Abstract
High-mobility group box 1 (HMGB1) was originally identified as a highly conserved nuclear DNA-binding protein that participates in DNA replication, repair and transcriptional regulation of gene expression. Although the nuclear role of HMGB1 is not quite understood, recent studies characterized the emerging role of extracellular HMGB1 as a prototypical danger signal that regulates inflammatory and repair responses. Under conditions of infection, injury and sterile inflammation, HMGB1 can be passively released from damaged cells or actively secreted from activated immune cells. Inflammasomes, large caspase-1-activating protein complexes, were recently shown to play a critical role in mediating the extracellular release of HMGB1 from activated and infected immune cells.
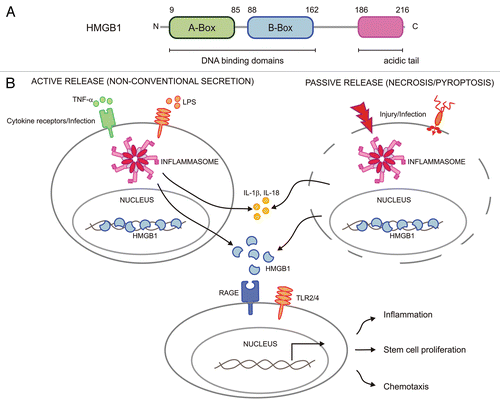
High-mobility group box 1 (HMGB1, also known as amphoterin) was identified originally as a highly conserved non-histone DNA-binding factor that is expressed by virtually all nucleated eukaryotic cells.Citation1 Structurally, HMGB1 is a 30 kDa protein that is composed of two homologous DNA-binding motifs (called ‘A’ and ‘B’ boxes) followed by a negatively charged acidic tail in its carboxyl-terminus ().Citation2,Citation3 Although the nuclear role of HMGB1 is incompletely understood, the protein has been implicated in bending DNA to facilitate gene transcription, DNA replication and repair.Citation4–Citation7 The importance of HMGB1 in regulating these nuclear processes is emphasized by the phenotype of HMGB1-deficient mice, which die shortly after birth because of severe energy deficits and hypoglycemia, possibly as a result of impaired glucocorticoid receptor signaling.Citation8
In addition to its roles in regulating nucleosome function and transcription, HMGB1 more recently emerged as an extracellularly released mediator of inflammation and repair responses in lipopolysaccharide (LPS)-induced endotoxemia and sepsis.Citation9 Passive immunization with HMGB1-neutralizing antibodies prevented organ damage in animal endotoxemia and sepsis models.Citation9–Citation12 Moreover, increased HMGB1 serum levels were confirmed in septic patients.Citation13 Subsequent studies implicated HMGB1 in the induction of systemic inflammation following trauma and ischemia/reperfusion injury in the liver, heart, kidney and brain.Citation14–Citation17 Elevated HMGB1 levels in circulation were also associated with lung injury, myocardial infarction and rheumatoid arthritis.Citation18–Citation21 HMGB1 induces the recruitment of inflammatory cells, contributes to dendritic cell maturation and to proliferation of activated T cells.Citation22,Citation23 Thus, extracellular HMGB1 appears to be associated with inflammatory and repair responses during both infectious and (sterile) trauma and autoimmune disorders. Therefore, HMGB1 is widely considered to be a danger-associated molecular pattern (DAMP) that—similar to interleukin-33 and in agreement with Matzinger's danger model—engages immune receptors upon its release in the extracellular milieu.Citation24,Citation25 Indeed, recombinant HMGB1 produced in Escherichia coli is a potent inducer of pro-inflammatory cytokines when administered to cells or injected in mice.Citation9,Citation26–Citation30 HMGB1-induced activation of immune receptors may involve the assembly of immunostimulatory complexes with endogenous or microbial co-factors. Indeed, the receptor for advanced glycation endproducts (RAGE) was potently activated by HMGB1/CpG DNA complexes, whereas highly purified HMGB1 has only weak pro-inflammatory activity.Citation30,Citation31 In addition to RAGE, HMGB1 was suggested to induce secretion of pro-inflammatory cytokines and chemokines by inducing NF-κB signaling downstream of Toll-like receptor (TLR)2 and TLR4 activation.Citation32–Citation34 Thus, extracellular HMGB1 might bind microbial components and/or endogenous substances that are released from injured tissue to induce or augment the production of inflammatory mediators during infections and in response to (sterile) trauma.
