
Abstract
Uropathogenic Escherichia coli (UPEC) produces ~80% of community-acquired UTI, the second most common infection in humans. During UTI, UPEC has a complex life cycle, replicating and persisting in intracellular and extracellular niches. Host and environmental stresses may affect the integrity of the UPEC genome and threaten its viability. We determined how the host inflammatory response during UTI drives UPEC genome instability and evaluated the role of multiple factors of genome replication and repair for their roles in the maintenance of genome integrity and thus virulence during UTI. The urinary tract environment enhanced the mutation frequency of UPEC ~100-fold relative to in vitro levels. Abrogation of inflammation through a host TLR4-signaling defect significantly reduced the mutation frequency, demonstrating in the importance of the host response as a driver of UPEC genome instability. Inflammation induces the bacterial SOS response, leading to the hypothesis that the UPEC SOS-inducible translesion synthesis (TLS) DNA polymerases would be key factors in UPEC genome instability during UTI. However, while the TLS DNA polymerases enhanced in vitro, they did not increase in vivo mutagenesis. Although it is not a source of enhanced mutagenesis in vivo, the TLS DNA polymerase IV was critical for the survival of UPEC during UTI during an active inflammatory assault. Overall, this study provides the first evidence of a TLS DNA polymerase being critical for UPEC survival during urinary tract infection and points to independent mechanisms for genome instability and the maintenance of genome replication of UPEC under host inflammatory stress.
Acknowledgments
The authors thank Drs. Joseph St. Geme, Ravi Jhaveri and Roel M. Schaaper for their thoughtful comments and critiques. Support for this work was partially provided by K08DK074443 and ORWH SCOR P50DK064540.
Figures and Tables
Figure 1 UTI promotes higher mutation frequencies in UTI89. (A) UTI89 infection rates (CFU/Bladder × 104 at 48 hpi). (B–D) mutation frequency per 108 cells measured for: 6-aza (B), lacI (C) and 5-FC (D) markers.
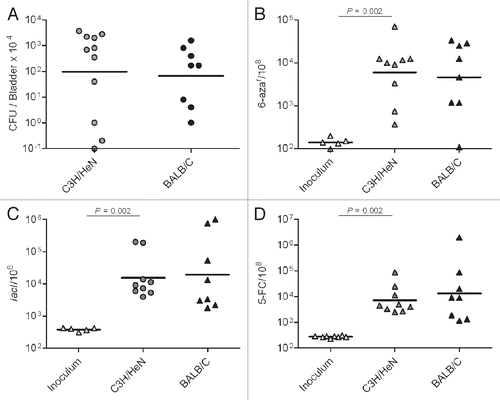
Figure 2 Growth under urine stress increases mutability for multiple E. coli strains. E. coli strains UPEC : CI5, E80, DS17, CFT073, GR12, PY2, UTI89, r-UTI89, NU14, EC45 and K-12: MG1655 and KA796 were grown overnight in LB broth and urine, in parallel, at 37°C. Mutation frequencies plotted as resistant colonies per 108 cells. Each plotted point is based on three independent cultures.
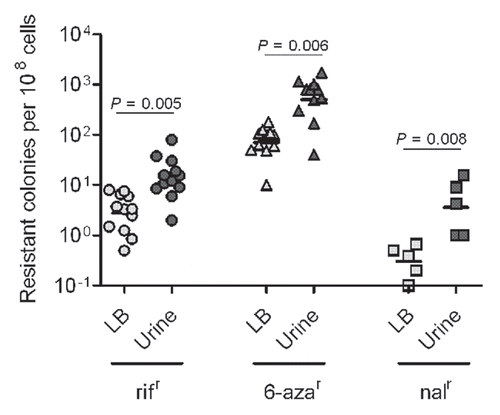
Figure 3 Host environment and innate immune response stimulate progressive increases mutation frequencies. Mutation frequencies are shown as number of 6-azar colonies per 108 cells.
