
Abstract
Previous investigations demonstrated that optimization of murine immunological reactivity in tissue culture required a sulfhydryl compound; the most effective being 2-mercaptoethanol (2-Me). Since these reports, 2-Me was found beneficial for both growth/function of other cell-types in vitro, including those of other species, and when fed orally, it impeded and/or reversed some in situ physiological changes associated with aging. More recently, thiol-containing compounds possessing oxidation-reduction potentials weaker than 2-Me were found to impart beneficial effects for many other, including human, diseases. Based on these effects, the research herein addressed the question: What consequences might dietary 2-Me impart on health and disease of mice other than those associated with aging? The main parameters monitored over the lifetime of individual animals exposed to dietary 10-3 M 2-Me in their drinking water were: quality of life (obesity and development of recumbent, emaciated and/or cachectic health); longevity; and appearance of tumors. Instead of anticipated toxic attributes, the following unique benefits were found; mean survival of a moderately-lived strain (A/J) was increased 40.8%, high-fat-diet obesity was curtailed in C57BL/10 mice, and a goal of aging intervention protocols, namely preventing loss of quality of life during aging (recumbent, emaciated and/or cachectic) was achieved. Various mechanisms are discussed as they pertain to these findings.
Acknowledgements
A special thanks to Annette Adelmann for technical assistance and to Dr. Ellen Heber-Katz for editorial assistance. This research was funded, in part, by NIH Grants R01CA023678 and R01AI019643.
Ethics Statement
There was no part of the research that inflicted pain or suffering, except that which occurred naturally because of aging. Euthanasia was used to ameliorate suffering of old, incapacitated mice.
Figures and Tables
Figure 1 Survival of nontreated and 2-Me-treated C57BL/10 mice fed different diets. (A) Diet was 10%T, (B) diet was 6%T, (C) diet was 4%P, (D) diet was 10%W. Data for not treated (n = 14) shown in black and for treated (n = 14) in red. p values of treated vs. not treated for 10%T was 0.13, for 6%T was 0.17, for 4%P was 0.099, and for 10%W was 0.24.
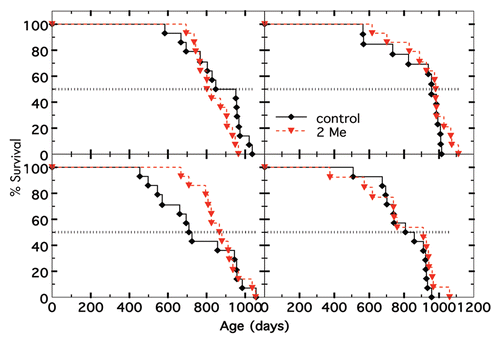
Figure 2 Mean body weight changes ±SE of mice on different diets plus/minus 2-Me. (A) Mice (n = 14) not treated with 2-Me. Each diet is indicated by a different color. (B) Mice (n = 14) treated with 2-Me. Each diet is indicated by same color as in (A). p values for treated vs. non-treated on a given diet were all <0.01.
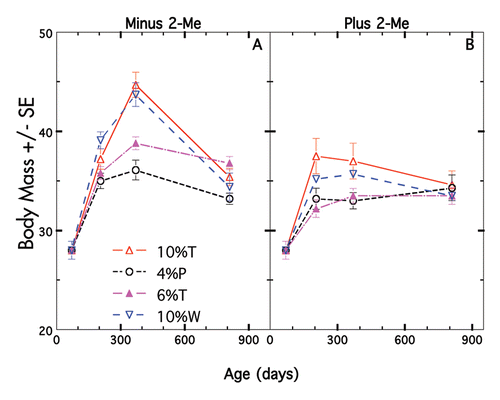
Figure 3 Body weight changes of individual mice on two different diets ± 2-Me. (A) Diet was 10%T minus 2-Me, (B) diet was 4%P minus 2-Me, (C) diet was 10%T plus 2-Me, (D) diet was 4%P plus 2-Me. Individual mice depicted by different colors.
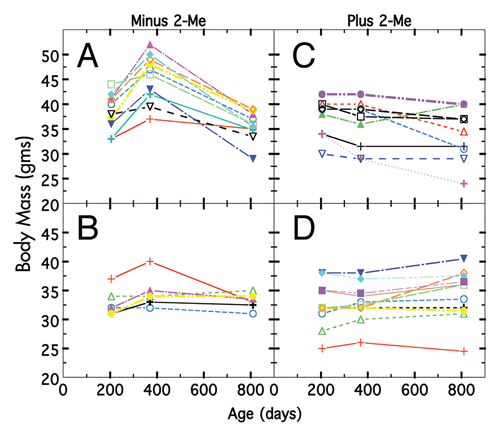
Figure 4 Cholesterol levels on four diets ±2-Me. Mean ± SE M serum cholesterol of individual nonfasted animals. p values for 4%P was 0.005 (n = 15 t, n = 6 nt), for 10%T was 0.010 (n = 9 t, n = 8 nt), for 6%T was not calculable (n = 6 t, n = 4 nt) and for 10%W was 0.029 (n = 9 t, n = 6 nt).
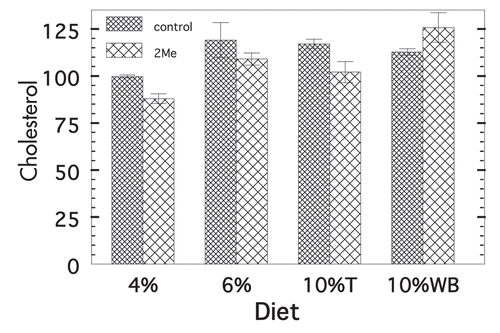
Figure 5 Survival of 2-Me treated and nontreated A/J and B10.A(4R) mice. Data for not treated shown in black and for 2-Me treated in red. For A/J, n = 7 for not treated and 9 for treated. For 4R mice, n = 22 for both treated and not treated.
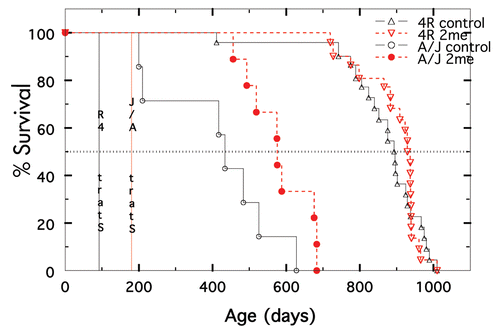
Table 1 Average daily food, water and 2-Me consumed/mouse