An intriguing question that has attracted considerable attention is how HMGB1 travels from the nucleus to reach the extracellular milieu given that it lacks a classical secretion signal. Such signal peptides are usually required for transporting secretory proteins through the endoplasmic reticulum (ER) lumen to the extracellular space in Golgi-derived secretory vesicles.Citation35,Citation36 However, certain cytoplasmic and nuclear proteins that lack classical secretion signal peptides can reach the extracellular milieu through ER- and Golgi-independent pathways.Citation37 Indeed, HMGB1 was shown to be released from LPS-activated monocytes and macrophage cell lines consistent with its role as a prototypical alarmin.Citation38 Another protein thought to undergo unconventional protein secretion is the pro-inflammatory cytokine IL-1β.Citation39 IL-1β and the related cytokine IL-18 are both produced as inactive cytosolic precursor proteins that are only released following their maturation by the cysteine protease caspase-1. Caspase-1 itself requires activation by large protein complexes termed ‘inflammasomes,’ which are assembled on NOD-like receptor (NLR) and HIN-200 protein scaffolds.Citation40 NLRs contain a centrally located NACHT domain, whereas HIN-200 proteins such as the interferon-inducible member absent in melanoma 2 (AIM2) are characterized by the presence of a carboxy-terminal DNA-binding HIN domain.Citation41,Citation42 Inflammasomes activate signaling pathways that contribute to immune responses and host defense mechanisms when they (directly or indirectly) sense the presence of particular microbial pathogens and DAMPs in the host cytosol or in intracellular compartments.Citation43 For instance, the NLRC4 inflammasome is responsible for caspase-1 activation in macrophages infected with the bacterial pathogens Salmonella typhimurium, Pseudomonas aeruginosa, Legionella pneumophila and Shigella flexneri.Citation43 On the other hand, the NLRP3 inflammasome induces caspase-1 activation upon infection with Staphylococcus aureus and Klebsiella pneumonia.Citation44–Citation46 The NLRP3 inflammasome also induces secretion of IL-1β and IL-18 from LPS-primed macrophages that are subsequently treated with endogenous alarmins such as ATP and uric acid, or exposed to microbial toxins such as nigericin.Citation43 Intriguingly, NLRP3 inflammasome activation was recently shown to be essential not only for IL-1b secretion, but also for HMGB1 release from LPS-primed macrophages that are treated with ATP or exposed to nigericin.Citation10 Similarly, extracellular release of HMGB1 from S. typhimurium-infected macrophages relied on activation of caspase-1 by the NLRC4 inflammasome.Citation10 Unlike IL-1β and IL-18, HMGB1 did not undergo caspase-1-mediated processing. Nevertheless, the enzymatic activity of caspase-1 is required for secretion of IL-1β, IL-18, HMGB1 and growth factors such as fibroblast growth factor 2.Citation10,Citation47 The observation that HMGB1 release proceeded unabated in macrophages lacking known caspase-1-substrates (IL-1β, IL-18 and caspase-7) suggests that caspase-1 may induce non-canonical secretion of HMGB1 and other proteins through proteolytic activation of an unknown secretion apparatus (). In this context, the small GTPase Rab39a was recently characterized as a caspase-1 substrate that has been implicated in secretion of IL-1β from LPS-activated THP-1 cells.Citation48 Moreover, several mutually non-exclusive models have been proposed to explain the release of IL-1β, IL-18 and DAMPS such as HMGB1 in shedded microvesicles, secretory lysosomes or exosomes.Citation37
In addition to mediating the non-conventional secretion of pro-inflammatory cytokines, DAMPs and growth factors, inflammasome activation in activated immune cells triggers the induction of a specialized caspase-1-mediated cell death program called pyroptosis.Citation49,Citation50 Pyroptosis is generally regarded as a pro-inflammatory cell death mode because it is associated with osmotic lysis and the release of the intracellular content into the extracellular milieu (). Concurrently, reduced serum HMGB1 levels in caspase-1-deficient mice were shown to correlate with their resistance to LPS, and HMGB1 neutralizing antibodies protected IL-1β/IL-18-deficient mice from LPS-induced endotoxemia.Citation10 In this regard, pyroptosis may resemble necrosis, an inherently proinflammatory and pathological cell death mode that results in extensive tissue damage.Citation49 Indeed, HMGB1 has been identified as a critical danger signal contributing to necrosis-associated inflammation.Citation38 This is illustrated by the observation that necrotic cell debris from HMGB1-deficient cells was significantly impaired in inducing pro-inflammatory cytokines.Citation38,Citation51 It is thus tempting to speculate that cellular injury in response to trauma and microbial infections may trigger pyroptosis and necrotic cell death in order to induce inflammatory and repair responses through the extracellular release of DAMPs such as HMGB1 and S100 proteins ().Citation49
In conclusion, inflammasomes may release HMGB1 in circulation through unconventional protein secretion, or as a consequence of cell lysis during pyroptosis. Depending on the pathological context, these mechanisms might contribute differentially to the extracellular release of HMGB1 and the induction of HMGB1-mediated immune responses. Further characterization of the molecular mechanisms of pyroptosis and unconventional protein secretion may shed light on the intricacies of inflammasome-mediated release of danger signals and inflammatory cytokines.