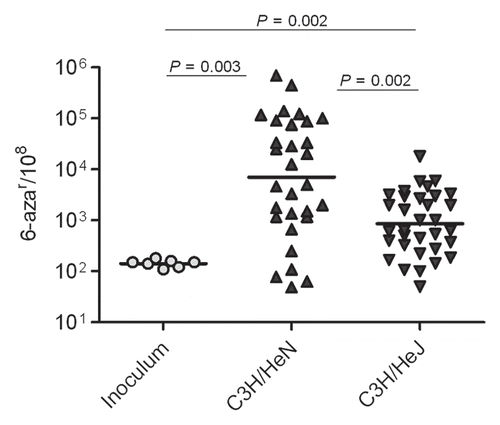
Figure 4 MMR is not suppressed or saturated during UTI. (A) Infection rates (CFU/Bladder × 104) of UTI89 and UTI89 ΔmutL strains. (B) Mutation frequencies per 108 cells measured for 6-azar marker.
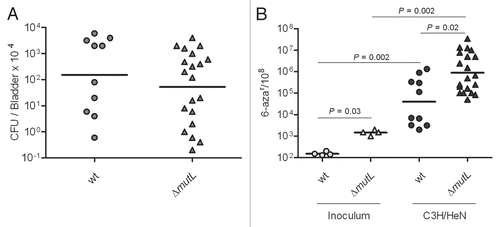
Figure 5 Mutation frequencies of UPEC UTI89 derivatives lacking genes encoding for TLS DNA polymerases. Strains were grown over night in LB broth and urine, in parallel, at 37°C. Mean mutation frequencies for 6-azar are based on eight independent cultures. Error bars show standard error (SE). Statistically significant differences (p < 0.05) in mutation rates frequencies compared to wt UTI89 are indicated by a star (*). p values were calculated using the non-parametric Mann-Whitney test.
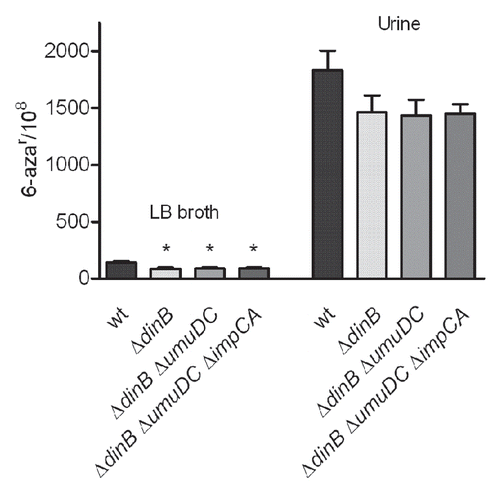
Figure 6 DNA polymerase IV improves survival of UTI89 strain in vivo but does not contribute to genome instability. (A) Infection rates (CFU/Bladder × 104) of UTI89 and its mutant derivatives. (B) Mutation frequencies per 108 cells measured for the 6-azar marker. The mutation frequencies were statistically different (p < 0.05) between the inocula and the bacteria recovered from infected bladders for all comparisons.
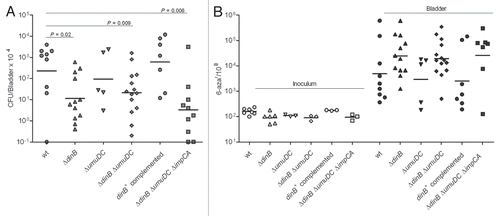
Figure 7 The ΔdinB mutant is not attenuated in the LPS hyporesponsive TLR4-host. (A) Infection rates (CFU/Bladder × 104) of UTI89 and its mutant derivatives. (B) mutation frequency per 108 cells measured for 6-azar marker. The mutation frequencies were statistically different (p < 0.05) between the inocula and the bacteria recovered from infected bladders for all comparisons.
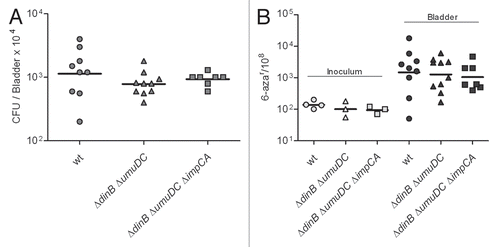
Table 1 In vitro mutation frequencies for UPEC UTI89Table Footnotea
Table 2 Growth in normal human urine increases the mutability of UPEC UTI89 in vitroTable Footnotea
Table 3 The UTI89 MMR mutant has enhanced mutability in vitroTable Footnotea
Table 4 E. coli strains used in this study