
Abstract
Earlier studies have shown adjudin to cause aspermatogenesis by depleting virtually all germ cells from the seminiferous epithelium, leading to transient infertility; spermatogenesis and fertility were re-established several weeks later after germ cell differentiation was reinitiated by spermatogonia. While adjudin is known to exert its initial effects at the apical ectoplasmic specialization (a testis-specific atypical anchoring junction), thereby perturbing spermatid adhesion, its molecular target at this site is not known. Herein, we report the production of a specific antibody against adjudin after this compound was conjugated to an adjuvant (i.e., keyhole limpet hemocyanin) to maximize immune response in rabbits. This antibody was utilized for co-immunoprecipitation by using an affinity resin to pull-down the binding partners of adjudin. Using this approach coupled with mass spectrometry and immunoblotting, we show testin (a protein largely restricted to the apical ES in the adult testis) and actin-myosin to be molecular targets of adjudin. These findings provide a platform for future functional studies, not only to better understand the molecular mechanism behind adjudin-induced germ cell loss from the seminiferous epithelium, but also to understand the molecular basis of spermiation.
Acknowledgments
This work was supported in part by grants from the National Institutes of Health (NICHD, R01 HD056034 and R01 HD056034-02-S1 to C.Y.C.; U54 HD029990 Project 5 to C.Y.C.; R03 HD061401 to D.D.M.).
Figures and Tables
Figure 1 Use of Sertoli and germ cell co-cultures to assess the effects of adjudin on disrupting germ cell adhesion in vitro. (A) This is a typical Sertoli cell epithelium in which cells were cultured for 4 days. A functional Sertoli cell barrier, manifested by the presence of TJs (see blue arrowheads) and basal ES, is visible between adjacent Sertoli cells. The basal ES is typified by the presence of actin filament bundles (see green arrowheads) sandwiched in between cisternae of endoplasmic reticulum (ER) and apposing Sertoli cell plasma membranes (see orange arrowheads). Desmosomes, typified by the presence of electron-dense material, are also visible (see red arrowheads). (B) A germ cell attaches to the Sertoli cell epithelium ∼48 hours after seeding germ cells (Nu, Sertoli cell nucleus). (C) This is a step 9–10 spermatid with a functional apical ES, typified by the presence of actin filament bundles (see green arrowheads) sandwiched in between cisternae of ER and apposing Sertoli cell-spermatid plasma membranes (see orange arrowheads). Both the acrosome (Ac) and condensed general material within the nucleus (Nu) are visible. Microvilli (A–C) are also seen in all micrographs (see black arrows). (D) Germ cells, such as those shown in (B and C) were fluorescently labeled and added onto the Sertoli cell epithelium (A) at time 0. After Sertoli-germ cell adhesion was established, adjudin (1 µg/ml) was added into triplicate wells per time point to assess its effects on germ cell adhesion (see Materials and Methods). A loss in germ cell adhesion was detected 1 hour (hr) after treatment when the amount of fluorescence remaining in co-cultures was quantified by cytofluorometry. Error bars represent mean ± SD from at least three different experiments. *p < 0.05; ns, not significant. (two-way ANOVA followed by Tukey's post-hoc test). Bar in A = 0.5 µm, B = 4 µm, C = 0.3 µm.
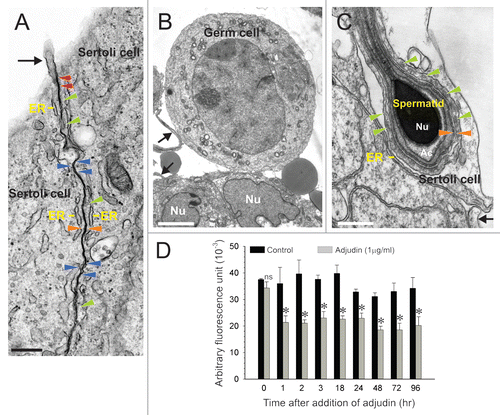
Figure 2 Conjugation of adjudin to KLH (keyhole limpet hemocyanin, an adjuvant), and its use as an immunogen for anti-adjudin antibody production. KLH was first reacted with SFB (succinimidyl 4-formylbenzoate, a heterobifunctional crosslinker) to generate a benzaldehyde at its N-terminus. This modified KLH was then reacted spontaneously in physiological buffer with the hydrazide group in adjudin, forming a stable KLH-adjudin conjugate via a hydrazone linkage.