Abbreviations
| DAMP | = | danger-associated molecular pattern |
| ER | = | endoplasmic reticulum |
| HMGB1 | = | high-mobility group box 1 |
| IL | = | interleukin |
| LPS | = | lipopolysaccharide |
| NLR | = | NOD-like receptor |
| NOD | = | nucleotide-binding and oligomerization domain |
| RAGE | = | receptor for advanced glycation endproducts |
| TLR | = | Toll-like receptor |
Figures and Tables
Figure 1 Structure of HMGB1 and its functions in the extracellular environment. (A) Human HMGB1 is a protein of 216 amino acids that is composed of two homologous DN A-binding domains called ‘A and B boxes’ that are followed by a negatively charged acidic tail in the carboxyl-terminus. (B) HMGB1 can be secreted through non-canonical mechanisms along with pro-inflammatory cytokines such as IL-1β and IL-18 following inflammasome activation in stimulated and/or infected immune cells, or it may be passively released from damaged and infected cells undergoing necrotic or pyroptotic cell death. Extracellular HMGB1 can bind to its receptors RAGE and TLR2/4 on effector cells in order to induce inflammation, chemotaxis and repair responses.
Acknowledgements
This work was supported by National Institute of Health Grant AR056296, a Cancer Center Support Grant (CCSG 2 P30 CA 21765), Centers of Excellence for Influenza Research and Surveillance (CEIRS) project and the American Lebanese Syrian Associated Charities (ALSAC) to T-D.K. LVW and ML are supported by the Fonds voor Wetenschappelijk Onderzoek-Vlaanderen, and by the European Union Framework Program 7 Marie-Curie grant 256432 to ML.
Addendum to:
References
- Goodwin GH, Sanders C, Johns EW. A new group of chromatin-associated proteins with a high content of acidic and basic amino acids. Eur J Biochem 1973; 38:14 - 19
- Hardman CH, Broadhurst RW, Raine AR, Grasser KD, Thomas JO, Laue ED. Structure of the A-domain of HMG1 and its interaction with DNA as studied by heteronuclear three- and four-dimensional NMR spectroscopy. Biochemistry 1995; 34:16596 - 16607
- Weir HM, Kraulis PJ, Hill CS, Raine AR, Laue ED, Thomas JO. Structure of the HMG box motif in the B-domain of HMG1. EMBO J 1993; 12:1311 - 1319
- Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nature Rev Immunol 2005; 5:331 - 342
- Prendergast P, Onate SA, Christensen K, Edwards DP. Nuclear accessory factors enhance the binding of progesterone receptor to specific target DNA. J Steroid Biochem Mol Biol 1994; 48:1 - 13
- Zhang CC, Krieg S, Shapiro DJ. HMG-1 stimulates estrogen response element binding by estrogen receptor from stably transfected HeLa cells. Mol Endocrinol 1999; 13:632 - 643
- Bustin M. Regulation of DNA-dependent activities by the functional motifs of the high-mobility-group chromosomal proteins. Mol Cell Biol 1999; 19:5237 - 5246
- Calogero S, Grassi F, Aguzzi A, Voigtlander T, Ferrier P, Ferrari S, et al. The lack of chromosomal protein Hmg1 does not disrupt cell growth but causes lethal hypoglycaemia in newborn mice. Nat Genet 1999; 22:276 - 280
- Wang H, Bloom O, Zhang M, Vishnubhakat JM, Ombrellino M, Che J, et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science 1999; 285:248 - 251
- Lamkanfi M, Sarkar A, Vande Walle L, Vitari AC, Amer AO, Wewers MD, et al. Inflammasome-dependent release of the alarmin HMGB1 in endotoxemia. J Immunol 2010; 185:4385 - 4392
- Yang H, Ochani M, Li J, Qiang X, Tanovic M, Harris HE, et al. Reversing established sepsis with antagonists of endogenous high-mobility group box 1. Proc Natl Acad Sci USA 2004; 101:296 - 301
