90224-7; PMID: 916702
- Janas KM, Cvikrova M, Palagiewicz A, Eder J. Alterations in phenylpropanoid content in soybean roots daring low temperature acclimation. Plant Physiol Biochem 2000; 38:587 - 93; 10.1016/S0981-9428(00)00778-6
- Shetty P, Atallah MT, Shetty K. Enhancement of total phenolic, L-DOPA and proline contents in germinating faba bean (Vicia faba) in response to bacterial elicitors. Food Biotechnol 2001; 15:47 - 67; 10.1081/FBT-100103894
- Ali G, Srivastava PS, Iqbal M. Proline accumulation, protein pattern and photosynthesis in bacopa monniera regenerants grown under NaCl stress. Biol Plant 1999; 42:89 - 95; 10.1023/A:1002127711432
- Ashraf M. Breeding for salinity tolerance in plants. Crit Rev Plant Sci 1994; 13:17 - 42
- Hsu SY, Hsu YT, Kao CH. The effect of polyethylene glycol on proline accumulation in rice leaves. Biol Plant 2003; 46:73 - 8; 10.1023/A:1022362117395
- Kishor PBK, Sangam S, Amrutha RN, Laxmi PS, Naidu KR, Rao K, et al. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr Sci 2005; 88:424 - 38
- Oztürk L, Demir Y. In vivo and in vitro protective role of proline. Plant Growth Regul 2002; 38:259 - 64; 10.1023/A:1021579713832
- Rains DW, Hamlyn G. Jones TJF, Jones MB. Plant tissue and protoplast culture: applications to stress physiology and biochemistry Plants under Stress. Cambridge University Press, 1989.
- Rhodes D, Verslues PE, Sharp RE. Role of amino acids in abiotic stress resistance. In: Singh BK, ed. Plant Amino Acids: Biochemistry and Biotechnology. New York: Marcel Dekker, 1999:319-56.
- Singh TN, Aspinall D, Paleg LG. Proline accumulation and varietal adaptability to drought in barley: a potential metabolic measure of drought resistance. Nat New Biol 1972; 236:188 - 90; PMID: 4503738
- Rai VK. Role of amino acids in plant responses to stresses. Biol Plant 2002; 45:481 - 7; 10.1023/A:1022308229759
- Hanower P, Brzozowska J. Effects of osmotic stress on composition of free amino acids in cotton leaves. Phytochem 1975; 14:1691 - 4; 10.1016/0031-9422(75)85275-7
- Slukhai SI, Shvedova OE. Dynamics of free amino acid contents in maize plants in connection with soil water regimes. Fizio Biokhim kul’t Rast 1972; 4:151 - 6
- Thakur PS, Rai VK. Dynamics of amino acid accutnulation in two differentially drought resistant Zea mays cultivars in response to osmotic stress. Environ Exp Bot 1982; 22:221 - 6; 10.1016/0098-8472(82)90042-9
- Yang CW, Lin CC, Kao CH. Proline, ornithine, arginine and glutamic acid contents in detached rice leaves. Biol Plant 2000; 43:305 - 7; 10.1023/A:1002733117506
- Thakur P, Rai V. Exogenously supplied amino acids and water deficits in Zea mays cultivars. Biol Plant 1985; 27:458 - 61; 10.1007/BF02894717
- Rai VK, Sharma UD. Amino acids can modulate ABA induced stomatal closure, stomatal resistance and K+ fluxes in Vicia faba leaves. Beitr Biol Pflanzenphysiol 1991; 66:393 - 405
- Rai VK, Kumari A. Modulation of membrane permeability by amino acids in Vinca major.. Experientia (Basel) 1983; 39:301 - 3; 10.1007/BF01955316
- Rana U, Rai VK. Modulation of calcium uptake by exogenous amino acids in Phaseolus vulgaris seedlings. Acta Physiol Plant 1996; 18:117 - 20
- Santos MA, Camara R, Rodriguez P, Glaparols I, Torne JM. Influence of exogenous maize callus subjects to salt stress. Plant Cell Tissue Organ Cult 1996; 47:59 - 65; 10.1007/BF02318966
- Palfi G, Koves F, Bito M, Sebestyen R. The role of amino acids during water stress in species accumulating proline. Phyton Intl J Exp Bot 1974; 32:121 - 7
- Sánchez E, Garcia PC, López-Lefebre LR, Rivero RM, Ruiz JM, Romero L. Proline metabolism in response to nitrogen deficiency in French Bean plants (Phaseolus vulgaris L. cv Strike). Plant Growth Regul 2002; 36:261 - 5; 10.1023/A:1016583430792
- Heuer B. Influence of exogenous application of proline and glycinebetaine on growth of salt-stressed tomato plants. Plant Sci 2003; 165:693 - 9; 10.1016/S0168-9452(03)00222-X
- Hare PD, Cress WA, van Staden J. The effects of exogenous proline and proline analogues on in vitro shoot organogenesis in Arabidopsis.. Plant Growth Regul 2001; 34:203 - 7; 10.1023/A:1013326526875
- Hare PD. A regulatory role for proline metabolism in Arabidopsis thaliana (L) Heynth. Pietermaritzburg, South Africa: University of Natal, 1998.
- García-Ríos M, Fujita T, LaRosa PC, Locy RD, Clithero JM, Bressan RA, et al. Cloning of a polycistronic cDNA from tomato encoding γ-glutamyl kinase and γ-glutamyl phosphate reductase. Proc Natl Acad Sci U S A 1997; 94:8249 - 54; 10.1073/pnas.94.15.8249; PMID: 9223347
- Zhang CS, Lu Q, Verma DP. Removal of feedback inhibition of delta 1-pyrroline-5-carboxylate synthetase, a bifunctional enzyme catalyzing the first two steps of proline biosynthesis in plants. J Biol Chem 1995; 270:20491 - 6; 10.1074/jbc.270.35.20491; PMID: 7657626
- Rodriguez MM, Heyser JW. Growth inhibition by exogenous proline and its metabolism in saltgrass (Distichlis spicata) suspension cultures. Plant Cell Rep 1988; 7:305 - 8; 10.1007/BF00269924
- Chen SL, Kao CH. Cadmium induced changes in proline level and peroxidase activity in roots of rice seedlings. Plant Growth Regul 1995; 17:67 - 71
- Nanjo T, Fujita M, Seki M, Kato T, Tabata S, Shinozaki K. Toxicity of free proline revealed in an arabidopsis T-DNA-tagged mutant deficient in proline dehydrogenase. Plant Cell Physiol 2003; 44:541 - 8; 10.1093/pcp/pcg066; PMID: 12773641