Abstract
The story of prevention and control of infectious diseases remains open and a series of highly virulent pathogens are emerging both in and beyond the hospital setting. Antibiotics were an absolute success story for a previous era. The academic and industrial biomedical communities have now come together to formulate consensus beliefs regarding the pursuit of novel and effective alternative anti-infective countermeasures. Photodynamic therapy was established and remains a successful modality for malignancies but photodynamic inactivation has been transformed recently to an antimicrobial discovery and development platform. The concept of photodynamic inactivation is quite straightforward and requires microbial exposure to visible light energy, typically wavelengths in the visible region, that causes the excitation of photosensitizer molecules (either exogenous or endogenous), which results in the production of singlet oxygen and other reactive oxygen species that react with intracellular components, and consequently produce cell inactivation. It is an area of increasing interest, as research is advancing i) to identify the photochemical and photophysical mechanisms involved in inactivation; ii) to develop potent and clinically compatible photosensitizer; iii) to understand how photoinactivation is affected by key microbial phenotypic elements (multidrug resistance and efflux, virulence and pathogenesis determinants, biofilms); iv) to explore novel delivery platforms inspired by current trends in pharmacology and nanotechnology; and v) to identify photoinactivation applications beyond the clinical setting such as environmental disinfectants.
Acknowledgments
Research conducted by Tyler G. St. Denis was supported by the Columbia University I. I. Rabi Fellows Program. George P. Tegos is supported by the NIH (grant 5U54MH084690-02). Research conducted in the Hamblin Laboratory was supported by NIH (RO1 AI050875 to M.R.H.) and US Air Force MFEL Program (FA9550-04-1-0079). T.D. was partially supported by a Bullock-Wellman Fellowship Award and an Airlift Research Foundation Extremity Trauma Research Grant (grant 109421).
Figures and Tables
Figure 1 Schematic illustration of photodynamic therapy including the Jablonski diagram. The PS initially absorbs a photon that excites it to the first excited singlet state and this can relax to the more long-lived triplet state. This triplet PS can interact with molecular oxygen in two pathways, type I and type II, leading to the formation of reactive oxygen species (ROS) and singlet oxygen respectively.
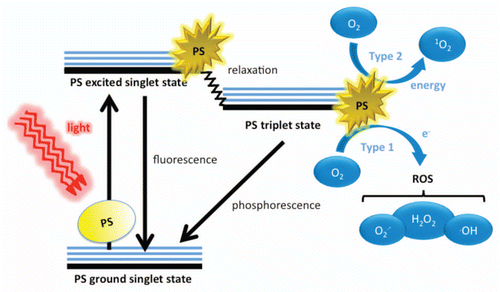
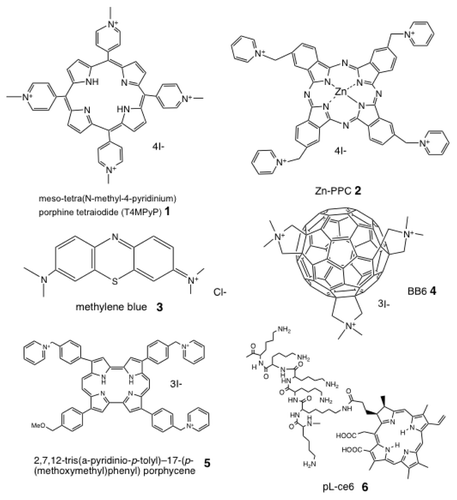
Figure 2 Chemical structures of some representative antimicrobial PS. (1) Cationic porphyrin, meso-tetra(N-methyl-4-pyridinium) porphine tetraiodide (T4MPyP); (2) cationic phthalocyanine, ZnPPC; (3) phenothiazinium salt, methylene blue; (4) cationic functionalized fullerene, BB6; (5) cationic porphycene, 2,7,12-tris(a-pyridinio-p-tolyl)-17-(p-(methoxymethyl)phenyl) porphycene; (6) poly-l-lysine chlorin(e6) conjugate, pL-ce6.