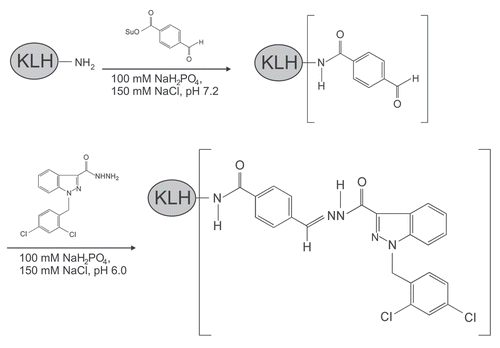
Figure 3 Specificity analysis of anti-adjudin antiserum. Structural formulae of adjudin (A) and lonidamine (B). (C) Displacement of [3H]-adjudin binding to anti-adjudin anti-serum by unlabelled (i.e., cold) adjudin, lonidamine, testosterone or estradiol-17β. Adjudin competed with the binding of [3H]-adjudin to its antibody. While lonidamine shares structural similarities with adjudin, it generated a parallel but incomplete displacement curve. Both testosterone and estradiol-17β failed to compete with the binding of [3H]-adjudin to its antibody.
![Figure 3 Specificity analysis of anti-adjudin antiserum. Structural formulae of adjudin (A) and lonidamine (B). (C) Displacement of [3H]-adjudin binding to anti-adjudin anti-serum by unlabelled (i.e., cold) adjudin, lonidamine, testosterone or estradiol-17β. Adjudin competed with the binding of [3H]-adjudin to its antibody. While lonidamine shares structural similarities with adjudin, it generated a parallel but incomplete displacement curve. Both testosterone and estradiol-17β failed to compete with the binding of [3H]-adjudin to its antibody.](/cms/asset/b2487503-9805-449f-83a3-26f148ea5f42/kspe_a_10916449_f0003.gif)
Figure 4 A study to assess the ability of the anti-adjudin antibody to protect adjudin-induced germ cell loss from the seminiferous epithelium in rat testes. (A) Adjudin competed with the binding of [3H]-adjudin to its antibody in a competitive binding assay. (B) Anti-adjudin IgG blocked the effects of adjudin on Sertoli-germ cell adhesion. (i) Cross-section of the control testis. (ii) Cross-section of the testis 21 days after treatment with adjudin (50 mg/kg b.w.). (iii) Cross-section of the testis 21 days after treatment with non-immune rabbit IgG and adjudin (50 mg/kg b.w.). (iv) Cross-section of the testis 21 days after treatment with anti-adjudin IgG and adjudin. All cross-sections were stained with H&E. Boxed areas in (i–iv) represent magnified views, and these are shown to the right of each image. Bar in (Bi) [also applies to (Bii–iv)] = 125 µm; bar in magnified views = 75 µm. Error bars represent mean ± SD from four different experiments. *p < 0.05. (two-way ANOVA followed by Tukey's post-hoc test).
![Figure 4 A study to assess the ability of the anti-adjudin antibody to protect adjudin-induced germ cell loss from the seminiferous epithelium in rat testes. (A) Adjudin competed with the binding of [3H]-adjudin to its antibody in a competitive binding assay. (B) Anti-adjudin IgG blocked the effects of adjudin on Sertoli-germ cell adhesion. (i) Cross-section of the control testis. (ii) Cross-section of the testis 21 days after treatment with adjudin (50 mg/kg b.w.). (iii) Cross-section of the testis 21 days after treatment with non-immune rabbit IgG and adjudin (50 mg/kg b.w.). (iv) Cross-section of the testis 21 days after treatment with anti-adjudin IgG and adjudin. All cross-sections were stained with H&E. Boxed areas in (i–iv) represent magnified views, and these are shown to the right of each image. Bar in (Bi) [also applies to (Bii–iv)] = 125 µm; bar in magnified views = 75 µm. Error bars represent mean ± SD from four different experiments. *p < 0.05. (two-way ANOVA followed by Tukey's post-hoc test).](/cms/asset/a2e087da-7e32-4b58-b9ef-bbaf579a4237/kspe_a_10916449_f0004.gif)
Figure 5 Identification of actin and myosin as molecular targets of adjudin in the rat testis by co-immunoprecipitation and mass spectrometry. (A) Actin and myosin were found to bind adjudin as determined by co-immunoprecipitation using affinity chromatography with an anti-adjudin IgG-Protein A resin, coupled with SDS-PAGE, Coomassie blue gel staining and mass spectrometry. M, molecular weight marker. (B) Sequences corresponding to actin (i) and myosin (ii) as obtained by mass spectrometry.
