
Abstract
In bacteria, transcription, translation and gene regulation are highly coupled processes. The achievement of a certain functional structure at a distinct temporal and spatial position is therefore essential for RNA molecules. Proteins that facilitate this proper folding of RNA molecules are called RNA chaperones. Here a prominent example from E. coli is reviewed: the nucleoid associated protein StpA. Based on its various RNA remodeling functions, we propose a mechanistic model that explains how StpA promotes RNA folding. Through transient interactions via the RNA backbone, thereby shielding repelling charges in RNA, it pre-positions the RNA molecules for the successful formation of transition states from encounter complexes.
Acknowledgements
We would like to thank all members of the Schroeder Lab for helpful discussions on the topic of RNA chaperones. We are indebted to Jennifer L. Boots and Bob Zimmermann for critical reading of the manuscript and helpful comments. This work is supported by F.W.F. through a Lise Meitner-Position (M1157-B12) to B.F. and grant F1703 to R.S. and by the European Community (EU-NMR, Contract # RII3-026145). M.D. is funded by the University of Vienna.
Figures and Tables
Figure 1 Domain architecture of StpA in comparison to H-NS: (A) displays the primary structure of both proteins; in a color-coded scheme the distribution of charged amino-acids is displayed, blue and red bars indicate positively (R and K) and negatively (D and E) charged amino-acids, respectively; the green letters separate every ten amino-acids; the over-bars indicate the domain structure of StpA as discussed in the text and the grey under-bars show the domains of H-NS for which high-resolution structures (pdb-codes: 1ni8 and 1 hns) are available; residues involved in H-NS DNA binding are highlighted by arrow heads; in (B) the three-dimensional structures are displayed in cartoon representation; the color-coding on the surface representation equates the electrostatic potential; residues involved in H-NS DNA binding are highlighted by stick representation.
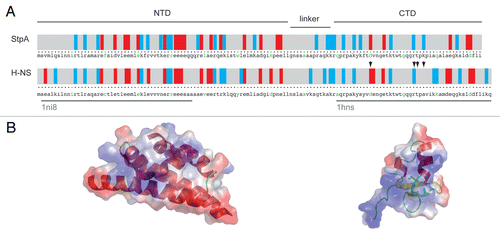
Figure 2 Two simplistic models of how proteins facilitate the remodeling of RNA conformations. (A) Energy landscape for a tight binding protein that acts as a co-factor for RNA and through binding induces a conformational change that is more stable (red) then the respective RNA conformation in the absence of the protein (blue). (B) As an experimental example for such a scenario the changes in the td intron structure upon binding of Cyt-18 are shown. Without the protein residues, A46 and A47 are moderately accessible to DMS modifications (blue line). In the presence of Cyt-18 these residues become involved in a stable tertiary interaction and are protected (red line). No apparent changes are monitored for A48 which is part of a secondary structure element. (C) Energy landscape for proteins that interact only transiently with the transition state of an RNA and thereby lowers the activation energy in the remodeling reaction (blue versus red reaction path, for RNA alone and RNA/protein, respectively). (D) As an experimental example for this scenario the changes in the td intron structure upon interaction with StpA are shown. Without the protein residues A46 and A47 are moderately accessible to DMS modifications. In the presence of StpA these residues sample more open conformations and are easier to access by DMS (red lines). No apparent changes are monitored for A48 as it is involved in a secondary structure interaction; (B and C) are adapted from reference (Waldsich et al.).Citation13
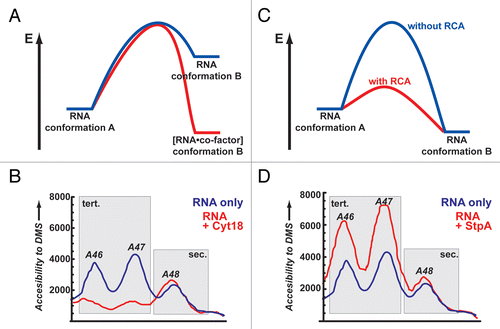
Figure 3 A Generalized Model for StpA Activities. (A) Two annealing RNAs (R1 and R2 with complementary sequences) run through different states before they form a duplex. In addition to the most stable double-strand, alternative duplexes (indicated by the subfix ‘alt’) can form and, depending on their thermodynamic stability, eventually fall apart again. (B) Proposed mechanisms for StpA-facilitated RNA-RNA annealing and strand displacement. Partial opening of the R1R2 duplex (indicated through parentheses) allows the R1-complementary R3 RNA to invade the double-strand.
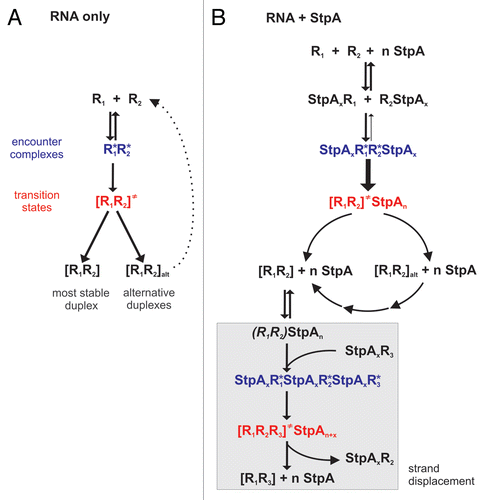
Table 1 The table summarizes the results of various in vitro chaperone assays for WT StpA, the N terminal domain (NTD) and the C terminal domain (CTD) of StpA as well as two StpA mutants G126V and L30P