- Suda K, Kitagawa Y, Ozawa S, Saikawa Y, Ueda M, Ebina M, et al. Anti-high-mobility group box chromosomal protein 1 antibodies improve survival of rats with sepsis. World J Surg 2006; 30:1755 - 1762
- Sunden-Cullberg J, Norrby-Teglund A, Rouhiainen A, Rauvala H, Herman G, Tracey KJ, et al. Persistent elevation of high mobility group box-1 protein (HMGB1) in patients with severe sepsis and septic shock. Crit Care Med 2005; 33:564 - 573
- Levy RM, Mollen KP, Prince JM, Kaczorowski DJ, Vallabhaneni R, Liu S, et al. Systemic inflammation and remote organ injury following trauma require HMGB1. Am J Physiol Regul Integr Comp Physiol 2007; 293:R1538 - R1544
- Tsung A, Sahai R, Tanaka H, Nakao A, Fink MP, Lotze MT, et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J Exp Med 2005; 201:1135 - 1143
- Goldstein RS, Gallowitsch-Puerta M, Yang L, Rosas-Ballina M, Huston JM, Czura CJ, et al. Elevated high-mobility group box 1 levels in patients with cerebral and myocardial ischemia. Shock 2006; 25:571 - 574
- Faraco G, Fossati S, Bianchi ME, Patrone M, Pedrazzi M, Sparatore B, et al. High mobility group box 1 protein is released by neural cells upon different stresses and worsens ischemic neurodegeneration in vitro and in vivo. J Neurochem 2007; 103:590 - 603
- Abraham E, Arcaroli J, Carmody A, Wang H, Tracey KJ. HMG-1 as a mediator of acute lung inflammation. J Immunol 2000; 165:2950 - 2954
- Kohno T, Anzai T, Naito K, Miyasho T, Okamoto M, Yokota H, et al. Role of high-mobility group box 1 protein in post-infarction healing process and left ventricular remodelling. Cardiovasc Res 2009; 81:565 - 573
- Kokkola R, Sundberg E, Ulfgren AK, Palmblad K, Li J, Wang H, et al. High mobility group box chromosomal protein 1: a novel proinflammatory mediator in synovitis. Arthritis Rheum 2002; 46:2598 - 2603
- Taniguchi N, Kawahara K, Yone K, Hashiguchi T, Yamakuchi M, Goto M, et al. High mobility group box chromosomal protein 1 plays a role in the pathogenesis of rheumatoid arthritis as a novel cytokine. Arthritis Rheum 2003; 48:971 - 981
- Sundberg E, Fasth AE, Palmblad K, Harris HE, Andersson U. High mobility group box chromosomal protein 1 acts as a proliferation signal for activated T lymphocytes. Immunobiology 2009; 214:303 - 309
- Blanco P, Palucka AK, Pascual V, Banchereau J. Dendritic cells and cytokines in human inflammatory and autoimmune diseases. Cytokine Growth Factor Rev 2008; 19:41 - 52
- Matzinger P. Tolerance, danger, and the extended family. Annu Rev Immunol 1994; 12:991 - 1045
- Lamkanfi M, Dixit VM. IL-33 raises alarm. Immunity 2009; 31:5 - 7
- Kokkola R, Andersson A, Mullins G, Ostberg T, Treutiger CJ, Arnold B, et al. RAGE is the major receptor for the proinflammatory activity of HMGB1 in rodent macrophages. Scand J Immunol 2005; 61:1 - 9
- Dumitriu IE, Baruah P, Valentinis B, Voll RE, Herrmann M, Nawroth PP, et al. Release of high mobility group box 1 by dendritic cells controls T cell activation via the receptor for advanced glycation end products. J Immunol 2005; 174:7506 - 7515
- Wang H, Yang H, Tracey KJ. Extracellular role of HMGB1 in inflammation and sepsis. J Intern Med 2004; 255:320 - 331
- Li J, Wang H, Mason JM, Levine J, Yu M, Ulloa L, et al. Recombinant HMGB1 with cytokine-stimulating activity. J Immunol Methods 2004; 289:211 - 223