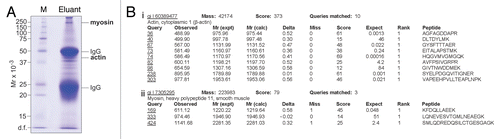
Figure 6 Characterization of actin as a molecular target of adjudin. (A) Adjudin binds to actin (42 kDa) as determined by co-immunoprecipitation. Aliquots (lane 1, 10 µl; lane 2, 30 µl) of the eluant () were resolved by SDS-PAGE and proteins were transferred to nitrocellulose for immunoblotting using anti-actin IgG. Ctrl, testis lysate (20 µg). (B) The interaction of adjudin with actin was specific since unlabeled (i.e., cold) adjudin displaced the binding of [3H]-adjudin to actin IgG in a competitive binding assay. Error bars represent mean ± SD from three different experiments. *p < 0.05; ns, not significant. (Student's t-test).
![Figure 6 Characterization of actin as a molecular target of adjudin. (A) Adjudin binds to actin (42 kDa) as determined by co-immunoprecipitation. Aliquots (lane 1, 10 µl; lane 2, 30 µl) of the eluant (Fig. 5A) were resolved by SDS-PAGE and proteins were transferred to nitrocellulose for immunoblotting using anti-actin IgG. Ctrl, testis lysate (20 µg). (B) The interaction of adjudin with actin was specific since unlabeled (i.e., cold) adjudin displaced the binding of [3H]-adjudin to actin IgG in a competitive binding assay. Error bars represent mean ± SD from three different experiments. *p < 0.05; ns, not significant. (Student's t-test).](/cms/asset/9cf8bd90-fb49-4bdd-b1f8-f6e734218702/kspe_a_10916449_f0006.gif)
Figure 7 Characterization of testin as a molecular target of adjudin. (A) Adjudin binds to testin (35 and 37 kDa) as determined by co-immunoprecipitation. Aliquots (lane 1, 10 µl; lane 2, 30 µl) of the eluant () were resolved by SDS-PAGE and proteins were transferred to nitrocellulose for immunoblotting using a monospecific anti-testin antibody.Citation22,Citation23 Ctrl, testis lysate (100 µg). (B) The interaction of adjudin with testin was specific since unlabeled (i.e., cold) adjudin displaced the binding of [3H]-adjudin to testin IgG in a competitive binding assay. Error bars represent mean ± SD from three different experiments. *p < 0.05. (Student's t-test).
![Figure 7 Characterization of testin as a molecular target of adjudin. (A) Adjudin binds to testin (35 and 37 kDa) as determined by co-immunoprecipitation. Aliquots (lane 1, 10 µl; lane 2, 30 µl) of the eluant (Fig. 5A) were resolved by SDS-PAGE and proteins were transferred to nitrocellulose for immunoblotting using a monospecific anti-testin antibody.Citation22,Citation23 Ctrl, testis lysate (100 µg). (B) The interaction of adjudin with testin was specific since unlabeled (i.e., cold) adjudin displaced the binding of [3H]-adjudin to testin IgG in a competitive binding assay. Error bars represent mean ± SD from three different experiments. *p < 0.05. (Student's t-test).](/cms/asset/7850e355-e0d9-40e6-bbaf-593b3f6b09a2/kspe_a_10916449_f0007.gif)