- Rouhiainen A, Tumova S, Valmu L, Kalkkinen N, Rauvala H. Pivotal advance: analysis of proinflammatory activity of highly purified eukaryotic recombinant HMGB1 (amphoterin). J Leukoc Biol 2007; 81:49 - 58
- Tian J, Avalos AM, Mao SY, Chen B, Senthil K, Wu H, et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated by HMGB1 and RAGE. Nat Immunol 2007; 8:487 - 496
- van Zoelen MA, Yang H, Florquin S, Meijers JC, Akira S, Arnold B, Nawroth PP, et al. Role of toll-like receptors 2 and 4, and the receptor for advanced glycation end products in high-mobility group box 1-induced inflammation in vivo. Shock 2009; 31:280 - 284
- Park JS, Gamboni-Robertson F, He Q, Svetkauskaite D, Kim JY, Strassheim D, Sohn JW, et al. High mobility group box 1 protein interacts with multiple Toll-like receptors. Am J Physiol Cell Physiol 2006; 290:C917 - C924
- Park JS, Svetkauskaite D, He Q, Kim JY, Strassheim D, Ishizaka A, et al. Involvement of toll-like receptors 2 and 4 in cellular activation by high mobility group box 1 protein. J Biol Chem 2004; 279:7370 - 7377
- Lee MC, Miller EA, Goldberg J, Orci L, Schekman R. Bi-directional protein transport between the ER and Golgi. Annu Rev Cell Dev Biol 2004; 20:87 - 123
- Trombetta ES, Parodi AJ. Quality control and protein folding in the secretory pathway. Annu Rev Cell Dev Biol 2003; 19:649 - 676
- Nickel W, Rabouille C. Mechanisms of regulated unconventional protein secretion. Nature Rev Mol Cell Biol 2009; 10:148 - 155
- Scaffidi P, Misteli T, Bianchi ME. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 2002; 418:191 - 195
- Rubartelli A, Cozzolino F, Talio M, Sitia R. A novel secretory pathway for interleukin-1 beta, a protein lacking a signal sequence. EMBO J 1990; 9:1503 - 1510
- Lamkanfi M, Dixit VM. Inflammasomes: guardians of cytosolic sanctity. Immunol Rev 2009; 227:95 - 105
- Kanneganti TD, Lamkanfi M, Nunez G. Intracellular NOD-like receptors in host defense and disease. Immunity 2007; 27:549 - 559
- O'Neill LA, Bowie AG. Sensing and signaling in antiviral innate immunity. Curr Biol 2010; 20:R328 - R333
- Lamkanfi M, Dixit VM. The inflammasomes. PLoS Pathog 2009; 5:e1000510
- Mariathasan S, Weiss DS, Newton K, McBride J, O'Rourke K, Roose-Girma M, et al. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 2006; 440:228 - 232
- Munoz-Planillo R, Franchi L, Miller LS, Nunez G. A critical role for hemolysins and bacterial lipoproteins in Staphylococcus aureus-induced activation of the Nlrp3 inflammasome. J Immunol 2009; 183:3942 - 3948
- Willingham SB, Allen IC, Bergstralh DT, Brickey WJ, Huang MT, Taxman DJ, et al. NLRP3 (NALP3, Cryopyrin) facilitates in vivo caspase-1 activation, necrosis, and HMGB1 release via inflammasome-dependent and -independent pathways. J Immunol 2009; 183:2008 - 2015
- Keller M, Ruegg A, Werner S, Beer HD. Active caspase-1 is a regulator of unconventional protein secretion. Cell 2008; 132:818 - 831
- Becker CE, Creagh EM, O'Neill LA. Rab39a binds caspase-1 and is required for caspase-1-dependent interleukin-1beta secretion. J Biol Chem 2009; 284:34531 - 34537
- Lamkanfi M, Dixit VM. Manipulation of host cell death pathways during microbial infections. Cell Host Microbe 2010; 8:44 - 54
- Lamkanfi M. Emerging inf lammasome effector mechanisms. Nat Rev Immunol 2011; 11:213 - 220
- Rovere-Querini P, Capobianco A, Scaffidi P, Valentinis B, Catalanotti F, Giazzon M, et al. HMGB1 is an endogenous immune adjuvant released by necrotic cells. EMBO Rep 2004; 5:825 - 830